
Possible Contribution of Corticular Photosynthesis to Grapevine Winter Hardiness

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Weather
2.3. Variable Chlorophyll Fluorescence Measurements
2.3.1. Field Measurements
2.3.2. Laboratory Measurements
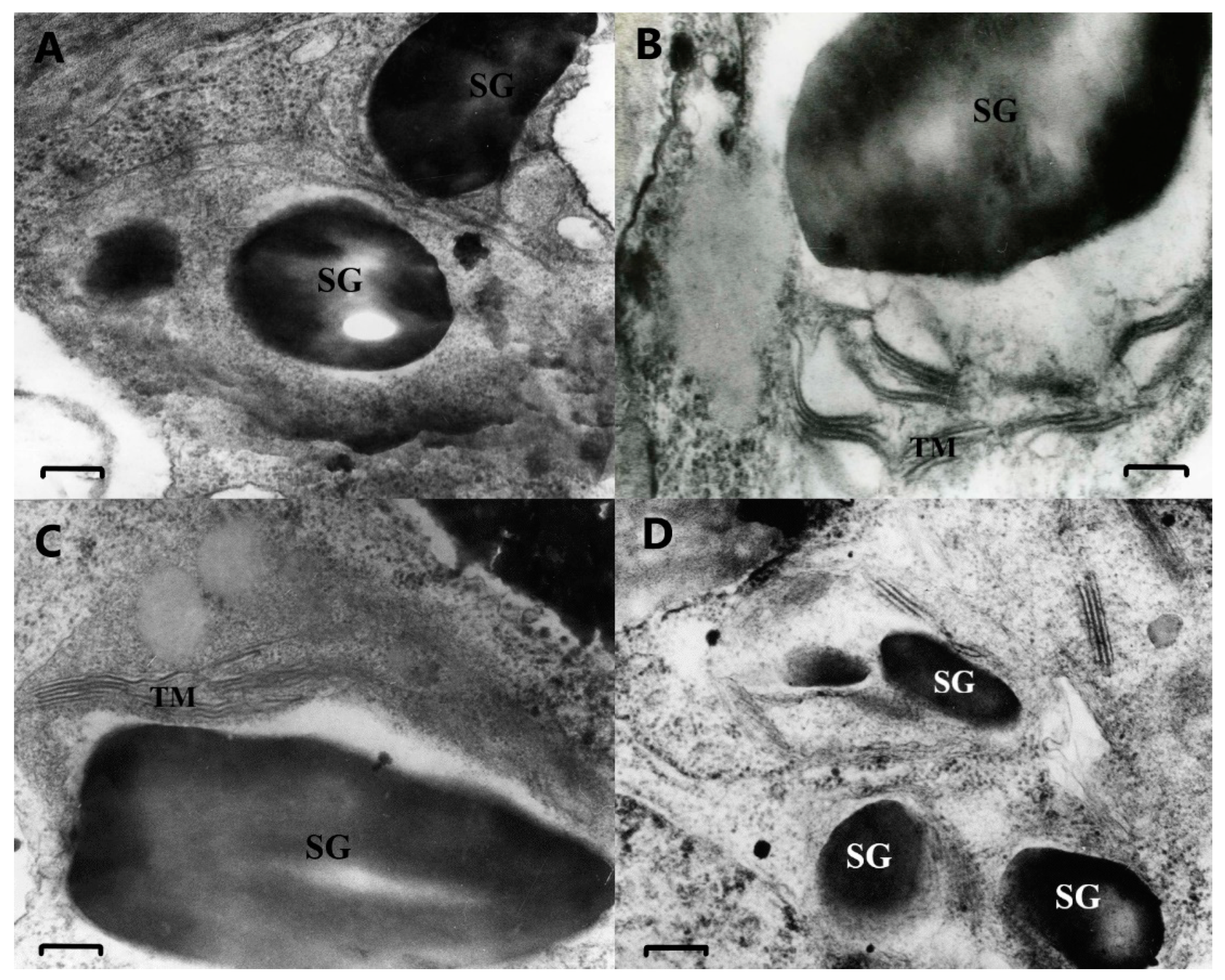
2.4. Microscopy Analysis
2.4.1. Electron Microscopy Analysis
2.4.2. Fluorescent Microscopy Analysis
2.5. Pigment Analysis
2.6. Statistical Analysis
3. Results
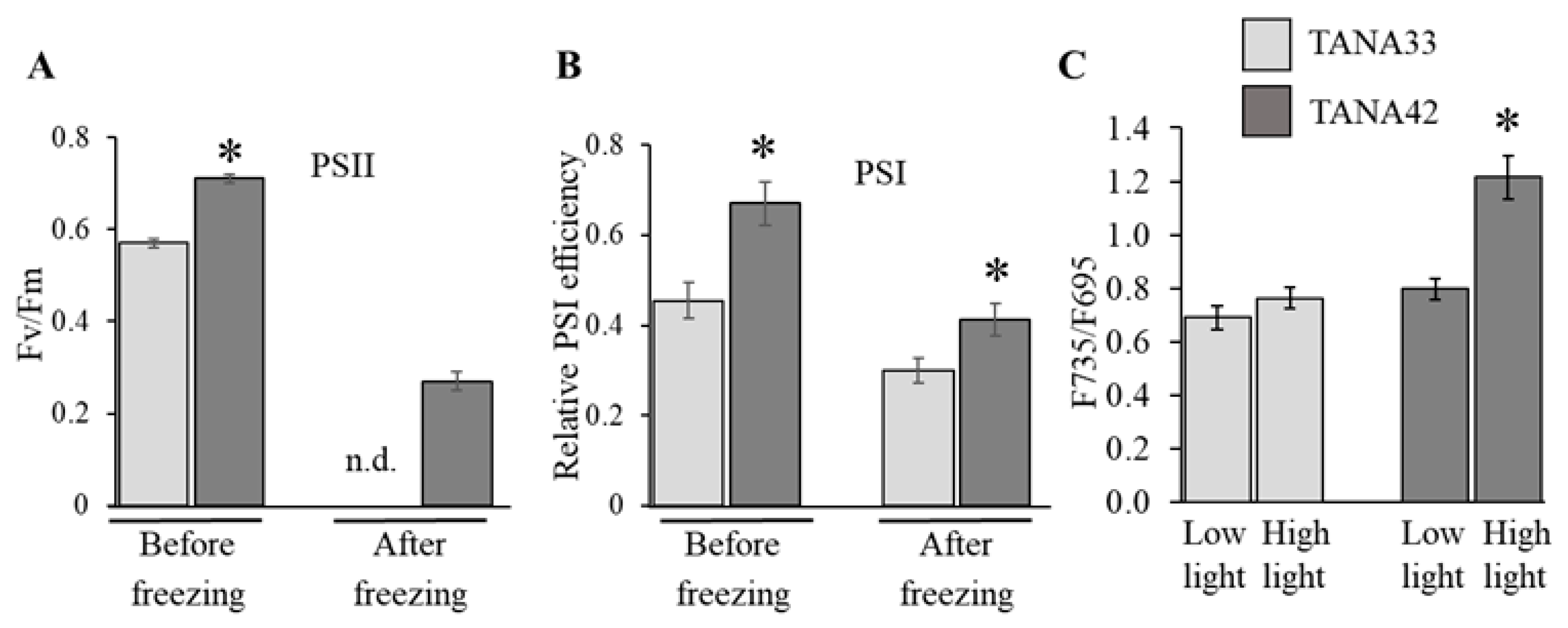
3.1. Analysis of the Y(II) of the Corticular Photosynthetic Apparatus of the Grapevine Varieties Differing in Frost Resistance
3.2. Comparative Analysis of Grapevine Hybrid Forms Differing in Freeze-Tolerance and CPA Activity
3.2.1. Laboratory Study
3.2.2. Field Study
3.3. Effect of Light on the Intensity of Freeze-Induced Injuries
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Available online: https://earthobservatory.nasa.gov/global-maps/MOD_LSTD_M (accessed on 4 June 2023).
- Bag, P.; Chukhutsina, V.; Zhang, Z.; Paul, S.; Ivanov, A.G.; Shutova, T.; Croce, R.; Holzwarth, A.R.; Jansson, S. Direct energy transfer from photosystem II to photosystem I confers winter sustainability in Scots Pine. Nat. Commun. 2020, 11, 6388. [Google Scholar] [CrossRef]
- Bag, P.; Shutova, T.; Shevela, D.; Lihavainen, J.; Nanda, S.; Ivanov, A.G.; Messinger, J.; Jansson, S. Flavodiiron-mediated O2 photoreduction at photosystem I acceptor-side provides photoprotection to conifer thylakoids in early spring. Nat. Commun. 2023, 14, 3210. [Google Scholar] [CrossRef] [PubMed]
- Oquist, G.; Huner, N.P. Photosynthesis of overwintering evergreen plants. Annu. Rev. Plant Biol. 2003, 54, 329–355. [Google Scholar] [CrossRef]
- Krol, M.; Hurry, V.M.; Maxwell, D.P.; Malek, L.; Ivanov, A.G.; Huner, N.P. Low growth temperature inhibition of photosynthesis in cotyledons of jack pine seedlings (Pinus banksiana) is due to impaired chloroplast development. Can. J. Bot. 2002, 80, 1042–1051. [Google Scholar] [CrossRef]
- Saveyn, A.; Steppe, K.; Ubierna, N.; Dawson, T.E. Woody tissue photosynthesis and its contribution to trunk growth and bud development in young plants. Plant Cell Environ. 2010, 33, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, K.G.; Khristin, M.S.; Klimov, V.V.; Sundireva, M.A.; Kreslavski, V.D.; Sidorov, R.A.; Tsidendambayev, V.D.; Savchenko, T.V. Structural and functional characteristics of photosynthetic apparatus of chlorophyll-containing grape vine tissue. Russ. J. Plant Phys. 2017, 64, 73–82. [Google Scholar] [CrossRef]
- Yanykin, D.; Sundyreva, M.; Khorobrykh, A.; Semenova, G.; Savchenko, T. Functional characterization of the corticular photosynthetic apparatus in grapevine. Biochim. Biophys. Acta (BBA) Bioenerg. 2020, 1861, 148260. [Google Scholar] [CrossRef]
- Aschan, G.; Pfanz, H. Non-foliar photosynthesis—A strategy of additional carbon acquisition. Flora Morphol. Distrib. Funct. Ecol. Plants 2003, 198, 81–97. [Google Scholar] [CrossRef]
- Wittmann, C.; Pfanz, H.; Loreto, F.; Centritto, M.; Pietrini, F.; Alessio, G. Stem CO2 release under illumination: Corticular photosynthesis, photorespiration or inhibition of mitochondrial respiration? Plant Cell Environ. 2006, 29, 1149–1158. [Google Scholar] [CrossRef]
- Ávila-Lovera, E.; Garcillán, P.P.; Silva-Bejarano, C.; Santiago, L.S. Functional traits of leaves and photosynthetic stems of species from a sarcocaulescent scrub in the southern Baja California Peninsula. Am. J. Bot. 2020, 107, 1410–1422. [Google Scholar] [CrossRef]
- Burrows, G.E.; Connor, C. Chloroplast Distribution in the Stems of 23 Eucalypt Species. Plants 2020, 9, 1814. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Luo, M.; Kang, Y.; Zhao, P.; Tang, Z.; Meng, Y.; Huang, L.; Guo, Y.; Lu, X.; Ouyang, L.; et al. Comparison between the stem and leaf photosynthetic productivity in Eucalyptus urophylla plantations with different age. Planta 2023, 257, 56. [Google Scholar] [CrossRef] [PubMed]
- Natale, S.; La Rocca, N.; Battistuzzi, M.; Morosinotto, T.; Nardini, A.; Alboresi, A. Structure and function of bark and wood chloroplasts in a drought-tolerant tree (Fraxinus ornus L.). Tree Physiol. 2023, 43, 893–908. [Google Scholar] [CrossRef]
- Savchenko, T.V.; Sundyreva, M.A.; Khristin, M.S.; Klimov, V.V.; Biryukov, S.V. Method of the grapes frost resistance determining. 2018. [Google Scholar]
- Ketel, D.H.; Wielen, M.J.W.V.D.; Lotz, L.A.P. Prediction of a low dose herbicide effect from studies on binding of metribuzin to the chloroplasts of Chenopodium album L. Ann. Appl. Biol. 1996, 128, 519–531. [Google Scholar] [CrossRef]
- Genty, B.; Harbinson, J.; Cailly, A.; Rizza, F. Fate of excitation at PS II in leaves: The non-photochemical side. Plant Physiol. Biochem. 1996, 86, 28. [Google Scholar]
- Lichtenthaller, H.K.; Wellburn, A.R. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzym. 1987, 148, 350–382. [Google Scholar]
- Kufner, R.; Czygan, F.C.; Schneider, L. Changes in pigment content and ultrastructure in the plastids of the needle leaves of Taxus baccata (L.) during their development. Ber. Deut. Bot. Ges. 1978, 91, 325–337. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence as a tool in plant physiology: II. Interpretation of fluorescence signals. Photosynth. Res. 1984, 5, 139–157. [Google Scholar] [CrossRef]
- Minagawa, J. State transitions—The molecular remodeling of photosynthetic supercomplexes that controls energy flow in the chloroplast. Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1807, 897–905. [Google Scholar] [CrossRef]
- Allen, J.F.; Bennett, J.; Steinback, K.E.; Arntzen, C.J. Chloroplast protein phosphorylation couples plastoquinone redox state to distribution of excitation energy between photosystems. Nature 1981, 291, 25–29. [Google Scholar] [CrossRef]
- Bennett, J. Phosphorylation of chloroplast membrane polypeptides. Nature 1977, 269, 344–346. [Google Scholar] [CrossRef]
- Murata, N. Control of excitation transfer in photosynthesis I. Light-induced change of chlorophyll a fluoresence in Porphyridium cruentum. Biochim. Biophys. Acta (BBA) Bioenerg. 1969, 172, 242–251. [Google Scholar] [CrossRef]
- Egerton, J.J.G.; Banks, J.C.G.; Gibson, A.; Cunningham, R.B.; Ball, M.C. Facilitation of seedling establishment: Reduction in irradiance enhances winter growth of Eucalyptus pauciflora. Ecology 2000, 81, 1437–1449. [Google Scholar] [CrossRef]
- Öquist, G.; Ögren, E. Effects of winter stress on photosynthetic electron transport and energy distribution between the two photosystems of pine as assayed by chlorophyll fluorescence kinetics. Photosynth. Res. 1985, 7, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.M.; Warren, J.M.; Hanson, P.J.; Childs, J.; Wullschleger, S.D. Needle age and season influence photosynthetic temperature response and total annual carbon uptake in mature Picea mariana trees. Ann. Bot. 2015, 116, 821–832. [Google Scholar] [CrossRef]
- Yang, Q.; Blanco, N.E.; Hermida-Carrera, C.; Lehotai, N.; Hurry, V.; Strand, Å. Two dominant boreal conifers use contrasting mechanisms to reactivate photosynthesis in the spring. Nat. Commun. 2020, 11, 128. [Google Scholar] [CrossRef]
- Gunderson, C.A.; O’hara, K.H.; Campion, C.M.; Walker, A.V.; Edwards, N.T. Thermal plasticity of photosynthesis: The role of acclimation in forest responses to a warming climate. Glob. Change Biol. 2010, 16, 2272–2286. [Google Scholar] [CrossRef]
- Kenchanmane Raju, S.K.; Barnes, A.C.; Schnable, J.C.; Roston, R.L. Low-temperature tolerance in land plants: Are transcript and membrane responses conserved? Plant Sci. 2018, 276, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.G.; Krol, M.; Sveshnikov, D.; Malmberg, G.; Gardestrom, P.; Hurry, V.; Oquist, G.; Huner, N.P. Characterization of the photosynthetic apparatus in cortical bark chlorenchyma of Scots pine. Planta 2006, 223, 1165–1177. [Google Scholar] [CrossRef]
- Damesin, C. Respiration and photosynthesis characteristics of current-year stems of Fagus sylvatica: From the seasonal pattern to an annual balance. New Phytol. 2003, 158, 465–475. [Google Scholar] [CrossRef]
- Tanja, S.; Berninger, F.; Vesala, T.; Markkanen, T.; Hari, P.; Mäkelä, A.; Ilvesniemi, H.; Hanninen, H.; Nikinmaa, E.; Huttula, T.; et al. Air Temperature Triggers the Recovery of Evergreen Boreal Forest Photosynthesis in Spring. Glob. Change Biol. 2003, 9, 1410–1426. [Google Scholar] [CrossRef]
- Monson, R.K.; Sparks, J.P.; Rosenstiel, T.N.; Scott-Denton, L.E.; Huxman, T.E.; Harley, P.C.; Turnipseed, A.A.; Burns, S.P.; Backlund, B.; Hu, J. Climatic influences on net ecosystem CO2 exchange during the transition from wintertime carbon source to springtime carbon sink in a high-elevation, subalpine forest. Oecologia 2005, 146, 130–147. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, D.; Tausz, M.; Moore, G.; Nicolas, M. Bark and leaf chlorophyll fluorescence are linked to wood structural changes in Eucalyptus saligna. AoB Plants 2014, 6, plt057. [Google Scholar] [CrossRef] [PubMed]
- Fennell, A. Freezing tolerance and injury in grapevines. J. Crop Improv. 2004, 10, 201–235. [Google Scholar] [CrossRef]
- Ren, C.; Fan, P.; Li, S.; Liang, Z. Advances in understanding cold tolerance in grapevine. Plant Physiol. 2023, 192, 1733–1746. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, Y.; Xia, H.; Fan, S.; Song, J.; Lv, X.; Kong, L. Photosynthetic characteristics of non-foliar organs in main C(3) cereals. Physiol. Plant 2019, 166, 226–239. [Google Scholar] [CrossRef]
- Pfanz, H.; Aschan, G.; Langenfeld-Heyser, R.; Wittmann, C.; Loose, M. Ecology and ecophysiology of tree stems: Corticular and wood photosynthesis. Die Nat. 2002, 89, 147–162. [Google Scholar]
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2020, 101, 1001–1015. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigment | TANA33 | TANA42 |
|---|---|---|
| Chlorophyll a (μg/g fresh weight) | 52.2 ± 2.8 | 70.8 ± 1.2 * |
| Chlorophyll b (μg/g fresh weight) | 24.8 ± 4 | 35.4 ± 3.8 * |
| Chlorophyll a + b (μg/g fresh weight) | 77 ± 6.4 | 106 ± 5.0 * |
| Carotenoids (μg/g fresh weight) | 34.2 ± 1.6 | 32.6 ± 1.4 |
| Chl a/Chl b | 2.25 ± 0.26 | 2.09 ± 0.21 |
| Chl/carotenoids | 2.29 ± 0.29.8 | 3.31 ± 0.28 * |
| Parameter | TANA33 | TANA42 |
|---|---|---|
| Sum of daytime temperatures, °C | 0.610 * | 0.790 * |
| Temperature during measurement, °C | 0.617 * | 0.805 * |
| Average temperature during 12 h before the measurement, °C | 0.590 * | 0.752 * |
| Average temperature during 24 h before measurement, °C | 0.561 * | 0.738 * |
| Average temperature during 48 h before measurement, °C | 0.455 | 0.669 * |
| Minimum temperature on the day of measurement, °C | 0.599 * | 0.750 * |
| Minimum temperature during 24 h before measurement, °C | 0.599 * | 0.742 * |
| Minimum temperature during 48 h before measurement, °C | 0.585 * | 0.633 * |
| Precipitation, mm | −0.438 | −0.514 * |
| Solar radiation, W/m2 | 0.119 | 0.119 |
| Relative air humidity, % | −0.014 | −0.240 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sundyreva, M.A.; Yanykin, D.V.; Khristin, M.S.; Gryaznova, U.V.; Lutskiy, E.O.; Semenova, G.A.; Naydov, I.A.; Savchenko, T.V. Possible Contribution of Corticular Photosynthesis to Grapevine Winter Hardiness. Horticulturae 2023, 9, 1181. https://doi.org/10.3390/horticulturae9111181
Sundyreva MA, Yanykin DV, Khristin MS, Gryaznova UV, Lutskiy EO, Semenova GA, Naydov IA, Savchenko TV. Possible Contribution of Corticular Photosynthesis to Grapevine Winter Hardiness. Horticulturae. 2023; 9(11):1181. https://doi.org/10.3390/horticulturae9111181
Chicago/Turabian StyleSundyreva, Maria A., Denis V. Yanykin, Mikhail S. Khristin, Ulyana V. Gryaznova, Evgenij O. Lutskiy, Galina A. Semenova, Ilya A. Naydov, and Tatyana V. Savchenko. 2023. "Possible Contribution of Corticular Photosynthesis to Grapevine Winter Hardiness" Horticulturae 9, no. 11: 1181. https://doi.org/10.3390/horticulturae9111181
APA StyleSundyreva, M. A., Yanykin, D. V., Khristin, M. S., Gryaznova, U. V., Lutskiy, E. O., Semenova, G. A., Naydov, I. A., & Savchenko, T. V. (2023). Possible Contribution of Corticular Photosynthesis to Grapevine Winter Hardiness. Horticulturae, 9(11), 1181. https://doi.org/10.3390/horticulturae9111181