
From Lab to Field: Biofertilizers in the 21st Century


Abstract
:1. Introduction
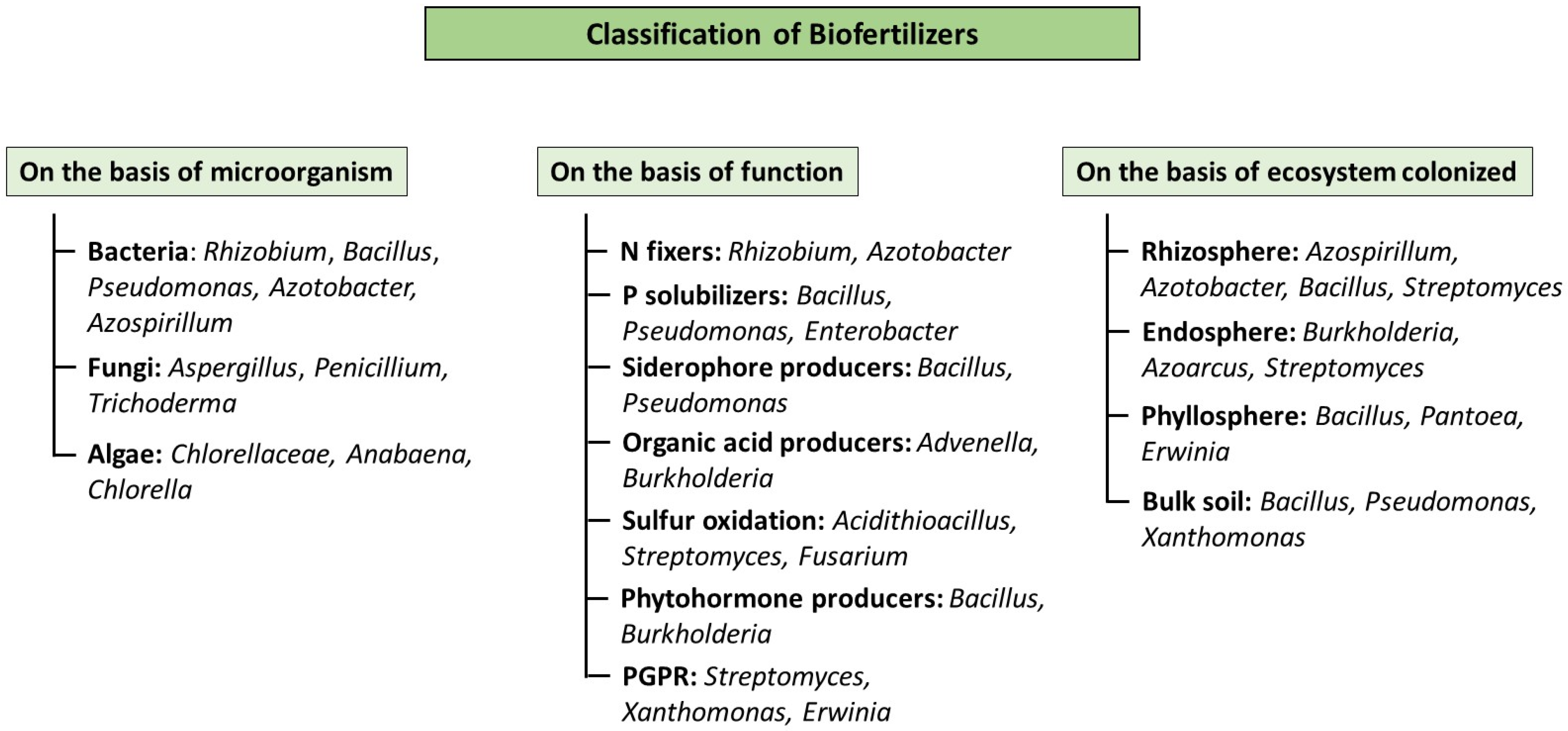
2. Unearthing the Basics
3. The Microscopic Maestros: Common Organisms in Biofertilizers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofertilizer Microorganisms | Action Mechanisms | Host Plant | Ref. |
|---|---|---|---|
| Advenella mimigardefordensis | Plant growth promotion; organic acid production; P and K solubilization; antifungal activity | Barley | [25] |
| Astragalus mongholicus | [35] | ||
| Aspergillus flavus | Increase antioxidant enzyme activity and chlorophyll content | Glycine max | [36] |
| Bacillus amyloliquefaciens | Induce SA and JA signalling, enhancing plant protection against pathogens | Solanum lycopersicum | [37] |
| Plant growth promotion | Arabidopsis thaliana | [38] | |
| Bacillus aryabhattai | Improve tolerance to salt stress | Oryzae sativa | [39] |
| Bacillus cereus | Plant growth promotion, organic acid production, P solubilization, phytohormone production | Arabidopsis thaliana | [40] |
| Barley | [25] | ||
| Potatoe | [41] | ||
| Bacillus licheniformis | Improve tolerance to salt stress | Chrysanthemum | [42] |
| Bacillus megaterium | Plant growth promotion; organic acid production; P solubilization; antifungal activity | Cucumis melo | [43] |
| Barley | [25] | ||
| Bacillus subtilis | Plant growth promotion; antimicrobial activity | Atractylodes macrocephala | [44] |
| Increase expression of auxin-related genes; plant growth promotion | Solanum lycopersicum | [45] | |
| Plant growth promotion; improve tolerance to infections | Oryzae sativa | [46] | |
| Bacillus velezensis | Protect plant from pathogens via systemic resistance response | Arabidopsis thaliana | [47] |
| Burkholderia fungorum | Plant growth promotion; organic acid production; P and K solubilization; antifungal activity | Barley | [25] |
| Burkholderia contaminans | Antifungal activity | Maize | [48] |
| Enterobacter cloacae | Plant growth promotion; organic acid and phytohormones production; P, Zn, and K solubilization; antifungal activity; N fixation | Barley | [25] |
| Abelmoschus esculentus | [49] | ||
| - | [50] | ||
| Paenibacillus polymyxa | Increase production of volatile fatty acids and antibiotics | Brassica napus | [51] |
| Pseudomonas aeruginosa | Protect plant from pathogens via chitinase production | Cruciferous vegetables | [52] |
| Pseudomonas koreensis | Plant growth promotion; organic acid production; P, Zn, and K solubilization; antifungal activity | Barley | [25] |
| Arabidopsis thaliana | [53] | ||
| Pseudomonas plecoglossicida | Plant growth promotion, organic acid production; P, Zn, and K solubilization; antifungal activity | - | [54,55] |
| Barley | [25] | ||
| Plant growth promotion | Triticum aestivum | [56] | |
| Antimicrobial activity; plant growth promotion | Capsicum annuum | [57] | |
| Pseudomonas putida | Increase the production of HCN against pathogens | Solanum tuberosum | [58] |
| Antimicrobial activity | Oryzae sativa | [59] | |
| Enhance urease, phosphatase, and invertase activity | Carthamus tinctorius | [60] | |
| Streptomyces spp. | Enhance plant immunity via increasing antioxidant enzymes | Oryzae sativa | [61] |
| Plant growth promotion; infection tolerance improvement; siderophore and VOC production; phosphate solubilization | Solanum lycopersicum | [62] | |
| Nitrogen fixation; phytohormone production | Solanum tuberosum | [63] | |
| Phytohormone production | Cereal crops | [64] | |
| Trichoderma atroviridae | Improve auxin production; tolerance to cold stress | Arabidopsis thaliana | [65] |
| Trichoderma harzianum | Provide protection from pathogens via JA signalling | Zea mays | [66] |
| Antimicrobial activity | Bupleurum chinense | [67] | |
| Increase colonization of non-host mycorrhizal plants | Arabidopsis thaliana, Brassica napus | [68] | |
| Trichoderma koningii | Antimicrobial activity | Nicotiana tabacum | [69] |
| Plant growth promotion; improve tolerance to abiotic stresses | Solanum lycopersicum | [70] | |
| Antimicrobial activity against Rhizoctonia solani | Cotton | [71] |
4. Biofertilizer Frontiers: Exploring New Sources
4.1. Sailing the Microbial Seas: Marine Microorganisms and Microalgae
4.2. Extreme Allies: Extremophilic Microorganisms against Abiotic Stress
4.3. Unisolated Microorganisms: How to Discover These Unseen Treasures
4.4. Customized Microbial Inoculants
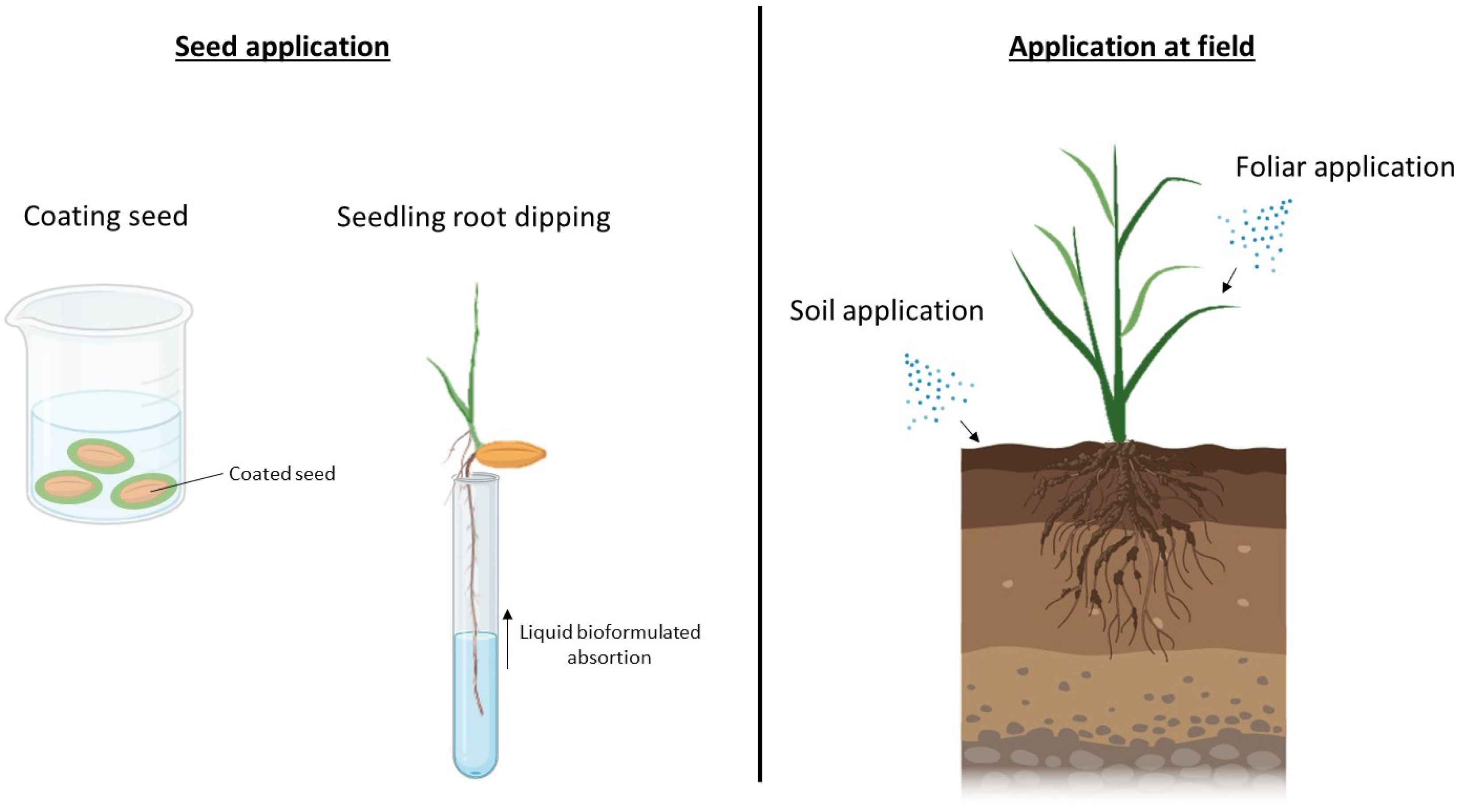
5. New Possibilities in Biofertilizer Application Techniques
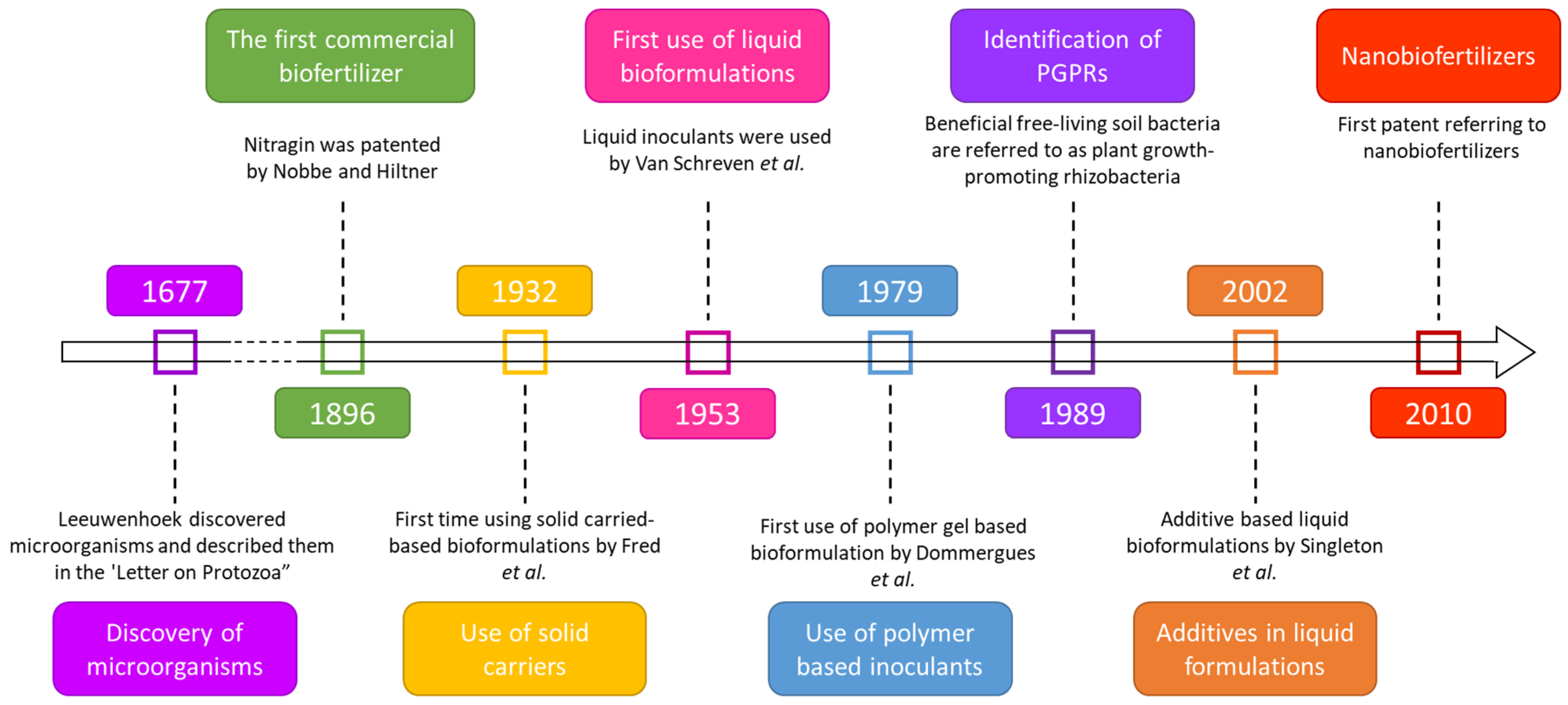
5.1. Biofertilizers: Inception

5.2. Microencapsulation and Nanobiofertilizers
5.3. Sprays and Foliar Application
5.4. Mulch Application
6. Overcoming Challenges in Biofertilizer Use
6.1. Scientific and Technological Concerns
6.2. Environmental Aspects
6.3. Practical Challenges
7. Biofertilizer Regulation
8. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Agricultural Production Statistics 2000–2021; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- Chaudhary, P.; Singh, S.; Chaudhary, A.; Sharma, A.; Kumar, G. Overview of Biofertilizers in Crop Production and Stress Management for Sustainable Agriculture. Front. Plant Sci. 2022, 13, 930340. [Google Scholar] [CrossRef]
- Allouzi, M.M.A.; Allouzi, S.M.A.; Keng, Z.X.; Supramaniam, C.V.; Singh, A.; Chong, S. Liquid Biofertilizers as a Sustainable Solution for Agriculture. Heliyon 2022, 8, e12609. [Google Scholar] [CrossRef] [PubMed]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as Biofertilizers, a Potential Approach for Sustainable Crop Production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Ibáñez, A.; Garrido-Chamorro, S.; Barreiro, C. Microorganisms and Climate Change: A Not so Invisible Effect. Microbiol. Res. 2023, 14, 918–947. [Google Scholar] [CrossRef]
- Adeyinka Fasusi, O.; Cruz, C.; Oluranti Babalola, O.; Agricultural, O.; Hasanuzzaman, M.; Ahmad Khan, E. Agricultural Sustainability: Microbial Biofertilizers in Rhizosphere Management. Agriculture 2021, 11, 163. [Google Scholar] [CrossRef]
- Mahmud, A.A.; Upadhyay, S.K.; Srivastava, A.K.; Bhojiya, A.A. Biofertilizers: A Nexus between Soil Fertility and Crop Productivity under Abiotic Stress. Curr. Res. Environ. Sustain. 2021, 3, 100063. [Google Scholar] [CrossRef]
- Mehra, A. Biofertilizers Market Worth $5.2 Billion by 2028; MarketsandMarkets: Pune, India, 2020. [Google Scholar]
- Pirttilä, A.M.; Tabas, H.M.P.; Baruah, N.; Koskimäki, J.J. Biofertilizers and Biocontrol Agents for Agriculture: How to Identify and Develop New Potent Microbial Strains and Traits. Microorganisms 2021, 9, 817. [Google Scholar] [CrossRef] [PubMed]
- Visser, J. Opening History: Gaining Perspectives. In Organic Fertilizers: History, Production and Applications; Larramendy, M., Soloneski, S., Eds.; IntechOpen: London, UK, 2019; pp. 1–22. [Google Scholar]
- Chojnacka, K.; Moustakas, K.; Witek-Krowiak, A. Bio-Based Fertilizers: A Practical Approach towards Circular Economy. Bioresour. Technol. 2020, 295, 122223. [Google Scholar] [CrossRef]
- Ortiz, A.; Sansinenea, E. The Role of Beneficial Microorganisms in Soil Quality and Plant Health. Sustainability 2022, 14, 5358. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Wood, E.E.; Llewellyn, C.A. Algae Biostimulants: A Critical Look at Microalgal Biostimulants for Sustainable Agricultural Practices. Biotechnol. Adv. 2021, 49, 107754. [Google Scholar] [CrossRef]
- Raimi, A.; Roopnarain, A.; Adeleke, R. Biofertilizer Production in Africa: Current Status, Factors Impeding Adoption and Strategies for Success. Sci. Afr. 2021, 11, e00694. [Google Scholar] [CrossRef]
- Htwe, A.Z.; Moh, S.M.; Moe, K.; Yamakawa, T. Biofertilizer Production for Agronomic Application and Evaluation of Its Symbiotic Effectiveness in Soybeans. Agronomy 2019, 9, 162. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; Enshasy, H. El Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef] [PubMed]
- Clúa, J.; Roda, C.; Zanetti, M.E.; Blanco, F.A. Compatibility between Legumes and Rhizobia for the Establishment of a Successful Nitrogen-Fixing Symbiosis. Genes 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Day, D.A.; Smith, P.M.C. Iron Transport across Symbiotic Membranes of Nitrogen-Fixing Legumes. Int. J. Mol. Sci. 2021, 22, 432. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Nobbe, F.; Hiltner, L. Inoculation of the Soil for Cultivating Leguminous Plant. U.S. Patent 570 813, 3 November 1896. [Google Scholar]
- Mitter, E.K.; Tosi, M.; Obregón, D.; Dunfield, K.E.; Germida, J.J. Rethinking Crop Nutrition in Times of Modern Microbiology: Innovative Biofertilizer Technologies. Front. Sustain. Food Syst. 2021, 5, 606815. [Google Scholar] [CrossRef]
- Dey, A. Liquid Biofertilizers and Their Applications: An Overview. In Environmental and Agricultural Microbiology; Wiley: Hoboken, NJ, USA, 2021; pp. 275–292. [Google Scholar]
- Patel, C.; Singh, J.; Karunakaran, A.; Ramakrishna, W. Evolution of Nano-Biofertilizer as a Green Technology for Agriculture. Agriculture 2023, 13, 1865. [Google Scholar] [CrossRef]
- Ibáñez, A.; Diez-Galán, A.; Cobos, R.; Calvo-Peña, C.; Barreiro, C.; Medina-Turienzo, J.; Sánchez-García, M.; Coque, J.J.R. Using Rhizosphere Phosphate Solubilizing Bacteria to Improve Barley (Hordeum vulgare) Plant Productivity. Microorganisms 2021, 9, 1619. [Google Scholar] [CrossRef]
- Munir, N.; Hanif, M.; Abideen, Z.; Sohail, M.; El-Keblawy, A.; Radicetti, E.; Mancinelli, R.; Haider, G. Mechanisms and Strategies of Plant Microbiome Interactions to Mitigate Abiotic Stresses. Agronomy 2022, 12, 2069. [Google Scholar] [CrossRef]
- Rahman, N.S.N.A.; Hamid, N.W.A.; Nadarajah, K. Effects of Abiotic Stress on Soil Microbiome. Int. J. Mol. Sci. 2021, 22, 9036. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Cao, Q.; Sun, L.; Yang, X.; Yang, W.; Zhang, H. Stomatal Conductance and Morphology of Arbuscular Mycorrhizal Wheat Plants Response to Elevated CO2 and NaCl Stress. Front. Plant Sci. 2018, 9, 410525. [Google Scholar] [CrossRef] [PubMed]
- Metwally, R.A.; Abdelhameed, R.E. Synergistic Effect of Arbuscular Mycorrhizal Fungi on Growth and Physiology of Salt-Stressed Trigonella Foenum-Graecum Plants. Biocatal. Agric. Biotechnol. 2018, 16, 538–544. [Google Scholar] [CrossRef]
- Khaliq, A.; Perveen, S.; Alamer, K.H.; Ul Haq, M.Z.; Rafique, Z.; Alsudays, I.M.; Althobaiti, A.T.; Saleh, M.A.; Hussain, S.; Attia, H. Arbuscular Mycorrhizal Fungi Symbiosis to Enhance Plant–Soil Interaction. Sustainability 2022, 14, 7840. [Google Scholar] [CrossRef]
- Gou, X.; Ni, H.; Sadowsky, M.J.; Chang, X.; Liu, W.; Wei, X. Arbuscular Mycorrhizal Fungi Alleviate Erosion-Induced Soil Nutrient Losses in Experimental Agro-Ecosystems. Catena 2023, 220, 106687. [Google Scholar] [CrossRef]
- El-Sawah, A.; El-Keblawy, A.; Ali, D.; Ibrahim, H.; El-Sheikh, M.; Sharma, A.; Alhaj Hamoud, Y.; Shaghaleh, H.; Brestic, M.; Skalicky, M.; et al. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Enhance Soil Key Enzymes, Plant Growth, Seed Yield, and Qualitative Attributes of Guar. Agriculture 2021, 11, 194. [Google Scholar] [CrossRef]
- Odoh, C.K.; Sam, K.; Zabbey, N.; Eze, C.N.; Nwankwegu, A.S.; Laku, C.; Dumpe, B.B. Microbial Consortium as Biofertilizers for Crops Growing Under the Extreme Habitats. In Plant Microbiomes for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 381–424. [Google Scholar]
- Brenner, K.; You, L.; Arnold, F.H. Engineering Microbial Consortia: A New Frontier in Synthetic Biology. Trends Biotechnol. 2008, 26, 483–489. [Google Scholar] [CrossRef]
- Li, Z.; Bai, X.; Jiao, S.; Li, Y.; Li, P.; Yang, Y.; Zhang, H.; Wei, G. A Simplified Synthetic Community Rescues Astragalus mongholicus from Root Rot Disease by Activating Plant-Induced Systemic Resistance. Microbiome 2021, 9, 217. [Google Scholar] [CrossRef]
- Lubna; Asaf, S.; Hamayun, M.; Khan, A.L.; Waqas, M.; Khan, M.A.; Jan, R.; Lee, I.-J.; Hussain, A. Salt Tolerance of Glycine max L. Induced by Endophytic Fungus Aspergillus flavus CSH1, via Regulating Its Endogenous Hormones and Antioxidative System. Plant Physiol. Biochem. 2018, 128, 13–23. [Google Scholar] [CrossRef]
- Beris, D.; Theologidis, I.; Skandalis, N.; Vassilakos, N. Bacillus amyloliquefaciens Strain MBI600 Induces Salicylic Acid Dependent Resistance in Tomato Plants against Tomato Spotted Wilt Virus and Potato Virus Y. Sci. Rep. 2018, 8, 10320. [Google Scholar] [CrossRef]
- Liu, S.; Tian, Y.; Jia, M.; Lu, X.; Yue, L.; Zhao, X.; Jin, W.; Wang, Y.; Zhang, Y.; Xie, Z.; et al. Induction of Salt Tolerance in Arabidopsis thaliana by Volatiles From Bacillus amyloliquefaciens FZB42 via the Jasmonic Acid Signaling Pathway. Front. Microbiol. 2020, 11, 562934. [Google Scholar] [CrossRef]
- Sultana, S.; Paul, S.C.; Parveen, S.; Alam, S.; Rahman, N.; Jannat, B.; Hoque, S.; Rahman, M.T.; Karim, M.M. Isolation and Identification of Salt-Tolerant Plant-Growth-Promoting Rhizobacteria and Their Application for Rice Cultivation under Salt Stress. Can. J. Microbiol. 2020, 66, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.; Chen, C.; Yin, Q.; Jiang, C.; Guo, J.; Zhao, H.; Niu, D. Function of MiR825 and MiR825* as Negative Regulators in Bacillus cereus AR156-Elicited Systemic Resistance to Botrytis cinerea in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 5032. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.M.; Awad, M.Y.M.; Hegab, S.A.; El Gawad, A.M.A.; Eissa, M.A. Effect of Potassium Solubilizing Bacteria (Bacillus cereus) on Growth and Yield of Potato. J. Plant Nutr. 2021, 44, 411–420. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, L.; Xie, Y.; Li, F.; Xiao, X.; Ma, Z.; Wang, J. Bacillus licheniformis SA03 Confers Increased Saline–Alkaline Tolerance in Chrysanthemum Plants by Induction of Abscisic Acid Accumulation. Front. Plant Sci. 2017, 8, 1143. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-M.; Radhakrishnan, R.; Lee, K.-E.; You, Y.-H.; Ko, J.-H.; Kim, J.-H.; Lee, I.-J. Mechanism of Plant Growth Promotion Elicited by Bacillus Sp. LKE15 in Oriental Melon. Acta Agric. Scand. B Soil. Plant Sci. 2015, 65, 637–647. [Google Scholar] [CrossRef]
- You, J.-M.; Xiong, K.; Mu, S.; Guo, J.; Guo, X.-L.; Duan, Y.-Y.; Li, J.; Cao, F.; Zou, Z.-C.; Tan, H. Identification of Endophytic Bacteria BZJN1 and Research on Biological Control of Root Rot of Atractylodes macrocephala. Zhongguo Zhong Yao Za Zhi 2018, 3, 478–483. [Google Scholar]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus subtilis MBI600 Promotes Growth of Tomato Plants and Induces Systemic Resistance Contributing to the Control of Soilborne Pathogens. Plants 2021, 10, 1113. [Google Scholar] [CrossRef]
- Ji, S.-H.; Kim, J.-S.; Lee, C.-H.; Seo, H.-S.; Chun, S.-C.; Oh, J.; Choi, E.-H.; Park, G. Enhancement of Vitality and Activity of a Plant Growth-Promoting Bacteria (PGPB) by Atmospheric Pressure Non-Thermal Plasma. Sci. Rep. 2019, 9, 1044. [Google Scholar] [CrossRef]
- Rashid, M.H.-O.; Khan, A.; Hossain, M.T.; Chung, Y.R. Induction of Systemic Resistance against Aphids by Endophytic Bacillus velezensis YC7010 via Expressing PHYTOALEXIN DEFICIENT4 in Arabidopsis. Front. Plant Sci. 2017, 8, 211. [Google Scholar] [CrossRef]
- Bazie Tagele, S.; Woo Kim, S.; Gu Lee, H.; Seung Kim, H.; Lee, Y. Effectiveness of Multi-Trait Burkholderia Contaminans KNU17BI1 in Growth Promotion and Management of Banded Leaf and Sheath Blight in Maize Seedling. Microbiol. Res. 2018, 214, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Roslan, M.A.M.; Zulkifli, N.N.; Sobri, Z.M.; Zuan, A.T.K.; Cheak, S.C.; Abdul Rahman, N.A. Seed Biopriming with P- and K-Solubilizing Enterobacter hormaechei Sp. Improves the Early Vegetative Growth and the P and K Uptake of Okra (Abelmoschus esculentus) Seedling. PLoS ONE 2020, 15, e0232860. [Google Scholar] [CrossRef] [PubMed]
- Swamy, C.T.; Gayathri, D.; Devaraja, T.N.; Bandekar, M.; D’Souza, S.E.; Meena, R.M.; Ramaiah, N. Plant Growth Promoting Potential and Phylogenetic Characteristics of a Lichenized Nitrogen Fixing Bacterium, Enterobacter cloacae. J. Basic Microbiol. 2016, 56, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Rybakova, D.; Rack-Wetzlinger, U.; Cernava, T.; Schaefer, A.; Schmuck, M.; Berg, G. Aerial Warfare: A Volatile Dialogue between the Plant Pathogen Verticillium longisporum and Its Antagonist Paenibacillus polymyxa. Front. Plant Sci. 2017, 8, 1294. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Arora, N.K. Evaluation of Rhizospheric Pseudomonas and Bacillus as Biocontrol Tool for Xanthomonas campestris Pv Campestris. World J. Microbiol. Biotechnol. 2012, 28, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Ma, Y.; Wang, J.; Xia, Z.; Wei, H. Genomic Insights into a Plant Growth-promoting Pseudomonas koreensis Strain with Cyclic Lipopeptide-mediated Antifungal Activity. Microbiologyopen 2020, 9, e1092. [Google Scholar] [CrossRef]
- Jošić, D.; Ćirić, A.; Soković, M.; Stanojković-Sebić, A.; Pivić, R.; Lepšanović, Z.; Glamočlija, J. Antifungal Activities of Indigenous Plant Growth Promoting Pseudomonas Spp. from Alfalfa and Clover Rhizosphere. Front. Life Sci. 2015, 8, 131–138. [Google Scholar] [CrossRef]
- Jha, B.K.; Gandhi Pragash, M.; Cletus, J.; Raman, G.; Sakthivel, N. Simultaneous Phosphate Solubilization Potential and Antifungal Activity of New Fluorescent Pseudomonad Strains, Pseudomonas aeruginosa, P. plecoglossicida and P. mosselii. World J. Microbiol. Biotechnol. 2009, 25, 573–581. [Google Scholar] [CrossRef]
- Feoktistova, A.; Bakaeva, M.; Timergalin, M.; Chetverikova, D.; Kendjieva, A.; Rameev, T.; Hkudaygulov, G.; Nazarov, A.; Kudoyarova, G.; Chetverikov, S. Effects of Humic Substances on the Growth of Pseudomonas plecoglossicida 2,4-D and Wheat Plants Inoculated with This Strain. Microorganisms 2022, 10, 1066. [Google Scholar] [CrossRef]
- Volynchikova, E.; Kim, K.D. Anti-Oomycete Activity and Pepper Root Colonization of Pseudomonas plecoglossicida YJR13 and Pseudomonas putida YJR92 against Phytophthora capsici. Plant Pathol. J. 2023, 39, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Chinchilla, D.; Tan, C.; Mène-Saffrané, L.; L’Haridon, F.; Weisskopf, L. Contribution of Hydrogen Cyanide to the Antagonistic Activity of Pseudomonas Strains Against Phytophthora infestans. Microorganisms 2020, 8, 1144. [Google Scholar] [CrossRef]
- Santamaría-Hernando, S.; De Bruyne, L.; Höfte, M.; Ramos-González, M. Improvement of Fitness and Biocontrol Properties of Pseudomonas putida via an Extracellular Heme Peroxidase. Microb. Biotechnol. 2022, 15, 2652–2666. [Google Scholar] [CrossRef] [PubMed]
- Nosheen, A.; Yasmin, H.; Naz, R.; Bano, A.; Keyani, R.; Hussain, I. Pseudomonas putida Improved Soil Enzyme Activity and Growth of Kasumbha under Low Input of Mineral Fertilizers. Soil. Sci. Plant Nutr. 2018, 64, 520–525. [Google Scholar] [CrossRef]
- Hata, E.M.; Yusof, M.T.; Zulperi, D. Induction of Systemic Resistance against Bacterial Leaf Streak Disease and Growth Promotion in Rice Plant by Streptomyces shenzhenesis TKSC3 and Streptomyces Sp. SS8. Plant Pathol. J. 2021, 37, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.P.; Bastos, M.S.; Xavier, V.B.; Cassel, E.; Astarita, L.V.; Santarém, E.R. Plant Growth and Resistance Promoted by Streptomyces Spp. in Tomato. Plant Physiol. Biochem. 2017, 118, 479–493. [Google Scholar] [CrossRef]
- Papp, O.; Kocsis, T.; Biró, B.; Jung, T.; Ganszky, D.; Abod, É.; Tirczka, I.; Tóthné Bogdányi, F.; Drexler, D. Co-Inoculation of Organic Potato with Fungi and Bacteria at High Disease Severity of Rhizoctonia solani and Streptomyces Spp. Increases Beneficial Effects. Microorganisms 2021, 9, 2028. [Google Scholar] [CrossRef]
- Patel, J.K.; Madaan, S.; Archana, G. Antibiotic Producing Endophytic Streptomyces Spp. Colonize above-Ground Plant Parts and Promote Shoot Growth in Multiple Healthy and Pathogen-Challenged Cereal Crops. Microbiol. Res. 2018, 215, 36–45. [Google Scholar] [CrossRef]
- González-Pérez, E.; Ortega-Amaro, M.A.; Salazar-Badillo, F.B.; Bautista, E.; Douterlungne, D.; Jiménez-Bremont, J.F. The Arabidopsis-Trichoderma Interaction Reveals That the Fungal Growth Medium Is an Important Factor in Plant Growth Induction. Sci. Rep. 2018, 8, 16427. [Google Scholar] [CrossRef]
- Yu, C.; Fan, L.; Gao, J.; Wang, M.; Wu, Q.; Tang, J.; Li, Y.; Chen, J. The Platelet-Activating Factor Acetylhydrolase Gene Derived from Trichoderma harzianum Induces Maize Resistance to Curvularia lunata through the Jasmonic Acid Signaling Pathway. J. Environ. Sci. Health Part B 2015, 50, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, Y.; Cao, H.; Fan, Y.; Du, K.; Bu, X.; Gao, D. Effects of Trichoderma harzianum Biofertilizer on Growth, Yield, and Quality of Bupleurum chinense. Plant Direct 2022, 6, e461. [Google Scholar] [CrossRef]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma harzianum Favours the Access of Arbuscular Mycorrhizal Fungi to Non-Host Brassicaceae Roots and Increases Plant Productivity. Sci. Rep. 2019, 9, 11650. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated Application of Selenium and Silicon Enhances Growth and Anatomical Structure, Antioxidant Defense System and Yield of Wheat Grown in Salt-Stressed Soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef]
- Tripathi, R.; Keswani, C.; Tewari, R. Trichoderma koningii Enhances Tolerance against Thermal Stress by Regulating ROS Metabolism in Tomato (Solanum lycopersicum L.) Plants. J. Plant Interact. 2021, 16, 116–125. [Google Scholar] [CrossRef]
- Gajera, H.P.; Hirpara, D.G.; Katakpara, Z.A.; Patel, S.V.; Golakiya, B.A. Molecular Evolution and Phylogenetic Analysis of Biocontrol Genes Acquired from SCoT Polymorphism of Mycoparasitic Trichoderma koningii Inhibiting Phytopathogen Rhizoctonia solani Kuhn. Infect. Genet. Evol. 2016, 45, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Kafy, A.A.; Rahman, A.F.; Al Rakib, A.; Akter, K.S.; Raikwar, V.; Jahir, D.M.A.; Ferdousi, J.; Kona, M.A. Assessment and Prediction of Seasonal Land Surface Temperature Change Using Multi-Temporal Landsat Images and Their Impacts on Agricultural Yields in Rajshahi, Bangladesh. Environ. Chall. 2021, 4, 100147. [Google Scholar] [CrossRef]
- NourEldeen, N.; Mao, K.; Yuan, Z.; Shen, X.; Xu, T.; Qin, Z. Analysis of the Spatiotemporal Change in Land Surface Temperature for a Long-Term Sequence in Africa (2003–2017). Remote Sens. 2020, 12, 488. [Google Scholar] [CrossRef]
- Dash, H.R.; Mangwani, N.; Chakraborty, J.; Kumari, S.; Das, S. Marine Bacteria: Potential Candidates for Enhanced Bioremediation. Appl. Microbiol. Biotechnol. 2013, 97, 561–571. [Google Scholar] [CrossRef]
- Mahapatra, G.P.; Raman, S.; Nayak, S.; Gouda, S.; Das, G.; Patra, J.K. Metagenomics Approaches in Discovery and Development of New Bioactive Compounds from Marine Actinomycetes. Curr. Microbiol. 2020, 77, 645–656. [Google Scholar] [CrossRef]
- Lähteenmäki-Uutela, A.; Rahikainen, M.; Camarena-Gómez, M.T.; Piiparinen, J.; Spilling, K.; Yang, B. European Union Legislation on Macroalgae Products. Aquac. Int. 2021, 29, 487–509. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022. [Google Scholar]
- Parmar, P.; Kumar, R.; Neha, Y.; Srivatsan, V. Microalgae as next Generation Plant Growth Additives: Functions, Applications, Challenges and Circular Bioeconomy Based Solutions. Front. Plant Sci. 2023, 14, 1073546. [Google Scholar] [CrossRef] [PubMed]
- Slinksienė, R.; Sendzikiene, E.; Mikolaitiene, A.; Makareviciene, V.; Paleckiene, R.; Ragauskaite, D. Use of Microalgae Biomass for Production of Granular Nitrogen Biofertilizers. Green Chem. Lett. Rev. 2022, 15, 415–425. [Google Scholar] [CrossRef]
- Youssef, S.M.; El-Serafy, R.S.; Ghanem, K.Z.; Elhakem, A.; Abdel Aal, A.A. Foliar Spray or Soil Drench: Microalgae Application Impacts on Soil Microbiology, Morpho-Physiological and Biochemical Responses, Oil and Fatty Acid Profiles of Chia Plants under Alkaline Stress. Biology 2022, 11, 1844. [Google Scholar] [CrossRef] [PubMed]
- Kholssi, R.; Marks, E.A.N.; Miñón, J.; Montero, O.; Debdoubi, A.; Rad, C. Biofertilizing Effect of Chlorella sorokiniana Suspensions on Wheat Growth. J. Plant Growth Regul. 2019, 38, 644–649. [Google Scholar] [CrossRef]
- Barone, V.; Baglieri, A.; Stevanato, P.; Broccanello, C.; Bertoldo, G.; Bertaggia, M.; Cagnin, M.; Pizzeghello, D.; Moliterni, V.M.C.; Mandolino, G.; et al. Root Morphological and Molecular Responses Induced by Microalgae Extracts in Sugar Beet (Beta vulgaris L.). J. Appl. Phycol. 2018, 30, 1061–1071. [Google Scholar] [CrossRef]
- Chittora, D.; Meena, M.; Barupal, T.; Swapnil, P. Cyanobacteria as a Source of Biofertilizers for Sustainable Agriculture. Biochem. Biophys. Rep. 2020, 22, 100737. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef]
- Guedes, W.A.; Araújo, R.H.C.R.; Rocha, J.L.A.; de Lima, J.F.; Dias, G.A.; de Oliveira, Á.M.F.; de Lima, R.F.; Oliveira, L.M. Production of Papaya Seedlings Using Spirulina platensis as a Biostimulant Applied on Leaf and Root. J. Exp. Agric. Int. 2018, 28, 1–9. [Google Scholar] [CrossRef]
- Plaza, B.M.; Gómez-Serrano, C.; Acién-Fernández, F.G.; Jimenez-Becker, S. Effect of Microalgae Hydrolysate Foliar Application (Arthrospira platensis and Scenedesmus Sp.) on Petunia x Hybrida Growth. J. Appl. Phycol. 2018, 30, 2359–2365. [Google Scholar] [CrossRef]
- Carreiras, J.; Cruz-Silva, A.; Fonseca, B.; Carvalho, R.C.; Cunha, J.P.; Proença Pereira, J.; Paiva-Silva, C.; A. Santos, S.; Janeiro Sequeira, R.; Mateos-Naranjo, E.; et al. Improving Grapevine Heat Stress Resilience with Marine Plant Growth-Promoting Rhizobacteria Consortia. Microorganisms 2023, 11, 856. [Google Scholar] [CrossRef]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine Microorganisms as an Untapped Source of Bioactive Compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- Shaaban, K.A. Marine Microbial Diversity as Source of Bioactive Compounds. Mar. Drugs 2022, 20, 304. [Google Scholar] [CrossRef]
- Kapinusova, G.; Lopez Marin, M.A.; Uhlik, O. Reaching Unreachables: Obstacles and Successes of Microbial Cultivation and Their Reasons. Front. Microbiol. 2023, 14, 1089630. [Google Scholar] [CrossRef]
- Santos, A.P.; Belfiore, C.; Úrbez, C.; Ferrando, A.; Blázquez, M.A.; Farías, M.E. Extremophiles as Plant Probiotics to Promote Germination and Alleviate Salt Stress in Soybean. J. Plant Growth Regul. 2023, 42, 946–959. [Google Scholar] [CrossRef]
- Zgonik, V.; Mulec, J.; Eleršek, T.; Ogrinc, N.; Jamnik, P.; Ulrih, N.P. Extremophilic Microorganisms in Central Europe. Microorganis 2021, 9, 2326. [Google Scholar] [CrossRef]
- Jeong, S.W.; Choi, Y.J. Extremophilic Microorganisms for the Treatment of Toxic Pollutants in the Environment. Molecules 2020, 25, 4916. [Google Scholar] [CrossRef]
- Haddidi, I.; Duc, N.H.; Tonk, S.; Rápó, E.; Posta, K. Defense Enzymes in Mycorrhizal Tomato Plants Exposed to Combined Drought and Heat Stresses. Agronomy 2020, 10, 1657. [Google Scholar] [CrossRef]
- Rizvi, A.; Ahmed, B.; Khan, M.S.; Umar, S.; Lee, J. Psychrophilic Bacterial Phosphate-Biofertilizers: A Novel Extremophile for Sustainable Crop Production under Cold Environment. Microorganisms 2021, 9, 2451. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Mittler, R. Reactive Oxygen Species and Temperature Stresses: A Delicate Balance between Signaling and Destruction. Physiol. Plant 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Wang, R.; Mao, C.; Jiang, C.; Zhang, L.; Peng, S.; Zhang, Y.; Feng, S.; Ming, F. One Heat Shock Transcription Factor Confers High Thermal Tolerance in Clematis Plants. Int. J. Mol. Sci. 2021, 22, 2900. [Google Scholar] [CrossRef] [PubMed]
- Bokhari, A.; Essack, M.; Lafi, F.F.; Andres-Barrao, C.; Jalal, R.; Alamoudi, S.; Razali, R.; Alzubaidy, H.; Shah, K.H.; Siddique, S.; et al. Bioprospecting Desert Plant Bacillus Endophytic Strains for Their Potential to Enhance Plant Stress Tolerance. Sci. Rep. 2019, 9, 18154. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.; Kumar, K.; Srinivasan, T.; Kondreddy, A.; Kumar, K.R.R. An Invasive Weed-Associated Bacteria Confers Enhanced Heat Stress Tolerance in Wheat. Heliyon 2022, 8, e09893. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.M.; Kim, K.M.; Lee, I.J. Thermotolerance Effect of Plant Growth-Promoting Bacillus cereus SA1 on Soybean during Heat Stress. BMC Microbiol. 2020, 20, 175. [Google Scholar] [CrossRef] [PubMed]
- Manzanera, M. Dealing with Water Stress and Microbial Preservation. Environ. Microbiol. 2021, 23, 3351–3359. [Google Scholar] [CrossRef]
- Bhagat, N.; Raghav, M.; Dubey, S.; Bedi, N. Bacterial Exopolysaccharides: Insight into Their Role in Plant Abiotic Stress Tolerance. J. Microbiol. Biotechnol. 2021, 31, 1045–1059. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Yu, S.; Sun, M.; Yang, Z.; Gao, Z.Q. Development of Drought-Tolerant Transgenic Wheat: Achievements and Limitations. Int. J. Mol. Sci. 2019, 20, 3350. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Babar, M.A. Metabolic and Physiological Changes Induced by Plant Growth Regulators and Plant Growth Promoting Rhizobacteria and Their Impact on Drought Tolerance in Cicer arietinum L. PLoS ONE 2019, 14, e0213040. [Google Scholar] [CrossRef]
- Oren, A. Life at High Salt Concentrations, Intracellular KCl Concentrations, and Acidic Proteomes. Front. Microbiol. 2013, 4, 315. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Dopson, M. Life in Acid: PH Homeostasis in Acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Horikoshi, K. Alkaliphiles: Some Applications of Their Products for Biotechnology. Microbiol. Mol. Biol. Rev. 1999, 63, 735–750. [Google Scholar] [CrossRef]
- Xun, F.; Xie, B.; Liu, S.; Guo, C. Effect of Plant Growth-Promoting Bacteria (PGPR) and Arbuscular Mycorrhizal Fungi (AMF) Inoculation on Oats in Saline-Alkali Soil Contaminated by Petroleum to Enhance Phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G.M. Metals, Minerals and Microbes: Geomicrobiology and Bioremediation. Microbiology 2010, 156, 609–643. [Google Scholar] [CrossRef] [PubMed]
- Tang, L. Culturing Uncultivated Bacteria. Nat. Methods 2019, 16, 1078. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The Unseen Majority: Soil Microbes as Drivers of Plant Diversity and Productivity in Terrestrial Ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Schmeisser, C.; Steele, H.; Streit, W.R. Metagenomics, Biotechnology with Non-Culturable Microbes. Appl. Microbiol. Biotechnol. 2007, 75, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Amoutzias, G.; Chaliotis, A.; Mossialos, D. Discovery Strategies of Bioactive Compounds Synthesized by Nonribosomal Peptide Synthetases and Type-I Polyketide Synthases Derived from Marine Microbiomes. Mar. Drugs 2016, 14, 80. [Google Scholar] [CrossRef]
- Della Sala, G.; Hochmuth, T.; Teta, R.; Costantino, V.; Mangoni, A. Polyketide Synthases in the Microbiome of the Marine Sponge Plakortis halichondrioides: A Metagenomic Update. Mar. Drugs 2014, 12, 5425–5440. [Google Scholar] [CrossRef]
- Russo, P.; Kisialiou, A.; Lamonaca, P.; Moroni, R.; Prinzi, G.; Fini, M. New Drugs from Marine Organisms in Alzheimer’s Disease. Mar. Drugs 2015, 14, 5. [Google Scholar] [CrossRef]
- Davidson, S.K.; Allen, S.W.; Lim, G.E.; Anderson, C.M.; Haygood, M.G. Evidence for the Biosynthesis of Bryostatins by the Bacterial Symbiont “ Candidatus Endobugula Sertula” of the Bryozoan Bugula neritina. Appl. Environ. Microbiol. 2001, 67, 4531–4537. [Google Scholar] [CrossRef]
- Sharuddin, S.S.; Ramli, N.; Yusoff, M.Z.M.; Muhammad, N.A.N.; Ho, L.S.; Maeda, T. Advancement of Metatranscriptomics towards Productive Agriculture and Sustainable Environment: A Review. Int. J. Mol. Sci. 2022, 23, 3737. [Google Scholar] [CrossRef]
- Sham, A.; Al-Ashram, H.; Whitley, K.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Metatranscriptomic Analysis of Multiple Environmental Stresses Identifies RAP2.4 Gene Associated with Arabidopsis Immunity to Botrytis cinerea. Sci. Rep. 2019, 9, 17010. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Mattupalli, C.; Eversole, K.; Leach, J.E. Enabling Sustainable Agriculture through Understanding and Enhancement of Microbiomes. New Phytol. 2021, 230, 2129–2147. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, M.; Bastida, F.; Sciarrillo, R.; Guarino, C. Soil Metaproteomics for the Study of the Relationships Between Microorganisms and Plants: A Review of Extraction Protocols and Ecological Insights. Int. J. Mol. Sci. 2020, 21, 8455. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, H.; Zhang, Z.; Lin, R.; Zhang, Z.; Lin, W. Comparative Metaproteomic Analysis on Consecutively Rehmannia glutinosa-Monocultured Rhizosphere Soil. PLoS ONE 2011, 6, e20611. [Google Scholar] [CrossRef] [PubMed]
- Schlaeppi, K.; Bulgarelli, D. The Plant Microbiome at Work. Mol. Plant-Microbe Interact. 2015, 28, 212–217. [Google Scholar] [CrossRef]
- Flores-Duarte, N.J.; Pajuelo, E.; Mateos-Naranjo, E.; Navarro-Torre, S.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S.; Carrasco López, J.A. A Culturomics-Based Bacterial Synthetic Community for Improving Resilience towards Arsenic and Heavy Metals in the Nutraceutical Plant Mesembryanthemum Crystallinum. Int. J. Mol. Sci. 2023, 24, 7003. [Google Scholar] [CrossRef]
- Douds, D.D.; Nagahashi, G.; Reider, C.; Hepperly, P.R. Inoculation with Arbuscular Mycorrhizal Fungi Increases the Yield of Potatoes in a High P Soil. Biol. Agric. Hortic. 2007, 25, 67–78. [Google Scholar] [CrossRef]
- Goetten, L.C.; Moretto, G.; Stürmer, S.L. Influence of Arbuscular Mycorrhizal Fungi Inoculum Produced On-Farm and Phosphorus on Growth and Nutrition of Native Woody Plant Species from Brazil. Acta Bot. Bras. 2016, 30, 9–16. [Google Scholar] [CrossRef]
- Douds, D.D.; Carr, E.; Shenk, J.E.; Ganser, S. Positive Yield Response of Eggplant (Solanum melongena L.) to Inoculation with AM Fungi Produced on-Farm. Sci. Hortic. 2017, 224, 48–52. [Google Scholar] [CrossRef]
- Dass, A.; Jinger, D.; Kaur, R.; Kumar, S.V.; Kumari, K. Addressing Multinutrient Deficiency in Crops Using Customized Fertilizer. Indian Farming 2017, 67, 22–25. [Google Scholar]
- Pacheco-Aguirre, J.A.; Ruíz-Sánchez, E.; Ballina-Gómez, H.S.; Alvarado-López, C.J. Does Polymer-Based Encapsulation Enhance Performance of Plant Growth Promoting Microorganisms? A Meta-Analysis View. Agrociencia 2017, 51, 173–187. [Google Scholar]
- Sohaib, M.; Zahir, Z.A.; Khan, M.Y.; Ans, M.; Asghar, H.N.; Yasin, S.; Al-Barakah, F.N.I. Comparative Evaluation of Different Carrier-Based Multi-Strain Bacterial Formulations to Mitigate the Salt Stress in Wheat. Saudi J. Biol. Sci. 2020, 27, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Singh, I. Microbial Biofertilizers: Types and Applications. In Biofertilizers for Sustainable Agriculture and Environment; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–19. [Google Scholar]
- John, R.P.; Tyagi, R.D.; Brar, S.K.; Surampalli, R.Y.; Prévost, D. Bio-Encapsulation of Microbial Cells for Targeted Agricultural Delivery. Crit. Rev. Biotechnol. 2011, 31, 211–226. [Google Scholar] [CrossRef]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Chenari Bouket, A.; Alenezi, F.N.; Belbahri, L. Recent Advances in Encapsulation Techniques of Plant Growth-Promoting Microorganisms and Their Prospects in the Sustainable Agriculture. Appl. Sci. 2022, 12, 9020. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in Agriculture: An Overview on Concepts, Strategies and Effects on Soil Microorganisms. In Advances in Agronomy; Springer: Berlin/Heidelberg, Germany, 2020; pp. 31–87. [Google Scholar]
- Gewin, V. How Peat Could Protect the Planet. Nature 2020, 578, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Thapa, S.; Geat, N.; Mehriya, M.L.; Rajawat, M.V.S. Biofertilizers: Mechanisms and Application. In Biofertilizers; Elsevier: Amsterdam, The Netherlands, 2021; pp. 151–166. [Google Scholar]
- Lane, N. The Unseen World: Reflections on Leeuwenhoek (1677) ‘Concerning Little Animals’. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140344. [Google Scholar] [CrossRef] [PubMed]
- Fred, E.; Baldwin, I.; McCoy, E. Root Nodule Bacteria and Leguminous Plants; UW-Madison Libraries Parallel Press: Madison, WI, USA, 1932. [Google Scholar]
- van Schreven, D.A.; Harmsen, G.W.; Lindenbergh, D.J.; Otzen, D. Experiments on the Cultivation of Rhizobium in Liquid Media for Use on the Zuiderzee Polders. Antonie Van Leeuwenhoek 1953, 19, 300–308. [Google Scholar] [CrossRef]
- Dommergues, Y.R.; Diem, H.G.; Divies, C. Polyacrylamide-Entrapped Rhizobium as an Inoculant for Legumes. Appl. Environ. Microbiol. 1979, 37, 779–781. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-Living Bacterial Inocula for Enhancing Crop Productivity. Trends Biotechnol. 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Singleton, P.W.; Keyser, H.; Sande, E. Development and Evaluation of Liquid Inoculants. In Inoculants and Nitrogen Fixation of Legumes in Vietnam; Herridge, D., Ed.; CSIRO: Canberra, Australia, 2022; Volume 109, pp. 52–66. [Google Scholar]
- Li, Z. Nano Biofertilizer Production Technology and Preparation Method. CN102190523A, 17 March 2010. [Google Scholar]
- Singh, G.; Paithankar, I. Encapsulation of Biofertilizers, Biopesticides and Biocontrol Agents. In Sustainable Agriculture Reviews; Singh, N.K., Chattopadhyay, A., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2023; pp. 121–150. [Google Scholar]
- Sudheer, S.; Bai, R.G.; Usmani, Z.; Sharma, M. Insights on Engineered Microbes in Sustainable Agriculture: Biotechnological Developments and Future Prospects. Curr. Genom. 2020, 21, 321–333. [Google Scholar] [CrossRef]
- Young, C.; Rekha, P.D.; Lai, W.; Arun, A.B. Encapsulation of Plant Growth-promoting Bacteria in Alginate Beads Enriched with Humic Acid. Biotechnol. Bioeng. 2006, 95, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Saberi-Rise, R.; Moradi-Pour, M. The Effect of Bacillus subtilis Vru1 Encapsulated in Alginate—Bentonite Coating Enriched with Titanium Nanoparticles against Rhizoctonia solani on Bean. Int. J. Biol. Macromol. 2020, 152, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Panichikkal, J.; Prathap, G.; Nair, R.A.; Krishnankutty, R.E. Evaluation of Plant Probiotic Performance of Pseudomonas Sp. Encapsulated in Alginate Supplemented with Salicylic Acid and Zinc Oxide Nanoparticles. Int. J. Biol. Macromol. 2021, 166, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Amiet-Charpentier, C.; Gadille, P.; Benoit, J.P. Rhizobacteria Microencapsulation: Properties of Microparticles Obtained by Spray-Drying. J. Microencapsul. 1999, 16, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanasambandan, T. Advances and Trends in Nano-Biofertilizers. SSRN Electron. J. 2018. [Google Scholar] [CrossRef]
- Akhtar, N.; Ilyas, N.; Meraj, T.A.; Pour-Aboughadareh, A.; Sayyed, R.Z.; Mashwani, Z.-R.; Poczai, P. Improvement of Plant Responses by Nanobiofertilizer: A Step towards Sustainable Agriculture. Nanomaterials 2022, 12, 965. [Google Scholar] [CrossRef]
- Garg, D.; Sridhar, K.; Stephen Inbaraj, B.; Chawla, P.; Tripathi, M.; Sharma, M. Nano-Biofertilizer Formulations for Agriculture: A Systematic Review on Recent Advances and Prospective Applications. Bioengineering 2023, 10, 1010. [Google Scholar] [CrossRef]
- Li, C.-C.; Dang, F.; Li, M.; Zhu, M.; Zhong, H.; Hintelmann, H.; Zhou, D.-M. Effects of Exposure Pathways on the Accumulation and Phytotoxicity of Silver Nanoparticles in Soybean and Rice. Nanotoxicology 2017, 11, 699–709. [Google Scholar] [CrossRef]
- Alshaal, T.; El-Ramady, H. Foliar Application: From Plant Nutrition to Biofortification. Environ. Biodivers. Soil Secur. 2017, 1, 71–83. [Google Scholar] [CrossRef]
- Hong, J.; Wang, C.; Wagner, D.C.; Gardea-Torresdey, J.L.; He, F.; Rico, C.M. Foliar Application of Nanoparticles: Mechanisms of Absorption, Transfer, and Multiple Impacts. Environ. Sci. Nano 2021, 8, 1196–1210. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M.; et al. A Combined Phenotypic and Metabolomic Approach for Elucidating the Biostimulant Action of a Plant-Derived Protein Hydrolysate on Tomato Grown Under Limited Water Availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef] [PubMed]
- Filipczak, J.; Żurawicz, E.; Paszt, L.S. Influence of Selected Biostimulants on the Growth and Yielding of “Elkat” Strawberry Plants. Zesz. Nauk. Inst. Ogrod. 2016, 24, 43–58. [Google Scholar]
- Oancea, F.; Velea, S.; Fatu, V.; Mincea, C.; Ilie, L. Micro-Algae Based Plant Biostimulant and Its Effect on Water Stressed Tomato Plants. Rom. J. Plant Prot. 2013, 6, 104–117. [Google Scholar]
- López-Padrón, I.; Martínez-González, L.; Pérez-Domínguez, G.; Cedeño-Rodríguez, L.; Reyes-Guerrero, Y.; Cárdenas-Travieso, R.M.; Núñez-Vázquez, M.; Cabrera-Rodríguez, J.A. Effects of Bioactive Products on Cicer arietinum L. Plants. Cultiv. Trop. 2021, 42, e06. [Google Scholar]
- Dias, G.A.; Rocha, R.H.C.; Araújo, J.L.; de Lima, J.F.; Guedes, W.A. Growth, Yield, And Postharvest Quality In Eggplant Produced Under Different Foliar Fertilizer (Spirulina platensis) Treatments. Semin. Cienc. Agrarias. Univ. Estadual De Londrina 2016, 37, 3893–3902. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Weselowski, B. Influence of Foliar Sprays of Bacillus subtilis QST 713 on Development of Early Blight Disease and Yield of Field Tomatoes in Ontario. Can. J. Plant Pathol. 2014, 36, 170–178. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Weselowski, B. Efficacy of Bacillus subtilis QST 713 Formulations, Copper Hydroxide, and Their Tank Mixes on Bacterial Spot of Tomato. Crop Prot. 2015, 74, 70–76. [Google Scholar] [CrossRef]
- El Habbasha, E.S.; Mekhail Tawkik, M.; El Kramany, M.F. Comparative Efficacy of Different Bio-Chemical Foliar Applications on Growth, Yield and Yield Attributes of Some Wheat Cultivars. World J. Agric. Sci. 2013, 9, 345–353. [Google Scholar]
- Costa, R.R.G.F.; Quirino, G.D.S.F.; Naves, D.C.D.F.; Santos, C.B.; Rocha, A.F.D.S. Efficiency of Inoculant with Azospirillum brasilense on the Growth and Yield of Second-Harvest Maize. Pesqui. Agropecu. Trop. 2015, 45, 304–311. [Google Scholar] [CrossRef]
- Preininger, C.; Sauer, U.; Bejarano, A.; Berninger, T. Concepts and Applications of Foliar Spray for Microbial Inoculants. Appl. Microbiol. Biotechnol. 2018, 102, 7265–7282. [Google Scholar] [CrossRef]
- Abbate, C.; Scavo, A.; Pesce, G.R.; Fontanazza, S.; Restuccia, A.; Mauromicale, G. Soil Bioplastic Mulches for Agroecosystem Sustainability: A Comprehensive Review. Agriculture 2023, 13, 197. [Google Scholar] [CrossRef]
- Huang, F.; Zhang, Q.; Wang, L.; Zhang, C.; Zhang, Y. Are Biodegradable Mulch Films a Sustainable Solution to Microplastic Mulch Film Pollution? A Biogeochemical Perspective. J. Hazard. Mater. 2023, 459, 132024. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, D.R. Morphology and Ecology of Lithic-Mulch Agriculture. Geogr. Rev. 1994, 84, 172. [Google Scholar] [CrossRef]
- Vinci, G.; Ruggieri, R.; Billi, A.; Pagnozzi, C.; Di Loreto, M.V.; Ruggeri, M. Sustainable Management of Organic Waste and Recycling for Bioplastics: A LCA Approach for the Italian Case Study. Sustainability 2021, 13, 6385. [Google Scholar] [CrossRef]
- Skoczinski, P.; Carus, M.; de Guzman, D.; Käb, H.; Chinthapalli, R.; Ravenstijn, J.; Baltus, W.; Raschka, A. Bio-Based Building Blocks and Polymers—Global Capacities, Production and Trends 2020–2025; Language: Hürt, Germany, 2021. [Google Scholar]
- ASTM D5488-94DE1; Standard Terminology of Environmental Labeling of Packaging Materials and Packages. ASTM: West Conshohocken, PA, USA, 2002.
- Mansoor, Z.; Tchuenbou-Magaia, F.; Kowalczuk, M.; Adamus, G.; Manning, G.; Parati, M.; Radecka, I.; Khan, H. Polymers Use as Mulch Films in Agriculture—A Review of History, Problems and Current Trends. Polymers 2022, 14, 5062. [Google Scholar] [CrossRef]
- Lasmini, S.A.; Idham; Pagiu, S.; Yusuf, R.; Hayati, N.; Yunus, M.; Pasaru, F.; Nasir, B.H.; Rosmini; Khasanah, N.; et al. Application of Mulch and Soil Microbes to Increase Growth and Yield of Chili Pepper. AIP Conf. Proc. 2023, 2606, 040001. [Google Scholar] [CrossRef]
- Kellenberger, E. Exploring the Unknown. EMBO Rep. 2001, 2, 5–7. [Google Scholar] [CrossRef]
- Bodor, A.; Bounedjoum, N.; Vincze, G.E.; Erdeiné Kis, Á.; Laczi, K.; Bende, G.; Szilágyi, Á.; Kovács, T.; Perei, K.; Rákhely, G. Challenges of Unculturable Bacteria: Environmental Perspectives. Rev. Environ. Sci. Biotechnol. 2020, 19, 1–22. [Google Scholar] [CrossRef]
- Velasco-Rodríguez, Ó.; Fil, M.; Heggeset, T.M.B.; Degnes, K.F.; Becerro-Recio, D.; Kolsaková, K.; Haugen, T.; Jønsson, M.; Toral-Martínez, M.; García-Estrada, C.; et al. Characterization of Microbial Diversity in Decayed Wood from a Spanish Forest: An Environmental Source of Industrially Relevant Microorganisms. Microorganisms 2022, 10, 1249. [Google Scholar] [CrossRef]
- López-Jiménez, A.T.; Mostowy, S. Emerging Technologies and Infection Models in Cellular Microbiology. Nat. Commun. 2021, 12, 6764. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Gaudin, A.C.M. What Is the Agronomic Potential of Biofertilizers for Maize? A Meta-Analysis. FEMS Microbiol. Ecol. 2018, 94, 94. [Google Scholar] [CrossRef] [PubMed]
- Agake, S.I.; Ohwaki, Y.; Kojima, K.; Yoshikawa, E.; Artigas Ramirez, M.D.; Bellingrath-Kimura, S.D.; Yamada, T.; Ookawa, T.; Ohkama-Ohtsu, N.; Yokoyama, T. Biofertilizer with Bacillus pumilus TUAT1 Spores Improves Growth, Productivity, and Lodging Resistance in Forage Rice. Agronomy 2022, 12, 2325. [Google Scholar] [CrossRef]
- Audu, I.G.; Barde, A.; Yila, O.M.; Onwualu, P.A.; Lawal, B.M. Exploring Biogas and Biofertilizer Production from Abattoir Wastes in Nigeria Using a Multi-Criteria Assessment Approach. Recycling 2020, 5, 18. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Sevilla-Morán, B.; Sandín-España, P.; López-Goti, C.; Alonso-Prados, J.L. Biopesticides in the Framework of the European Pesticide Regulation (EC) No. 1107/2009. Pest Manag. Sci. 2014, 70, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Tur-Cardona, J.; Bonnichsen, O.; Speelman, S.; Verspecht, A.; Carpentier, L.; Debruyne, L.; Marchand, F.; Jacobsen, B.H.; Buysse, J. Farmers’ Reasons to Accept Bio-Based Fertilizers: A Choice Experiment in Seven Different European Countries. J. Clean. Prod. 2018, 197, 406–416. [Google Scholar] [CrossRef]
- Daniel, A.I.; Fadaka, A.O.; Gokul, A.; Bakare, O.O.; Aina, O.; Fisher, S.; Burt, A.F.; Mavumengwana, V.; Keyster, M.; Klein, A. Biofertilizer: The Future of Food Security and Food Safety. Microorganisms 2022, 10, 1220. [Google Scholar] [CrossRef] [PubMed]
- Hoehnel, A.; Zannini, E.; Arendt, E.K. Targeted Formulation of Plant-Based Protein-Foods: Supporting the Food System’s Transformation in the Context of Human Health, Environmental Sustainability and Consumer Trends. Trends Food Sci. Technol. 2022, 128, 238–252. [Google Scholar] [CrossRef]
- Gracia, A.; Gómez, M.I. Food Sustainability and Waste Reduction in Spain: Consumer Preferences for Local, Suboptimal, And/Or Unwashed Fresh Food Products. Sustainability 2020, 12, 4148. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, T.A. Future of Bio-Fertilizers in Indian Agriculture: An Overview. Int. J. Agric. Food Res. 2014, 3, 10–23. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Ploll, U.; Arato, M.; Börner, J.; Hartmann, M. Sustainable Innovations: A Qualitative Study on Farmers’ Perceptions Driving the Diffusion of Beneficial Soil Microbes in Germany and the UK. Sustainability 2022, 14, 5749. [Google Scholar] [CrossRef]
- Rai, P.K.; Rai, A.; Sharma, N.K.; Singh, T.; Kumar, Y. Limitations of Biofertilizers and Their Revitalization through Nanotechnology. J. Clean. Prod. 2023, 418, 138194. [Google Scholar] [CrossRef]
- Official Journal of the European Union. Regulation of the European Parliament and of the Council Laying down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32019R1009 (accessed on 5 November 2023).
- Malusá, E.; Vassilev, N. A Contribution to Set a Legal Framework for Biofertilisers. Appl. Microbiol. Biotechnol. 2014, 98, 6599–6607. [Google Scholar] [CrossRef] [PubMed]
- Raimi, A.; Roopnarain, A.; Chirima, G.J.; Adeleke, R. Insights into the Microbial Composition and Potential Efficiency of Selected Commercial Biofertilisers. Heliyon 2020, 6, e04342. [Google Scholar] [CrossRef]
- Praveen, K.V.; Singh, A. Realizing the Potential of a Low-Cost Technology to Enhance Crop Yields: Evidence from a Meta-Analysis of Biofertilizers in India. Agric. Econ. Res. Rev. 2019, 32, 77. [Google Scholar] [CrossRef]
- Suh, J.S.; Jiarong, P.; Toan, P.V. Quality Control of Biofertilizers. In Biofertilizers Manual; Forum for Nuclear Cooperation in Asia: Tokyo, Japan, 2006; pp. 112–115. [Google Scholar]



| Scientific and Technological | Challenges in the Isolation of New Microbial Species | |
|---|---|---|
| Environmental | Biotic | Negative interactions with resident microbiome (e.g., competition or predation) and different effect depending on the crop. |
| Abiotic | Variations in physicochemical soil properties (e.g., nutrients, moisture, temperature). Interaction with other agricultural components (amendments, chemical fertilizers, pesticides, etc.). | |
| Practical | Social aspects | Added value to the product from the consumers perspective may promote its application among farmers. |
| Accessibility | Products with limited versatility, shelf-life and adaptability to different agricultural practices. Limited knowledge of manipulation and application by farmers. | |
| Regulations | Lack of standardized and universal protocols and guidelines. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibáñez, A.; Garrido-Chamorro, S.; Vasco-Cárdenas, M.F.; Barreiro, C. From Lab to Field: Biofertilizers in the 21st Century. Horticulturae 2023, 9, 1306. https://doi.org/10.3390/horticulturae9121306
Ibáñez A, Garrido-Chamorro S, Vasco-Cárdenas MF, Barreiro C. From Lab to Field: Biofertilizers in the 21st Century. Horticulturae. 2023; 9(12):1306. https://doi.org/10.3390/horticulturae9121306
Chicago/Turabian StyleIbáñez, Ana, Sonia Garrido-Chamorro, María F. Vasco-Cárdenas, and Carlos Barreiro. 2023. "From Lab to Field: Biofertilizers in the 21st Century" Horticulturae 9, no. 12: 1306. https://doi.org/10.3390/horticulturae9121306
APA StyleIbáñez, A., Garrido-Chamorro, S., Vasco-Cárdenas, M. F., & Barreiro, C. (2023). From Lab to Field: Biofertilizers in the 21st Century. Horticulturae, 9(12), 1306. https://doi.org/10.3390/horticulturae9121306