


Exogenous Melatonin Application Induced Morpho-Physiological and Biochemical Regulations Conferring Salt Tolerance in Ranunculus asiaticus L.



Abstract
:1. Introduction
2. Materials and Methods
2.1. Material, Cultivation Conditions, and Trial Layout
2.2. Salinity Treatment and MT Application
2.3. Morphological Characteristics
2.4. Physiological and Biochemical Assesments
2.5. Statical Analysis
3. Results
3.1. The Effect of Melatonin on Plant Morphology under Salinity Stress
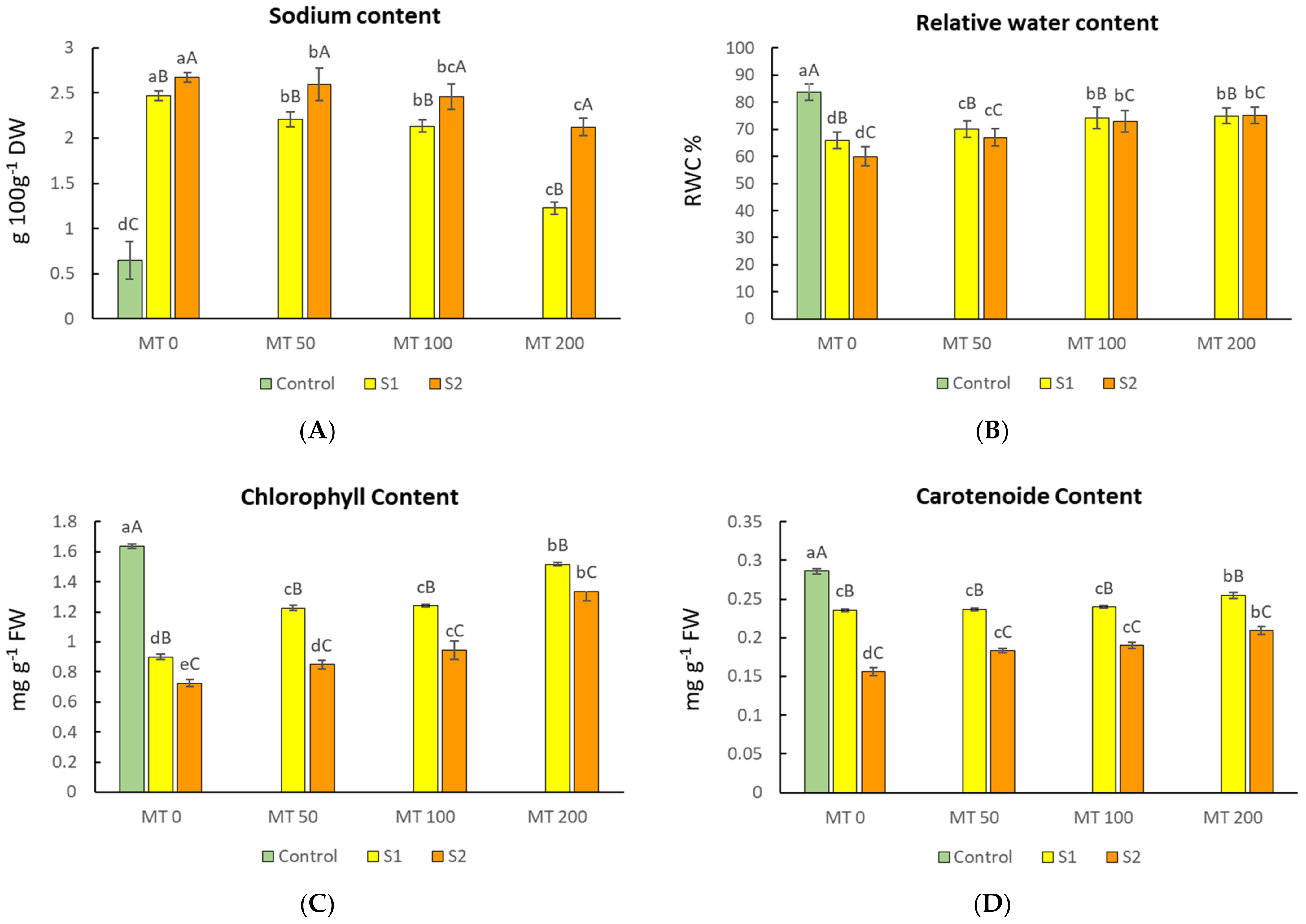
3.2. Changes in Sodium Ion Level and Relative Water Content (RWC) of the Leaves
3.3. Changes in Photosynthetic Pigments of Leaves
3.4. Changes in Electrolyte Leakage and Proline Content
3.5. Changes in Peroxidase Enzyme Activity (POD)
3.6. Changes in Flower Bud Emergence
3.7. Salt Tolerance Index (STI)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity Stress Alleviation Using Arbuscular Mycorrhizal Fungi: A Review. Agron. Sust. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.; Drechsel, P.; Noble, A. Economics of Salt-Induced Land Degradation and Restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Bui, E. Soil Salinity: A Neglected Factor in Plant Ecology and Biogeography. J. Arid Env. 2013, 92, 14–25. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boughattas, S.; Hu, S.; Oh, S.; Sa, T. A Meta-Analysis of Arbuscular Mycorrhizal Effects on Plants Grown under Salt Stress. Mycorrhiza 2014, 24, 611–625. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium Transport and Plant Salt Tolerance. Physiol. Plant 2007, 133, 651–669. [Google Scholar] [CrossRef]
- Pandolfi, C.; Mancuso, S.; Shabala, S. Physiology of Acclimation to Salinity Stress in Pea (Pisum sativum). Env. Exp. Bot. 2012, 84, 44–55. [Google Scholar] [CrossRef]
- Carter, C.T.; Grieve, C.M. Mineral Nutrition, Growth, and Germination of Antirrhinum majus L. (Snapdragon) When Produced Under Increasingly Saline Conditions. HortScience 2008, 43, 710–718. [Google Scholar] [CrossRef]
- Grieve, C.M. Salinity-Induced Enhancement of Horticultural Crop Quality. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Taylor and Francis: Boca Raton, FL, USA, 2011; pp. 1173–1194. [Google Scholar]
- Grieve, C.M.; Poss, J.A.; Shouse, P.J.; Carter, C.T. Modeling Growth of Matthiola Incana in Response to Saline Wastewaters Differing in Nitrogen Level. HortScience 2008, 43, 787–1793. [Google Scholar] [CrossRef]
- Shillo, R.; Ding, M.; Pasternak, D.; Zaccai, M. Cultivation of Cut Flower and Bulb Species with Saline Water. Sci. Hort. 2002, 92, 41–54. [Google Scholar] [CrossRef]
- De Hertogh, A.A. Holland Bulb Forcer’s Guide, 5th ed.; Alkemade Printing BV: Lisse, The Netherlands, 1996. [Google Scholar]
- Parkin, J. The Glossy Petals of Ranunculus. Ann. Bot. 1928, 4, 739–755. [Google Scholar] [CrossRef]
- Karlsson, M. Producing Ravishing Ranunculus; Greenhouse Product News: Sparta, MI, USA, 2003; pp. 44–48. Available online: https://gpnmag.com/article/producing-ravishing-ranunculus/ (accessed on 1 November 2022).
- Margherita, B.; Giampiero, C.; Pierre, D. Field Performance of Tissue-Cultured Plants of Ranunculus asiaticus L. Sci. Hortic. 1996, 66, 229–239. [Google Scholar] [CrossRef]
- Meynet, J. Ranunculus. In Physiology of Flower Bulbs: A Comprehensive Treatise on the Physiology and Utilization of Ornamental Flowering Bulbous and Tuberous Plants; De Hertogh, A.A., Le Nard, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 603–610. [Google Scholar]
- Horovitz, A. Ranunculus. In Handbook of Flowering; Halevy, A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; Volume 4, pp. 155–161. [Google Scholar]
- Rauter, S.; Sun, Y.; Stock, M. Visual Quality, Gas Exchange, and Yield of Anemone and Ranunculus Irrigated with Saline Water. Horttechnology 2021, 31, 763–770. [Google Scholar] [CrossRef]
- Valdez-Aguilar, L.A.; Grieve, C.M.; Poss, J.; Mellano, M.A. Hypersensitivity of Ranunculus asiaticus to Salinity and Alkaline PH in Irrigation Water in Sand Cultures. HortScience 2009, 44, 138–144. [Google Scholar] [CrossRef]
- Kamran, M.; Wennan, S.; Ahmad, I.; Xiangping, M.; Wenwen, C.; Xudong, Z.; Siwei, M.; Khan, A.; Qingfang, H.; Tiening, L. Application of Paclobutrazol Affect Maize Grain Yield by Regulating Root Morphological and Physiological Characteristics under a Semi-Arid Region. Sci. Rep. 2018, 1, 15. [Google Scholar]
- Qingbo, K.; Jun, Y.; Bomei, W.; Jianhong, R.; Lina, Y.; Xiping, D.; Shiwen, W. Melatonin Mitigates Salt Stress in Wheat Seedlings by Modulating Polyamine Metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar]
- Mishima, K. Melatonin as a Regulator of Human Sleep and Circadian Systems. Nihon. Rinsho. Jpn. J. Clin. Med. 2012, 70, 1139–1144. [Google Scholar]
- Hardeland, R.; Madrid, J.; Tan, D.-X.; Reiter, R. Melatonin, the Circadian Multioscillator System and Health: The Need for Detailed Analyses of Peripheral Melatonin Signaling. J. Pineal Res. 2012, 52, 139–166. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of Antioxidant Enzymes: A Significant Role for Melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Barrett, P.; Bolborea, M. Molecular Pathways Involved in Seasonal Body Weight and Reproductive Responses Governed by Melatonin. J. Pineal Res. 2012, 52, 376–388. [Google Scholar] [CrossRef]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of Melatonin in Plants and Its Effects on Plasma Melatonin Levels and Binding to Melatonin Receptors in Vertebrates. Biochem. Mol. Boil. Int. 1995, 35, 627–634. [Google Scholar]
- Tan, D.; Manchester, L.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Fundamental Issues Related to the Origin of Melatonin and Melatonin Isomers during Evolution: Relation to Their Biological Functions. Int. J. Mol. Sci. 2012, 15, 15858–15890. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Y.L.; Liang, D.; Li, C.; Ma, F.; Yue, Z. Delayed Senescence of Apple Leaves by Exogenous Melatonin Treatment: Toward Regulating the Ascorbate–Glutathione Cycle. J. Pineal Res. 2012, 53, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.; Guo, Y.D. Melatonin Promotes Seed Germination under High Salinity by Regulating Antioxidant Systems, ABA and GA4 Interaction in Cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Ahmad, S.; Cui, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Su, W.; Javed, T.; El Hamed, A.; Zhikuan, S.; et al. Exogenous Application of Melatonin Induces Tolerance to Salt Stress by Improving the Photosynthetic Efficiency and Antioxidant Defense System of Maize Seedling. J. Plant Growth Regul. 2021, 40, 1270–1283. [Google Scholar] [CrossRef]
- Yin, L.; Wang, P.; Li, M.; Ke, X.; Li, C.; Liang, D.; Wu, S.; Ma, X.; Li, C.; Zou, Y. Exogenous Melatonin Improves Malus Resistance to Marssonina Apple Blotch. J. Pineal Res. 2013, 54, 426–434. [Google Scholar] [CrossRef]
- Szafrańska, K.; Reiter, R.J.; Posmyk, M.M. Melatonin Application to Pisum sativum L. Seeds Positively Influences the Function of the Photosynthetic Apparatus in Growing Seedlings during Paraquat-Induced Oxidative Stress. Front. Plant Sci. 2016, 7, 1663. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of Melatonin in Plants: A Review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative Physiological, Metabolomic, and Transcriptomic Analyse Sreveal Mechanisms of Improved Abiotic Stress Resistance in Bermudagrass [Cynodon dactylon (L). Pers.] by Exogenous Melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Chen, H.; Wang, Z.; Gu, X.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X.; et al. Exogenous Melatonin Confers Salt Stress Tolerance to Watermelon by Improving Photosynthesis and Redox Homeostasis. Front. Plant Sci. 2017, 8, 295. [Google Scholar] [CrossRef]
- Wang, L.Y.; Liu, J.L.; Wang, W.X.; Sun, Y. Exogenous Melatonin Improves Growth and Photosynthetic Capacity of Cucumber under Salinity-Induced Stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- Castañares, J.L.; Bouzo, C.A. Effect of Exogenous Melatonin on Seed Germination and Seedling Growth in Melon (Cucumis melo L.) Under Salt Stress. Hortic. Plant J. 2019, 5, 79–87. [Google Scholar] [CrossRef]
- Bahcesular, B.; Yildirim, E.D.; Karaçocuk, M.; Kulak, M.; Karaman, S. Seed Priming with Melatonin Effects on Growth, Essential Oil Compounds and Antioxidant Activity of Basil (Ocimum basilicum L.) under Salinity Stress. Ind. Crops Prod. 2020, 146, 112165. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Yang, S.J.; Chen, Y.Y. Effects of Melatonin on Photosynthetic Performance and Antioxidants in Melon during Cold and Recovery. Biol. Plant. 2017, 61, 571–578. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Vander, W.J.; Sabbatini, P. Exogenous Melatonin Improves Glutathione Content, Redox State and Increases Essential Oil Production in Two Salvia Species under Drought Stress. Sci. Rep. 2020, 10, 6883. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S. Melatonin Alleviates Cold-Induced Oxidative Damage in Maize Seedlings by up-Regulating Mineral Elements and Enhancing Antioxidant Activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, I. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Drought-Stress-Induced Changes in Activities of Superoxide Dismutase, Catalase, and Peroxidase in Wheat Species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Korkmaz, D. Precipitation Titration: Determination of Chloride by the Mohr Method. Methods 2017, 4, 1–6. [Google Scholar]
- Sbei, H.; Shehzad, T.; Harrabi, M.; Okuno, K. Salinity Tolerance Evaluation of Asian Barley Accessions (Hordeum vulgare L.) at the Early Vegetative Stage. J. Arid L. Stud. 2014, 24, 183–186. [Google Scholar]
- West, S.G.; Finch, J.F.; Curran, P.J. Structural Equation Models with Nonnormal Variables: Problems and Remedies. In Structural Equation Modeling: Concepts, Issues, and Applications; Hoyle, R.H., Ed.; Sage Publications, Inc.: New York, NY, USA, 1995; pp. 56–75. [Google Scholar]
- Brown, M.B.; Forsythe, A.B. Robust Tests for the Equality of Variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Garson, G.D. Testing Statistical Assumptions; Statistical Associates Publishing: Asheboro, NC, USA, 2012. [Google Scholar]
- Tabachnick, B.; Fidell, L. IBM SPSS Statistics for Windows. In Using Multivar. Stat. Pearson, Boston. IBM Corp. Released 2020. IBM SPSS Stat. Wind. 27.0.; IBM Corp.: Armonk, NY, USA, 2013. [Google Scholar]
- IBM Corporation. Released 2020. IBM SPSS Statistics for Windows, Version 27.0.; IBM Corp.: Armonk, NY, USA, 2020. [Google Scholar]
- Olson, C.L. On Choosing a Test Statistic in Multivariate Analysis of Variance. Psychol. Bull. 1976, 83, 579. [Google Scholar] [CrossRef]
- Barbara, G.T.; Linda, S.F. Using Multivariate Statistics, 7th ed.; Pearson: New York, NY, USA, 2013; ISBN-13: 9780135350904; ISBN-10: 0134790545. [Google Scholar]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.; Villar, R. Causes and Consequences of Variation in Leaf Mass per Area (LMA): A Meta-Analysis. New Phytol. 2010, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.B.A.J.H.; Arnao, M.B.; Hernández-Ruiz, J. Melatonin in Flowering, Fruit Set and Fruit Ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef]
- Alyammahi, O.; Gururani, M.A. Chlorophyll-a Fluorescence Analysis Reveals Differential Response of Photosynthetic Machinery in Melatonin-Treated Oat Plants Exposed to Osmotic Stress. Agronomy 2020, 10, 1520. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abdalla, A.M.; Abd Elhamid, E.M.; Ezzo, M.I. Role of Melatonin in Improving Growth, Yield Quantity and Quality of Moringa oleifera L. Plant under Drought Stress. Bull. Natl. Res. Cent. 2020, 44, 18. [Google Scholar] [CrossRef]
- Han, Q.; Huang, B.; Ding, C.; Zhang, Z.; Chen, Y.; Hu, C.; Zhou, L.; Huang, Y.; Liao, J.; Yuan, S.; et al. Osystem II in Cold-Stressed Rice Seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef]
- Kamiab, F. Exogenous Melatonin Mitigates the Salinity Damages and Improves the Growth of Pistachio under Salinity Stress. J. Plant Nutr. 2020, 43, 1468–1484. [Google Scholar] [CrossRef]
- Imran, M.; Latif Khan, A.; Shahzad, R.; Aaqil Khan, M.; Bilal, S.; Khan, A.; Kang, S.; Lee, I. Exogenous Melatonin Induces Drought Stress Tolerance by Promoting Plant Growth and Antioxidant Defence System of Soybean Plants. AoB Plants 2021, 13, plab026. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of Melatonin in Abiotic Stress Resistance in Plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin Enhances Plant Growth and Abiotic Stress Tolerance in Soybean Plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef]
- Sheikhalipour, M.; Mohammadi, S.A.; Esmaielpour, B.; Zareei, E.; Kulak, M.; Ali, S.; Nouraein, M.; Bahrami, M.K.; Gohari, G. Exogenous Melatonin Increases Salt Tolerance in Bitter Melon by Regulating Ionic Balance, Antioxidant System and Secondary Metabolism-Related Genes. BMC Plant Biol. 2022, 33, 380. [Google Scholar] [CrossRef]
- Jiang, D.; Lu, B.; Liu, L.; Duan, W.; Meng, Y.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Dong, H.; et al. Exogenous Melatonin Improves the Salt Tolerance of Cotton by Removing Active Oxygen and Protecting Photosynthetic Organs. BMC Plant Biol. 2021, 21, 331. [Google Scholar] [CrossRef]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of Melatonin in Alleviating Cold Stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef]
- Hernández-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin: A Growth-Stimulating Compound Present in Lupin Tissues. Planta 2004, 220, 140–144. [Google Scholar] [CrossRef]
- Sharp, R.G.; Else, M.A.; Cameron, R.W.; Davies, W.J. Water Deficits Promote Flowering in Rhododendron via Regulation of Pre and Post Initiation Development. Sci. Hortic. 2009, 120, 511–517. [Google Scholar] [CrossRef]
- Wada, K.C.; Takeno, K. Stress-Induced Flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef]
- Kolář, J.; Johnson, C.H.; Macháčková, I. Exogenously Applied Melatonin (N -Acetyl-5-Methoxytryptamine) Affects Flowering of the Short-Day Plant Chenopodium rubrum. Physiol. Plant 2003, 118, 605–612. [Google Scholar] [CrossRef]
- Shi, H.; Wei, Y.; Wang, Q.; Reiter, R.; He, C. Melatonin Mediates the Stabilization of DELLA Proteins to Repress the Floral Transition in Arabidopsis. J. Pineal Res. 2016, 60, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hu, Q.; Liu, Y.; Cheng, P.; Cheng, H.; Liu, W.; Xing, X.; Guan, Z.; Fang, W.; Chen, S.; et al. Strigolactone Represses the Synthesis of Melatonin, Thereby Inducing Floral Transition in Arabidopsis thaliana in an FLC-Dependent Manner. J. Pineal Res. 2019, 67, e12582. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Lyu, M.J.A.; Leng, B.Y.; Zhu, X.G.; Wang, B.S. The Transcriptome of NaCl-Treated Limonium Bicolor, Leaves Reveals the Genes Controlling Salt Secretion of Salt Gland. Plant. Mol. Biol. 2016, 91, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Regulation of Ion Homeostasis under Salt Stress. Curr. Opin. Plant Boil. 2003, 6, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Lyu, M.J.; Leng, B.Y.; Zheng, G.Y.; Feng, Z.T.; Li, P.H.; Zhu, X.G.; Wang, B.S. Comparative Transcriptome Analysis of Developmental Stages of the Limonium Bicolor Leaf Generates Insights into Salt Gland Differentiation. Plant Cell Environ. 2015, 38, 1637–1657. [Google Scholar] [CrossRef]
- Hand, M.J.; Taffouo, V.D.; Nouck, A.E.; Nyemene, K.P.J.; Tonfack, B.; Meguekam, T.L.; Youmbi, E. Effects of Salt Stress on Plant Growth, Nutrient Partitioning, Chlorophyll Content, Leaf Relative Water Content, Accumulation of Osmolytes and Antioxidant Compounds in Pepper (Capsicum annuum L.) Cultivars. Not. Bot. Hortic. Agrobot. Cluj-Napoca 2017, 45, 481–490. [Google Scholar] [CrossRef]
- Wei, L.; Zhao, H.; Wang, B.; Wu, X.; Lan, R.; Huang, X.; Chen, B.; Chen, G.; Jiang, C.; Wang, J.; et al. Exogenous Melatonin Improves the Growth of Rice Seedlings by Regulating Redox Balance and Ion Homeostasis under Salt Stress. J. Plant Growth Regul. 2022, 41, 2108–2121. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The Mitigation Effects of Exogenous Melatonin on Salinity-Induced Stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef]
- Farouk, S.; AL-Huqail, A.A. Sustainable Biochar and/or Melatonin Improve Salinity Tolerance in Borage Plants by Modulating Osmotic Adjustment, Antioxidants, and Ion Homeostasis. Plants 2022, 11, 765. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, J.; Wu, Z.; Loka, D.A.; Zhao, W.; Chen, B.; Wang, Y.; Meng, Y.; Zhou, Z.; Gao, L. Effects of Single and Combined Exogenous Application of Abscisic Acid and Melatonin on Cotton Carbohydrate Metabolism and Yield under Drought Stress. Ind. Crops Prod. 2022, 176, 114302. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous Melatonin Improves Seedling Health Index and Drought Tolerance in Tomato. Plant Growth Regul. 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Liang, C.; Zheng, G.; Li, W.; Wang, Y.; Hu, B.; Wang, H.; Wu, H.; Qian, Y.; Zhu, X.-G.; Tan, D.-X.; et al. Melatonin Delays Leaf Senescence and Enhances Salt Stress Tolerance in Rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef]
- Muller, O.; Cohu, C.M.; Stewart, J.J.; Protheroe, J.A.; Demmig-Adams, B.; Adams, W.W. Association between Photosynthesis and Contrasting Features of Minor Veins in Leaves of Summer Annuals Loading Phloem via Symplastic versus Apoplastic Routes. Physiol Plant. 2014, 152, 174–183. [Google Scholar] [CrossRef]
- Gohari, G.; Farhadi, H.; Panahirad, S.; Zareei, E.; Labib, P.; Jafari, H.; Mahdavinia, G.; Hassanpouraghdam, M.B.; Ioannou, A.; Kulak, M.; et al. Mitigation of Salinity Impact in Spearmint Plants through the Application of Engineered Chitosan-Melatonin Nanoparticles. Int. J. Biol. Macromol. 2023, 224, 893–907. [Google Scholar] [CrossRef]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Lin, L.; Deng, Q.; Luo, X. Exogenous Melatonin Promotes Biomass Accumulation and Photosynthesis of Kiwifruit Seedlings under Drought Stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Ashraf, M.; Ali, Q. Relative Membrane Permeability and Activities of Some Antioxidant Enzymes as the Key Determinants of Salt Tolerance in Canola (Brassica napus L.). Environ. Exp. Bot. 2008, 63, 266–273. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A Review on Potential Plant-Basedwater Stress Indicators for Vegetable Crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Khan, T.A.; Saleem, M.; Fariduddin, Q. Melatonin Influences Stomatal Behavior, Root Morphology, Cell Viability, Photosynthetic Responses, Fruit Yield, and Fruit Quality of Tomato Plants Exposed to Salt Stress. J. Plant Growth Regul. 2022, 1–25. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, L.; Wang, X.; Wang, Z.; Zhang, H.; Chen, J.; Liu, X.; Wang, Y.; Li, C. Beneficial Effects of Exogenous Melatonin on Overcoming Salt Stress in Sugar Beets (Beta vulgaris L.). Plants 2021, 10, 886. [Google Scholar] [CrossRef]
- Saneoka, H.; Moghaieb, R.E.; Premachandra, G.S.; Fujita, K. Nitrogen Nutrition and Water Stress Effects on Cell Membrane Stability and Leaf Water Relations in Agrostis palustris Huds. Environ. Exp. Bot. 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Catala, A. The Ability of Melatonin to Counteract Lipid Peroxidation in Biological Membranes. Curr. Mol. Med. 2007, 7, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Chen, Y.-E.; Zhao, Y.-Q.; Ding, C.-B.; Liao, J.-Q.; Hu, C.; Zhou, L.-J.; Zhang, Z.-W.; Yuan, S.; Yuan, M. Exogenous Melatonin Alleviates Oxidative Damages and Protects Photosystem II in Maize Seedlings under Drought Stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Plant Salt Tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.; Fuggi, A.; Carillo, P. Hordeum Vulgare and Hordeum Maritimum Respond to Extended Salinity Stress Displaying Different Temporal Accumulation Pattern of Metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef]
- Qiao, Y.; Ren, J.; Yin, L.; Liu, Y.; Deng, X.; Liu, P.; Wang, S. Exogenous Melatonin Alleviates PEG-Induced Short-Term Water Deficiency in Maize by Increasing Hydraulic Conductance. BMC Plant Biol. 2020, 20, 218. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Influence of Exogenous Salicylic Acid and Nitric Oxide on Growth, Photosynthesis, and Ascorbate-Glutathione Cycle in Salt Stressed Vigna angularis. Biomolecules 2020, 10, 42. [Google Scholar] [CrossRef]
- Liang, L.; Li, D.; Chen, Y.; Cheng, J.; Zhao, G.; Fahima, T.; Yan, J. Selenium Mitigates Salt-Induced Oxidative Stress in Durum Wheat (Triticum durum Desf.) Seedlings by Modulating Chlorophyll Fluorescence, Osmolyte Accumulation, and Antioxidant System. 3 Biotech 2020, 10, 368. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, J.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.B.M.; Chen, L.; Fu, J. Comparative Photosynthetic and Metabolic Analyses Reveal Mechanism of Improved Cold Stress Tolerance in Bermudagrass by Exogenous Melatonin. Plant Physiol. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef]
- Shi, H.; Chen, Y.; Tan, D.-X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin Induces Nitric Oxide and the Potential Mechanisms Relate to Innate Immunity against Bacterial Pathogen Infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef]
- Gao, W.; Feng, Z.; Bai, Q.; HeWang, J.Y. Melatonin-Mediated Regulation of Growth and Antioxidant Capacity in Salt-Tolerant Naked Oat under Salt Stress. Int. J. Mol. Sci. 2019, 20, 1176. [Google Scholar] [CrossRef]
- Zeng, L.; CaiJS, L.; Lu, G.; Li, C.; Fu, G.; Zhang, X.; Ma, H.; Liu, Q.; Zou, X.; Cheng, Y. Exogenous Application of a Low Concentration of Melatonin Enhances Salt Tolerance in Rapeseed (Brassica napus L.) Seedlings. J. Integr. Agric. 2018, 17, 328–335. [Google Scholar] [CrossRef]
- Mann, A.; Kaur, G.; Kumar, A.; Sanwal, S.K.; Singh, J.; Sharma, P.C. Physiological Response of Chickpea (Cicer arietinum L.) at Early Seedling Stage under Salt Stress Conditions. Legum. Res. 2019, 42, 625–632. [Google Scholar] [CrossRef]
- Roshdy, A.E.-D.; Alebidi, A.; Almutairi, K.; Al-Obeed, R.; Elsabagh, A. The Effect of Salicylic Acid on the Performances of Salt Stressed Strawberry Plants, Enzymes Activity, and Salt Tolerance Index. Agronomy 2021, 11, 775. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Shoot Length (Cm) | No of Leaves | Leaf Area (Cm2) | FW (g) | DW (g) |
|---|---|---|---|---|---|
| Control (non-stressed) | 18.88 ± 0.2 aA | 6.67 ± 0.15 aA | 47.96 ± 0.2 aA | 18.11 ± 0.2 aA | 3.06 ± 0.02 aA |
| S1 Treatments | |||||
| MT (0) | 12.88 ± 0.12 eB | 4.52 ± 0.12 eB | 19.98 ± 0.2 eB | 12.80 ± 0.1 eB | 1.77 ± 0.02 eB |
| MT (50) | 14.29 ± 0.22 dB | 5.07 ± 0.11 dB | 22.47 ± 0.2 dB | 13.90 ± 0.1 dB | 2.07 ± 0.01 dB |
| MT (100) | 15.62 ± 0.02 cB | 5.40 ± 0.18 cB | 27.99 ± 0.1 cB | 15.72 ± 0.2 cB | 2.25 ± 0.01 cB |
| MT (200) | 15.89 ± 1.3 bB | 5.80 ± 0.02 bB | 31.57 ± 0.2 bB | 16.63 ± 0.2 bB | 2.57 ± 0.02 bB |
| S2 Treatments | |||||
| MT (0) | 12.15 ± 0.2 eC | 4.40 ± 0.02 eC | 18.71 ± 0.2 eC | 11.48 ± 0.2 eC | 1.65 ± 0.02 eC |
| MT (50) | 14.20 ± 0.1 dC | 4.87 ± 0.01 dC | 20.60 ± 0.3 dC | 13.64 ± 0.3 dC | 1.80 ± 0.01 dC |
| MT (100) | 15.55 ± 0.2 cC | 5.27 ± 0.01 cC | 26.25 ± 0.12 cC | 14.67 ± 0.1 cC | 2.24 ± 0.02 cC |
| MT (200) | 15.80 ± 0.1 bC | 5.33 ± 0.11 bC | 29.71 ± 0.4 bC | 16.34 ± 0.2 bC | 2.34 ± 0.02 bC |
| Traits | STI % |
|---|---|
| Shoot length | 64.4 ± 0.2 |
| Number of leaves | 65.9 ± 0.2 |
| Total leaf area | 39.0 ± 0.03 |
| Shoot fresh weight (FW) | 63.4 ± 0.0 |
| Shoot dry weight (DW) | 53.9 ± 0.3 |
| Emergence of Flower buds | 74.3 ± 2.5 |
| Total chlorophyll content | 44.4 ± 1.2 |
| Total carotenoids content | 54.5 ± 2.1 |
| Leaf Na+ content | 383.1 ± 23.7 |
| Relative water content (RWC) | 71.7 ± 0.3 |
| Proline content | 202.5 ± 2.1 |
| Electrolyte leakage (El) | 309.9 ± 0.3 |
| Peroxidase (POD) | 321.3 ± 28.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eisa, E.A.; Honfi, P.; Tilly-Mándy, A.; Mirmazloum, I. Exogenous Melatonin Application Induced Morpho-Physiological and Biochemical Regulations Conferring Salt Tolerance in Ranunculus asiaticus L. Horticulturae 2023, 9, 228. https://doi.org/10.3390/horticulturae9020228
Eisa EA, Honfi P, Tilly-Mándy A, Mirmazloum I. Exogenous Melatonin Application Induced Morpho-Physiological and Biochemical Regulations Conferring Salt Tolerance in Ranunculus asiaticus L. Horticulturae. 2023; 9(2):228. https://doi.org/10.3390/horticulturae9020228
Chicago/Turabian StyleEisa, Eman Abdelhakim, Péter Honfi, Andrea Tilly-Mándy, and Iman Mirmazloum. 2023. "Exogenous Melatonin Application Induced Morpho-Physiological and Biochemical Regulations Conferring Salt Tolerance in Ranunculus asiaticus L." Horticulturae 9, no. 2: 228. https://doi.org/10.3390/horticulturae9020228
APA StyleEisa, E. A., Honfi, P., Tilly-Mándy, A., & Mirmazloum, I. (2023). Exogenous Melatonin Application Induced Morpho-Physiological and Biochemical Regulations Conferring Salt Tolerance in Ranunculus asiaticus L. Horticulturae, 9(2), 228. https://doi.org/10.3390/horticulturae9020228