
Mechanisms Underlying the Pathogenic and Endophytic Lifestyles in Diaporthe: An Omics-Based Approach



Abstract
:1. Introduction
2. Criteria Used for Selection of Studies and Search Strategy
3. Species of Diaporthe: Pathogens or Endophytes?

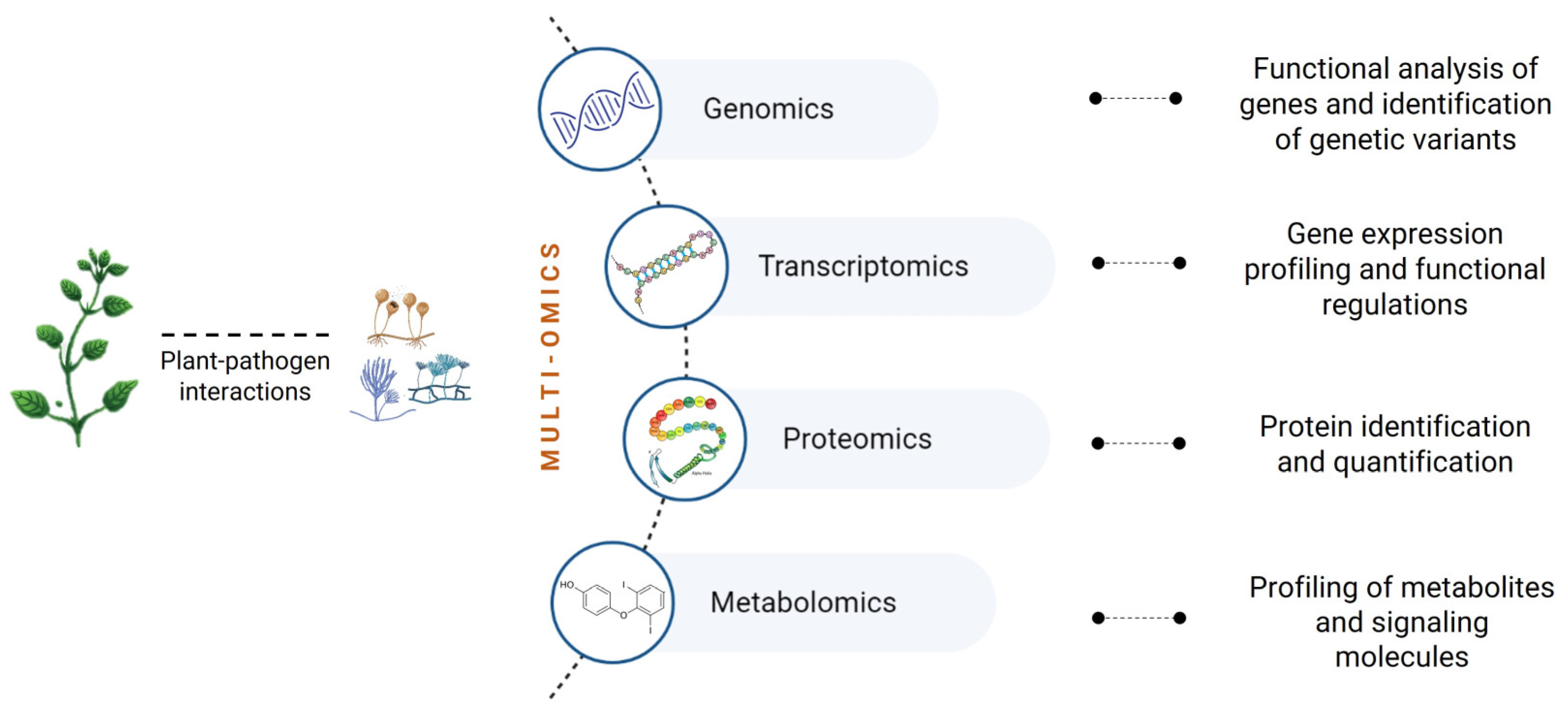{kind=link}
| Plant Hosts | Species | References |
|---|---|---|
| Agricultural Crops | ||
| Prunus dulcis | D. amygdali, D. novem, D. foeniculina | [39,45,46,47] |
| Malus domestica | D. malorum, D. leucospermi, D. eres, D. ambigua, D. foeniculina | [34,35,48] |
| Vaccinium corymbosum | D. foeniculina, D. rudis, D. leucospermi, D. eres, D. ambigua, D. crousii, D. amygdali, D. oxe, D. passiflorae, D. malorum, D. hybrida | [49,50,51,52,53,54,55,56] |
| Citrus spp. | D. foeniculina, D. citri, D. citrishinensis, D. limonicola, D. masirevicii, D. passifloricola | [11,21,57,58] |
| Vitis vinifera | D. amygdali, D. celeris, D. rudis, D. ampelina, D. eres, D. gulyae, D. hungariae, D. sojae, D. guangxiensis, D. novem | [4,9,39,44,59] |
| Corylus avellana | D. eres, D. amygdali, D. sojae, D. cercidis, D. hungariae | [32,60] |
| Prunus persica | D. amygdali, D. eres, D. caryae, D. cercidis, D. hongkongensis, D. unshiuensis | [15,45,61] |
| Pyrus communis | D. infecunda, D. eres, D. terebinthifolii, D. phaseolorum, D. oxe, D. sojae, D. amygdali, D. hongkongensis | [14,35] |
| Gycine max | D. caulivora, D. longicolla, D. sojae, D. novem | [16,43,62,63] |
| Helianthus annus | D. helianthi, D. gulyae, D. novem, D. caulivora, D. kochmanii, D. kongii, D. stewartii | [40,41,43,64] |
| Juglans regia | D. capcisi, D. eres, D. amygdali | [32,65,66] |
| Forest Trees | ||
| Acacia spp. | D. accacigena, D. acaciarum, D. heterophyllae, D. fraxini-angustifoliae | [67,68,69] |
| Acer spp. | D. acericola, D. leucospermi | [37,70] |
| Eucalyptus spp. | D. crousii, D. malorum, D. eucalyptorum | [71,72] |
| Fraxinus spp. | D. silvicola, D. fraxinicola | [42,73] |
| Pinus spp. | D. eres, D. rudis | [71] |
| Quercus spp. | D. eres, D. foeniculina, D. rudis | [60,73] |
| Ornamental Plants | ||
| Camellia sinensis | D. amygdali, D. eres, D. hongkongensis, D. tulliensis, D. passiflorae | [74,75,76] |
| Foeniculum vulgare | D. foeniculina, D. angelicae | [77] |
| Hydrangea macrophylla | D. foeniculina, D. leucospermi, D. novem | [43,70] |
| Lithocarpus glabra | D. amygdali, D. longicicolla, D. lithocarpus | [78] |
| Rosa spp. | D. rosae, D. rosiphthora, D. rudis, D. eres, D. foeniculina | [3,79] |
4. Omics to Study Fungal Plant Pathogens
- (1)
- Adaptation patterns of pathogens under changing environmental conditions.
- (2)
- Molecular traits underlying the infection processes.
- (3)
- Patterns of endophytic fungal community and their implications for disease development.

4.1. Genomics
- (1)
- (2)
- (3)
- Cellular transporters of ions (e.g., zinc, sulfur, copper), molecules that enhance pathogenicity (e.g., peroxiredoxin, tetraspanin), and sugars from plant polysaccharides degradation (e.g., xylose, inositol, and glycerol) [22].
- (4)
- (1)
- (2)
- (3)
- (4)
| Species | Effector Candidate | Effector Location | References |
|---|---|---|---|
| D. amygdali | glycosyl hydrolase family 61 | Apoplastic | [22] |
| aldehyde reductase 1 | Apoplastic | ||
| putative cfem domain-containing protein | Cytoplasmic | ||
| putative metalloprotease | Apoplastic | ||
| murein transglycosylase | Apoplastic | ||
| acetyl xylan esterase | Apoplastic | ||
| putative cerato-ulmin | Apoplastic | ||
| putative gas1-like protein | Apoplastic | ||
| putative secreted aspartic proteinase precursor | Apoplastic | ||
| Pectate lyase H | Apoplastic | ||
| glycosyl hydrolase family 61 | Apoplastic | ||
| D. capsici | sterigmatocystin biosynthesis peroxidase stcC | Apoplastic | [24] |
| pectate lyase F | Apoplastic | ||
| putative 1,4-beta-D-glucan cellobiohydrolase A | Apoplastic | ||
| putative proline-rich antigen | Apoplastic | ||
| chitin deacetylase | Apoplastic | ||
| xylanase G1 | Apoplastic | ||
| putative chitin binding protein | Apoplastic | ||
| putative mannose binding | Apoplastic | ||
| putative gas1-like protein | Apoplastic | ||
| glycoside hydrolase family 11 protein | Apoplastic | ||
| Cell wall glyco protein | Cytoplasmic | ||
| Poly(rC)-binding protein 4 | Cytoplasmic | ||
| D. caulivora | putative sterigmatocystin biosynthesis peroxidase stcC | Apoplastic | [24] |
| putative proline-rich antigen | Apoplastic | ||
| putative cytochrome p450 | Apoplastic | ||
| xylanase G1 | Apoplastic | ||
| glycoside hydrolase | Apoplastic | ||
| pectate lyase | Apoplastic | ||
| peptidase S41 family protein | Apoplastic | ||
| chitin deacetylase | Apoplastic | ||
| putative aldehyde dehydrogenase | Apoplastic | ||
| pectate lyase F | Apoplastic | ||
| putative 1,4-beta-D-glucan cellobiohydrolase A | Apoplastic | ||
| D. citri | chitin deacetylase | Apoplastic | [24] |
| Glucan endo-1,3-beta-glucosidase | Apoplastic | ||
| putative 1,4-beta-D-glucan cellobiohydrolase A | Apoplastic | ||
| putative sterigmatocystin biosynthesis peroxidase stcC | Apoplastic | ||
| cholera enterotoxin subunit A2 | Apoplastic | ||
| pectate lyase | Apoplastic | ||
| polysaccharide lyase family 3 protein | Apoplastic | ||
| Chitin binding protein | Apoplastic | ||
| Acetylxylan esterase-like protein | Apoplastic | ||
| pectate lyase F | Apoplastic | ||
| xylanase G1 | Apoplastic | ||
| putative riboflavin-aldehyde forming enzyme protein | Apoplastic | ||
| D. destruens | pectate lyase | Apoplastic | [24] |
| NPP1 domain-containing protein | Apoplastic | ||
| xylanase G1 | Apoplastic | ||
| cellulose binding CEL1 | Apoplastic | ||
| putative pectate lyase F | Apoplastic | ||
| Poly(rC)-binding protein 4 | Apoplastic | ||
| chitin deacetylase | Apoplastic | ||
| ribosomal protein s17 | Cytoplasmic | ||
| Protein CAP22 | Apoplastic | ||
| fungal cellulose binding domain-containing protein | Apoplastic | ||
| D. eres (syn. D. phragmitis) | pectate lyase | Apoplastic | [24] |
| Acetylxylan esterase 2 | Apoplastic | ||
| putative glutamine-serine-proline rich | Apoplastic | ||
| putative rhamnogalacturonan acetylesterase | Apoplastic | ||
| xylanase G1 | Apoplastic | ||
| Protein CAP22 | Apoplastic | ||
| lytic polysaccharide monooxygenase | Apoplastic | ||
| pectate lyase F | Apoplastic | ||
| putative proline-rich antigen | Apoplastic | ||
| chitin deacetylase | Apoplastic | ||
| D. eres (syn. D. vaccinii) | putative metalloprotease | Apoplastic | [22] |
| carbohydrate-binding module family 50 protein | Apoplastic | ||
| putative glycoside hydrolase family 61 protein | Apoplastic | ||
| acetylxylan esterase | Apoplastic | ||
| putative ricin b lectin | Apoplastic | ||
| putative pectate lyase b | Apoplastic | ||
| aldehyde reductase 1 | Apoplastic | ||
| putative npp1 domain | Cytoplasmic | ||
| putative pectinesterase | Cytoplasmic | ||
| putative pectate lyase | Apoplastic | ||
| disulfide-isomerase erp38 | Cytoplasmic | ||
| D. longicolla | polysaccharide lyase family 3 protein | Apoplastic | [24] |
| putative carbohydrate-binding module family 1 protein | Apoplastic | ||
| carbohydrate esterase family 5 protein | Apoplastic | ||
| starch binding domain-containing protein | Apoplastic | ||
| putative pectate lyase F | Apoplastic | ||
| Acetylxylan esterase 2 | Apoplastic | ||
| pectate lyase | Apoplastic | ||
| cell wall protein PhiA | Apoplastic | ||
| xylanase G1 | Apoplastic | ||
| cellulose binding CEL1 | Apoplastic | ||
| fungal cellulose binding domain-containing protein | Apoplastic | ||
| Protein CAP22 | Apoplastic |
4.2. Transcriptomics
4.3. Proteomics
4.4. Metabolomics
5. Omics to Underline Fungal Lifestyle Switches
6. Conclusions and Future Perspectives
- (1)
- Elucidate the broad spectrum of putative effectors underlying infection processes as well as to understand the ecological and evolutionary dynamics of plant pathogens.
- (2)
- Identify differentially expressed genes to understand regulatory networks involved in infection processes, which can provide helpful information for the development of disease control strategies.
- (3)
- Unravel novel proteins that might be potential candidates for the enhancement of tolerance to fungal diseases.
- (4)
- Identify secreted metabolites and metabolic pathways to identify candidate biomarkers in the early stage of disease development.
- (5)
- Provide evidence on the role of secreted effectors, pectin-degrading enzymes, secondary metabolism enzymes, MAPK signaling pathways and metabolites involved in pathogenicity as determinants of endophytism.
- (1)
- Are endophytic isolates location-specific and host-adapted?
- (2)
- Do isolates express multiple lifestyle?
- (3)
- Are endophytic and pathogenic fungi genetically differentiated?
- (4)
- What are the molecular differences between endophytes and pathogens?
- (5)
- Does an imbalance in the phyllosphere microbial community trigger life modes switching?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bhunjun, C.S.; Niskanen, T.; Suwannarach, N.; Wannathes, N.; Chen, Y.J.; McKenzie, E.H.; Maharachchikumbura, S.S.; Buyck, B.; Zhao, C.L.; Fan, Y.G.; et al. The numbers of fungi: Are the most speciose genera truly diverse? Fungal Divers. 2022, 114, 387–462. [Google Scholar] [CrossRef]
- Chepkirui, C.; Stadler, M. The genus Diaporthe: A rich source of diverse and bioactive metabolites. Mycol. Prog. 2017, 16, 477–494. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostert, L.; Crous, P.W.; Kang, J.C.; Phillips, A.J.L. Species of Phomopsis and a Libertella sp. occurring on grapevines with specific reference to South Africa: Morphological, cultural, molecular and pathological characterization. Mycologia 2001, 93, 146–167. [Google Scholar] [CrossRef]
- Luo, M.; Guo, W.; Zhao, M.; Manawasinghe, I.S.; Guarnaccia, V.; Liu, J.; Hyde, K.D.; Dong, Z.; You, C. Endophytic Diaporthe Associated with Morinda officinalis in China. J. Fungi 2022, 8, 806. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, F.; Duan, W.; Crous, P.W.; Cai, L. Diaporthe is paraphyletic. IMA Fungus 2017, 8, 163–187. [Google Scholar] [CrossRef]
- Chaisiri, C.; Liu, X.; Lin, Y.; Fu, Y.; Zhu, F.; Luo, C. Phylogenetic and haplotype network analyses of Diaporthe eres species in China based on sequences of multiple loci. Biology 2021, 10, 179. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Hilário, S.; Gonçalves, M.F.M.; Alves, A. Using genealogical concordance and coalescent-based species delimitation to assess species boundaries in the Diaporthe eres complex. J. Fungi 2021, 7, 507. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by Diaporthe spp. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hariharan, G.; Prasannath, K. Recent advances in molecular diagnostics of fungal plant pathogens: A mini review. Front. Cell. Infect. Microbiol. 2021, 10, 600234. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Wang, X.; Guo, Y.; Du, Y.; Yang, Z.; Huang, X.; Hong, N.; Xu, W.; Wang, G. Characterization of Diaporthe species associated with peach constriction canker, with two novel species from China. MycoKeys 2021, 80, 77. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. The Diaporthe sojae species complex: Phylogenetic reassessment of pathogens associated with soybean, cucurbits and other field crops. Fungal Biol. 2015, 119, 383–407. [Google Scholar] [CrossRef]
- Hilário, S.; Santos, L.; Alves, A. Diaporthe amygdali, a species complex or a complex species? Fungal Biol. 2021, 125, 505–518. [Google Scholar] [CrossRef]
- Nagel, J.H.; Wingfield, M.J.; Slippers, B. Increased abundance of secreted hydrolytic enzymes and secondary metabolite gene clusters define the genomes of latent plant pathogens in the Botryosphaeriaceae. BMC Genom. 2021, 22, 589. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Yan, J.; Guo, M.; Xu, L.; Hou, L.; Zou, Q. Comparative genome analysis of plant ascomycete fungal pathogens with different lifestyles reveals distinctive virulence strategies. BMC Genom. 2022, 23, 34. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, H.; Wang, C.; Xu, J.R. Comparative analysis of fungal genomes reveals different plant cell wall degrading capacity in fungi. BMC Genom. 2013, 14, 274. [Google Scholar] [CrossRef] [Green Version]
- Gai, Y.; Xiong, T.; Xiao, X.; Li, P.; Zeng, Y.; Li, L.; Riely, B.K.; Li, H. The Genome Sequence of the Citrus Melanose Pathogen Diaporthe citri and Two Citrus-Related Diaporthe Species. Phytopathology 2021, 111, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Hilário, S.; Gonçalves, M.F.M.; Fidalgo, C.; Tacão, M.; Alves, A. Genome Analyses of Two Blueberry Pathogens: Diaporthe amygdali CAA958 and Diaporthe eres CBS 160.32. J. Fungi 2022, 8, 804. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Darwish, O.; Alkharouf, N.W.; Musungu, B.; Matthews, B.F. Analysis of the genome sequence of Phomopsis longicolla: A fungal pathogen causing Phomopsis seed decay in soybean. BMC Genom. 2017, 18, 688. [Google Scholar] [CrossRef] [PubMed]
- Mena, E.; Garaycochea, S.; Stewart, S.; Montesano, M.; Ponce De León, I. Comparative genomics of plant pathogenic Diaporthe species and transcriptomics of Diaporthe caulivora during host infection reveal insights into pathogenic strategies of the genus. BMC Genom. 2022, 23, 175. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Jensen, B.; Jørgensen, H.J. Fungal endophytes in plants and their relationship to plant disease. Curr. Opin. Microbiol. 2022, 69, 102177. [Google Scholar] [CrossRef]
- Constantin, M.E.; Fokkens, L.; De Sain, M.; Takken, F.L.; Rep, M. Number of candidate effector genes in accessory genomes differentiates pathogenic from endophytic Fusarium oxysporum strains. Front. Plant Sci. 2021, 12, 761740. [Google Scholar] [CrossRef]
- Lowe, R.G.; Howlett, B.J. Indifferent, affectionate, or deceitful: Lifestyles and secretomes of fungi. PLoS Pathog. 2012, 8, e1002515. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Redkar, A.; Sabale, M.; Zuccaro, A.; Di Pietro, A. Determinants of endophytic and pathogenic lifestyle in root colonizing fungi. Curr. Opin. Plant Biol. 2022, 67, 102226. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmeier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A.; et al. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Yang, Q.; Bezerra, J.D.; Alvarez, L.V.; Tian, C. Diaporthe from walnut tree (Juglans regia) in China, with insight of the Diaporthe eres complex. Mycol. Prog. 2018, 17, 841–853. [Google Scholar] [CrossRef]
- Arciuolo, R.; Santos, C.; Soares, C.; Castello, G.; Spigolon, N.; Chiusa, G.; Lima, N.; Battilani, P. Molecular characterization of Diaporthe species associated with hazelnut defects. Front. Plant Sci. 2020, 11, 611655. [Google Scholar] [CrossRef]
- Thompson, S.M.; Grams, R.A.; Neate, S.M.; Shivas, R.G.; Ryley, M.J.; Tan, Y.P.; Aitken, E.A.B.; Wright, G.C.; O’Connor, D.J. First reports of Diaporthe kongii, D. masirevicii, and D. ueckerae associated with stem and peg dieback on peanut in Australia. Plant Dis. 2018, 102, 1459. [Google Scholar] [CrossRef]
- Santos, L.; Phillips, A.J.L.; Crous, P.W.; Alves, A. Diaporthe species on Rosaceae with descriptions of D. pyracanthae sp. Mycosphere 2017, 8, 485–511. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Lupo, S. Diversity of fungal latent pathogens and true endophytes associated with fruit trees in Uruguay. J. Phytopathol. 2018, 166, 633–647. [Google Scholar] [CrossRef]
- Murali, T.; Suryanarayanan, T.; Geeta, R. Endophytic Phomopsis species: Host range and implications for diversity estimates. Can. J. Microbiol. 2006, 52, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, A.J.; Phillips, A.J.L. Advances in understanding Diaporthe (Editorial). Mycosphere 2017, 8, 7019. [Google Scholar]
- Cinelli, T.; Mondello, V.; Marchi, G.; Burruano, S.; Alves, A.; Mugnai, L. First report of Diaporthe eres associated with cane blight of grapevine (Vitis vinifera) in Italy. Plant Dis. 2016, 100, 532. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Travadon, R.; Baumgartner, K. Diversity of Diaporthe species associated with wood cankers of fruit and nut crops in northern California of fruit and nut crops in northern California. Mycologia 2015, 107, 926–940. [Google Scholar] [CrossRef]
- Mathew, F.M.; Olson, T.R.; Science, P.; Dakota, S. Identification of Sunflower (Helianthus annuus) accessions resistant to Diaporthe helianthi and Diaporthe gulyae. Plant Health Prog. 2018, 19, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Thompson, S.M.; Tan, Y.P.; Young, A.J.; Neate, S.M.; Aitken, E.A.B.; Shivas, R.G. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Persoonia 2011, 27, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China, with twelve new species described. MycoKeys 2018, 39, 97–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.M.; Vrandecic, K.; Cosic, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Niekerk, J.M.; Groenewald, J.Z.; Farr, D.F.; Fourie, P.H.; Haleen, F.; Crous, P.W. Reassessment of Phomopsis on grapevine. Australas. Plant Pathol. 2005, 34, 27–39. [Google Scholar] [CrossRef]
- Diogo, E.L.; Santos, J.M.; Phillips, A.J.L. Phylogeny, morphology and pathogenicity of Diaporthe and Phomopsis species on almond in Portugal. Fungal Divers. 2010, 44, 107–115. [Google Scholar] [CrossRef]
- León, M.; Berbegal, M.; Rodríguez-Reina, J.M.; Elena, G.; Abad-Campos, P.; Ramón-Albalat, A.; Olmo, D.; Vicent, A.; Luque, J.; Miarnau, X.; et al. Identification and characterization of Diaporthe spp. associated with twig cankers and shoot blight of almonds in Spain. Agronomy 2020, 10, 1062. [Google Scholar] [CrossRef]
- Beluzán, F.; Miarnau, X.; Torguet, L.; Zazurca, L.; Abad-Campos, P.; Luque, J.; Armengol, J. Susceptibility of almond (Prunus dulcis) cultivars to twig canker and shoot blight caused by Diaporthe amygdali. Plant Dis. 2022, 106, 1890–1897. [Google Scholar] [CrossRef]
- Ali, S.; Renderos, W.; Bevis, E.; Hebb, J.; Abbasi, P.A. Diaporthe eres causes stem cankers and death of young apple rootstocks in Canada. Can. J. Plant Pathol. 2020, 42, 218–227. [Google Scholar] [CrossRef]
- Lombard, L.; Van Leeuwen, G.C.M.; Guarnaccia, V.; Polizii, G.; Van Rijswick, P.C.J.; Rosendahl, C.H.M.; Gabler, J.; Crous, P.W. Diaporthe species associated with Vaccinium, with specific reference to Europe. Phytopathol. Mediterr. 2014, 53, 287–299. [Google Scholar]
- Elfar, K.; Torres, R.; Díaz, G.A.; Latorre, B. Characterization of Diaporthe australafricana and Diaporthe spp. associated with stem canker of blueberry in Chile. Plant Dis. 2013, 97, 1042–1050. [Google Scholar] [CrossRef] [Green Version]
- Hilário, S.; Santos, L.; Alves, A. Diversity and Pathogenicity of Diaporthe Species Revealed from a Survey of Blueberry Orchards in Portugal. Agriculture 2021, 11, 1271. [Google Scholar] [CrossRef]
- Farr, D.F.; Castlebury, L.A.; Rossman, A.Y. Morphological and molecular characterization of Phomopsis vaccinii and additional isolates of Phomopsis from blueberry and cranberry in the eastern United States. Mycologia 2002, 94, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Cardinaals, J.; Wenneker, M.; Voogd, B.; Van Leeuwen, M. Pathogenicity of Diaporthe spp. on two blueberry cultivars (Vaccinium corymbosum). Bull. OEPP 2018, 48, 128–134. [Google Scholar] [CrossRef]
- Hilário, S.; Amaral, I.A.; Gonçalves, M.F.M.; Lopes, A.; Santos, L.; Alves, A. Diaporthe species associated with twig blight and dieback of Vaccinium corymbosum in Portugal, with description of four new species. Mycologia 2020, 112, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Nabetani, K.; Wood, B.; Sabaratnam, S. Role of pycnidia in twig and blossom blight and stem dieback of highbush blueberry caused by Phomopsis vaccinii in British Columbia. Can. J. Plant Pathol. 2017, 39, 405–421. [Google Scholar] [CrossRef]
- van Bruggen, A.H.C.; West, J.S.; van der Werf, W.; Potting, R.P.J.; Gardi, C.; Koufakis, I.; Zelenev, V.V.; Narouei-Khandan, H.; Shilder, A.; Harmon, P. Input data needed for a risk model for the entry, establishment and spread of a pathogen (Phomopsis vaccinii) of blueberries and cranberries in the EU. Ann. Appl. Biol. 2018, 172, 126–147. [Google Scholar] [CrossRef]
- Chaisiri, C.; Liu, X.Y.; Yin, W.X.; Luo, C.X.; Lin, Y. Morphology characterization, molecular phylogeny, and pathogenicity of Diaporthe passifloricola on Citrus reticulata cv. Nanfengmiju in Jiangxi Province, China. Plants 2021, 10, 218. [Google Scholar] [CrossRef]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 3621. [Google Scholar] [CrossRef] [PubMed]
- Manawasinghe, I.S.; Dissanayake, A.; Li, X.; Liu, M.; Wanasinghe, D.; Xu, J.; Zhao, W.; Zhang, W.; Zhou, Y.; Hyde, K.; et al. High genetic diversity and species complexity of Diaporthe associated with grapevine dieback in China. Front. Microbiol. 2019, 10, 1936. [Google Scholar] [CrossRef]
- Wiman, N.G.; Webber, J.B., III; Wiseman, M.; Merlet, L. Identity and pathogenicity of some fungi associated with hazelnut (Corylus avellana L.) trunk cankers in Oregon. PLoS ONE 2019, 14, e0223500. [Google Scholar] [CrossRef]
- Thomidis, T.; Michailides, T.J. Studies on Diaporthe eres as a new pathogen of peach trees in Greece. Plant Dis. 2009, 93, 1293–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mena, E.; Stewart, S.; Montesano, M.; Ponce de León, I. Soybean stem canker caused by Diaporthe caulivora; pathogen diversity, colonization process, and plant defense activation. Front. Plant Sci. 2020, 10, 1733. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Wang, T.C.; Seo, M.J. First report of soybean pod and stem blight caused by Diaporthe phaseolorum var. sojae in Taiwan. Plant Dis. 2009, 93, 202. [Google Scholar] [CrossRef]
- Olson, T.R.; Kontz, B.; Gulya, T.J.; Markell, S.G.; Mathew, F.M. First report of Diaporthe stewartii causing Phomopsis stem canker of sunflower (Helianthus annuus) in Minnesota. Plant Dis. 2017, 101, 382. [Google Scholar] [CrossRef]
- Fang, X.; Qin, K.; Li, S.; Han, S.; Zhu, T. Whole genome sequence of Diaporthe capsici, a new pathogen of walnut blight. Genomics 2020, 112, 3751–3761. [Google Scholar] [CrossRef]
- Meng, L.; Yu, C.; Wang, C.; Li, G. First report of Diaporthe amygdali causing walnut twig canker in Shandong province of China. Plant Dis. 2018, 102, 1859. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F.; et al. Genera of phytopathogenic fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Shivas, R.G.; Edwards, J.; Seifert, K.A.; Alfenas, A.C.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.S.J.; Hiscock, N.; et al. Fungal Planet description sheets: 69–91. Persoonia 2011, 26, 108–156. [Google Scholar] [CrossRef]
- Tan, Y.P.; Edwards, J.; Grice, K.R.E.; Shivas, R.G. Molecular phylogenetic analysis reveals six new species of Diaporthe from Australia. Fungal Divers. 2013, 61, 251–260. [Google Scholar] [CrossRef]
- Santos, J.M.; Correia, V.G.; Phillips, A.J.L. Primers for mating-type diagnosis in Diaporthe and Phomopsis: Their use in teleomorph induction in vitro and biological species definition. Fungal Biol. 2010, 114, 255–270. [Google Scholar] [CrossRef]
- Lopes, A.F.; Batista, E.; Hilário, S.; Santos, L.; Alves, A. Occurrence of Diaporthe species in Eucalyptus globulus, Pinus pinaster and Quercus suber in Portugal. For. Pathol. 2021, 51, e12674. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Decock, C.A.; Dreyer, L.L.; Granke, L.L.; Guest, D.I.; Hardy, G.; Hausbeck, M.K.; et al. Fungal Planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Piao, C.G.; Li, Y. Two new species of Diaporthe (Diaporthaceae, Diaporthales) associated with tree cankers in the Netherlands. MycoKeys 2021, 85, 31. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Wang, J.Y.; Withee, P.; Tanjira, M.; Lin, S.R.; Suwannarach, N.; Kumla, J.; Elgorban, A.M.; Cheewangkoon, R. Molecular phylogenetic diversity and biological characterization of Diaporthe species associated with leaf spots of Camellia sinensis in Taiwan. Plants 2021, 10, 1434. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, F.; Cai, L. Unravelling Diaporthe species associated with Camellia. Syst. Biodivers. 2016, 14, 102–117. [Google Scholar] [CrossRef]
- Li, Y.; Tan, P.; Zhao, D.G. Diaporthe nobilis, a new record on Camellia sinensis in Guizhou Province, China. Mycosphere 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Santos, J.M.; Phillips, A.J.L. Resolving the complex of Diaporthe (Phomopsis) species occurring on Foeniculum vulgare in Portugal. Fungal Divers. 2009, 34, 111–125. [Google Scholar]
- Gao, Y.; Su, Y.; Sun, W.; Cai, L. Diaporthe species occurring on Lithocarpus glabra in China, with descriptions of five new species. Fungal Biol. 2015, 119, 295–309. [Google Scholar] [CrossRef]
- Caio, P.; Bruno, F.; Carlos, A.P.; Robert, B. Diaporthe rosiphthora sp. nov.: Yet another rose dieback fungus. Crop Prot. 2021, 139, 105365. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Zang, W.; Yan, J.Y.; Li, X.H. Molecular phylogenetic analysis reveals seven new Diaporthe species from Italy. Mycosphere 2017, 8, 853–877. [Google Scholar] [CrossRef]
- Torres, C.; Camps, R.; Aguirre, R.; Besoain, X.A. First report of Diaporthe rudis in Chile causing stem-end rot on ‘Hass’ avocado fruit imported from California, USA. Plant Dis. 2016, 100, 1951. [Google Scholar] [CrossRef]
- Huang, F.; Udayanga, D.; Wang, X.; Hou, X.; Mei, X.; Fu, Y.; Hyde, K.D.; Li, H. Endophytic Diaporthe associated with Citrus: A phylogenetic reassessment with seven new species from China. Fungal Biol. 2015, 119, 331–347. [Google Scholar] [CrossRef]
- Sebastianes, F.L.; Lacava, P.T.; Fávaro, L.C.; Rodrigues, M.B.; Araújo, W.L.; Azevedo, J.L.; Pizzirani-Kleiner, A.A. Genetic transformation of Diaporthe phaseolorum, an endophytic fungus found in mangrove forests, mediated by Agrobacterium tumefaciens. Curr. Genet. 2012, 58, 21–33. [Google Scholar] [CrossRef]
- Kuo, H.C.; Hui, S.; Choi, J.; Asiegbu, F.O.; Valkonen, J.; Lee, Y.H. Secret lifestyles of Neurospora crassa. Sci. Rep. 2014, 4, 5135. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Bhattacharjee, A.; Sharma, S. An ecological insight into the multifaceted world of plant-endophyte association. Crit. Rev. Plant Sci. 2021, 40, 127–146. [Google Scholar] [CrossRef]
- Abramczyk, B.; Marzec-Grządziel, A.; Grządziel, J.; Król, E.; Gałązka, A.; Oleszek, W. Biocontrol Potential and Catabolic Profile of Endophytic Diaporthe eres Strain 1420S from Prunus domestica L. in Poland—A Preliminary Study. Agronomy 2022, 12, 165. [Google Scholar] [CrossRef]
- Ghanbary, E.; Fathizadeh, O.; Pazhouhan, I.; Zarafshar, M.; Tabari, M.; Jafarnia, S.; Parad, G.A.; Bader, M.K.F. Drought and pathogen effects on survival, leaf physiology, oxidative damage, and defense in two middle eastern oak species. Forests 2021, 12, 247. [Google Scholar] [CrossRef]
- Kogel, K.H.; Franken, P.; Hückelhoven, R. Endophyte or parasite—What decides? Curr. Opin. Plant Biol. 2006, 9, 358–363. [Google Scholar] [CrossRef]
- Hilário, S.; Pinto, G.; Monteiro, P.; Santos, L.; Alves, A. The impact of two Diaporthe spp. on Vaccinium corymbosum physiological fitness under different water availability scenarios. Eur. J. Plant Pathol. 2023, 1–17. [Google Scholar] [CrossRef]
- Álvarez-Loayza, P.; White, J.F., Jr.; Torres, M.S.; Balslev, H.; Kristiansen, T.; Svenning, J.C.; Gil, N. Light converts endosymbiotic fungus to pathogen, influencing seedling survival and niche-space filling of a common tropical tree, Iriartea deltoidea. PLoS ONE 2011, 6, e16386. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, M.T.; Arnold, A.E. Geographic locality and host identity shape fungal endophyte communities in cupressaceous trees. Mycol. Res. 2008, 112, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Materatski, P.; Varanda, C.; Carvalho, T.; Dias, A.B.; Campos, M.D.; Rei, F.; do Rosário Félix, M. Spatial and temporal variation of fungal endophytic richness and diversity associated to the phyllosphere of olive cultivars. Fungal Biol. 2019, 123, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Brader, G.; Corretto, E.; Sessitsch, A. Metagenomics of Plant Microbiomes. In Functional Metagenomics: Tools and Applications; Charles, T., Liles, M., Sessitsch, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 179–200. [Google Scholar]
- Akram, S.; Ahmed, A.; He, P.; He, P.; Liu, Y.; Wu, Y.; Munir, S.; He, Y. Uniting the Role of Endophytic Fungi against Plant Pathogens and Their Interaction. J. Fungi 2023, 9, 72. [Google Scholar] [CrossRef]
- Husseini, A.A.; Derakhshandeh, M.; Tatlisu, N.B. Comprehensive review of transcriptomics (RNAs) workflows from blood specimens. Sep. Purif. Rev. 2022, 51, 57–77. [Google Scholar] [CrossRef]
- Alexovič, M.; Lindner, J.R.; Bober, P.; Longuespée, R.; Sabo, J.; Davalieva, K. Human peripheral blood mononuclear cells: A review of recent proteomic applications. Proteomics 2022, 22, 2200026. [Google Scholar] [CrossRef]
- Crandall, S.G.; Gold, K.M.; Jiménez-Gasco, M.D.M.; Filgueiras, C.C.; Willett, D.S. A multi-omics approach to solving problems in plant disease ecology. PLoS ONE 2020, 15, e0237975. [Google Scholar] [CrossRef] [PubMed]
- Rani, M.; Mangat, H.K.; Pathak, R.K.; Yadav, I.S. Harnessing the potential of omics for prevention and management of the complex crop plant’s diseases. J. Proteins Proteom. 2021, 12, 227–245. [Google Scholar] [CrossRef]
- Xu, T.-C.; Lu, Y.-H.; Wang, J.-F.; Song, Z.-Q.; Hou, Y.-G.; Liu, S.-S.; Liu, C.-S.; Wu, S.-H. Bioactive secondary metabolites of the genus Diaporthe and anamorph Phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 2021, 9, 217. [Google Scholar] [CrossRef]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Luo, J.; Bhattacharya, D. Advances in fungal phylogenomics and their impact on fungal systematics. Adv. Genet. 2017, 100, 309–328. [Google Scholar] [PubMed]
- Aylward, J.; Steenkamp, E.T.; Dreyer, L.L.; Roets, F.; Wingfield, B.D.; Wingfield, M.J. A plant pathology perspective of fungal genome sequencing. IMA Fungus 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yao, X.; Xhang, X.; Zou, H.; Chen, J.; Fang, B.; Huang, L. Draft genome sequence of Diaporthe batatatis causing dry rot disease in sweet potato. Plant Dis. 2022, 106, 737–740. [Google Scholar] [CrossRef]
- Hill, R.; Leitch, I.J.; Gaya, E. Targeting Ascomycota genomes: What and how big? Fungal Biol. Rev. 2021, 36, 52–59. [Google Scholar] [CrossRef]
- Morales-Cruz, A.; Amrine, K.C.; Blanco-Ulate, B.; Lawrence, D.P.; Travadon, R.; Rolshausen, P.E.; Baumgartner, K.; Cantu, D. Distinctive expansion of gene families associated with plant cell wall degradation, secondary metabolism, and nutrient uptake in the genomes of grapevine trunk pathogens. BMC Genom. 2015, 16, 469. [Google Scholar] [CrossRef] [Green Version]
- Savitha, J.; Bhargavi, S.D.; Praveen, V.K. Complete genome sequence of the endophytic fungus Diaporthe (Phomopsis) ampelina. Genome Announc. 2016, 4, e00477. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Song, Q.; Martins, A.M.; Cregan, P. Draft genome sequence of Diaporthe aspalathi isolate MS-SSC91, a fungus causing stem canker in soybean. Genom. Data 2016, 7, 262–263. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhang, X.; Yang, Y.; Zou, H.; Fang, B.; Liu, W. High-Quality genome resource of Diaporthe destruens causing foot rot disease of sweet potato. Plant Dis. 2021, 105, 3279–3281. [Google Scholar] [CrossRef]
- Liu, X.Y.; Chaisiri, C.; Lin, Y.; Yin, W.X.; Luo, C.X. Whole-Genome sequence of Diaporthe citri Isolate NFHF-8-4, the causal Agent of Citrus melanose. Mol. Plant-Microbe Interact. 2021, 34, 845–847. [Google Scholar] [CrossRef]
- Wang, X.; Dong, H.; Lan, J.; Liu, Y.; Liang, K.; Lu, Q.; Fang, Z.; Liu, P. High-quality genome resource of the pathogen of Diaporthe (Phomopsis) phragmitis causing kiwifruit soft rot. Mol. Plant-Microbe Interact. 2021, 34, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, R.; Scala, F.; Vergara, M.; Thon, M.R.; Ruocco, M. Draft whole-genome sequence of the Diaporthe helianthi 7/96 strain, causal agent of sunflower stem canker. Genom. Data 2016, 10, 151–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emanuel, I.B.; Konkel, Z.M.; Scott, K.L.; Valero David, G.E.; Slot, J.C.; Peduto Hand, F. Whole-Genome Sequence Data for the Holotype Strain of Diaporthe ilicicola, a Fungus Associated with Latent Fruit Rot in Deciduous Holly. Microbiol. Resour. Announc. 2022, 11, e00631-22. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Song, Q.; Ji, P.; Cregan, P. Draft genome sequence of Phomopsis longicolla type strain TWH P74, a fungus causing Phomopsis seed decay in soybean. Genome Announc. 2015, 3, e00010-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, Z.; You, Q.; Li, Z.; Sun, B.; Xu, X.; Li, Y.; Li, Z.; Wang, H.; Gong, C.; Xu, X.; et al. The first genome sequence of Phomopsis vexans: A fungal pathogen causing Phomopsis blight in eggplant. Biologia 2023, 78, 543–548. [Google Scholar] [CrossRef]
- Tulsook, K.; Isarangkul, D.; Sriubolmas, N.; Kittakoop, P.; Wiyakrutta, S. Draft genome sequence of Diaporthe sp. strain HANT25, an endophytic fungus producing mycoepoxydiene. Microbiol. Resour. Announc. 2020, 9, e00805-20. [Google Scholar] [CrossRef]
- Vieira, A.; Silva, D.N.; Várzea, V.; Paulo, O.S.; Batista, D. Genome-wide signatures of selection in Colletotrichum kahawae reveal candidate genes potentially involved in pathogenicity and aggressiveness. Front. Microbiol. 2019, 10, 1374. [Google Scholar] [CrossRef]
- Hartmann, F.E. Using structural variants to understand the ecological and evolutionary dynamics of fungal plant pathogens. New Phytol. 2022, 234, 43–49. [Google Scholar] [CrossRef]
- Boufleur, T.R.; Massola Júnior, N.S.; Tikami, Í.; Sukno, S.A.; Thon, M.R.; Baroncelli, R. Identification and comparison of Colletotrichum secreted effector candidates reveal two independent lineages pathogenic to soybean. Pathogens 2021, 10, 1520. [Google Scholar] [CrossRef]
- Selin, C.; De Kievit, T.R.; Belmonte, M.F.; Fernando, W.D. Elucidating the role of effectors in plant-fungal interactions: Progress and challenges. Front. Microbiol. 2016, 7, 600. [Google Scholar] [CrossRef] [Green Version]
- Ball, B.; Langille, M.; Geddes-McAlister, J. Fun(gi)omics: Advanced and diverse technologies to explore emerging fungal pathogens and define mechanisms of antifungal resistance. MBio 2020, 11, e01020-20. [Google Scholar] [CrossRef]
- Félix, C.; Meneses, R.; Gonçalves, M.F.M.; Tilleman, L.; Duarte, A.S.; Jorrín-Novo, J.V.; Van de Peer, Y.; Deforce, D.; Van Nieuwerburgh, F.; Esteves, A.C.; et al. A multi-omics analysis of the grapevine pathogen Lasiodiplodia theobromae reveals that temperature affects the expression of virulence-and pathogenicity-related genes. Sci. Rep. 2019, 9, 13144. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, M.F.M.; Nunes, R.B.; Tilleman, L.; Van de Peer, Y.; Deforce, D.; Van Nieuwerburgh, F.; Esteves, A.C.; Alves, A. Dual RNA sequencing of Vitis vinifera during Lasiodiplodia theobromae infection unveils host-pathogen interactions. Int. J. Mol. Sci. 2019, 20, 6083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, S.; Visser, E.A.; Zwart, L.; Du Toit, Y.; Bhadauria, V.; Shuey, L.S. Dual RNA-seq to elucidate the plant-pathogen duel. Curr. Issues Mol. Biol. 2017, 27, 127–142. [Google Scholar] [PubMed]
- Zhang, W.; Yan, J.; Li, X.; Xing, Q.; Chethana, K.T.; Zhao, W. Transcriptional response of grapevine to infection with the fungal pathogen Lasiodiplodia theobromae. Sci. Rep. 2019, 9, 5387. [Google Scholar] [CrossRef] [Green Version]
- Westermann, A.J.; Gorski, S.A.; Vogel, J. Dual RNA-seq of pathogen and host. Nat. Rev. Microbiol. 2012, 10, 618. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Wang, Y.; Li, Z.; Zhang, H.; Ye, J.; Li, G. Gene expression changes during the gummosis development of peach shoots in response to Lasiodiplodia theobromae infection using RNA-Seq. Front. Physiol. 2016, 7, 170. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Hu, R.; Fan, Z.; Chen, Q.; Jiang, Y.; Huang, W.; Tao, X. Dual RNA-Seq reveals the genome-wide expression profiles during the compatible and incompatible interactions between Solanum tuberosum and Phytophthora infestans. Front. Plant Sci. 2022, 13, 817199. [Google Scholar] [CrossRef]
- Guan, Y.; Chen, M.; Ma, Y.; Du, Z.; Yuan, N.; Li, Y.; Xiao, J.; Zhang, Y. Whole-genome and time-course dual RNA-Seq analyses reveal chronic pathogenicity-related gene dynamics in the ginseng rusty root rot pathogen Ilyonectria robusta. Sci. Rep. 2020, 10, 1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, E.; Shuey, L.S.; Naidoo, S.; Mamni, T.; Berger, D.K.; Myburg, A.A.; van den Berg, N.; Naidoo, S. Dual RNA-sequencing of Eucalyptus nitens during Phytophthora cinnamomi challenge reveals pathogen and host factors influencing compatibility. Front. Plant Sci. 2016, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora-Ballesteros, C.; Pinto, G.; Amaral, J.; Valledor, L.; Alves, A.; Diez, J.J.; Martín-García, J. Dual RNA-sequencing analysis of resistant (Pinus pinea) and susceptible (Pinus radiata) hosts during Fusarium circinatum challenge. Int. J. Mol. Sci. 2021, 22, 5231. [Google Scholar] [CrossRef]
- Książkiewicz, M.; Rychel-Bielska, S.; Plewiński, P.; Nuc, M.; Irzykowski, W.; Jędryczka, M.; Krajewski, P. The resistance of narrow-leafed lupin to Diaporthe toxica is based on the rapid activation of defense response genes. Int. J. Mol. Sci. 2021, 22, 574. [Google Scholar] [CrossRef]
- Elverson, T.R.; Kontz, B.J.; Markell, S.G.; Harveson, R.M.; Mathew, F.M. Quantitative PCR assays developed for Diaporthe helianthi and Diaporthe gulyae for phomopsis stem canker diagnosis and germplasm screening in sunflower (Helianthus annuus). Plant Dis. 2020, 104, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, B.; Voegele, R.T.; Link, T.I. Establishment of a quadruplex real-time PCR assay to distinguish the fungal pathogens Diaporthe longicolla, D. caulivora, D. eres, and D. novem on soybean. PLoS ONE 2020, 16, e0257225. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, K.; Kobayashi, Y.O.; Usui, M.; Nishioka, K.; Nakamura, M.; Kawano, S.; Okada, Y.; Kobayashi, A.; Miyasaka, A.; Hirayae, K.; et al. Real-Time PCR assay for the diagnosis and quantification of co-infections by Diaporthe batatas and Diaporthe destruens in sweet potato. Front. Plant Sci. 2020, 12, 694053. [Google Scholar] [CrossRef]
- Qi, H.; Jiang, Z.; Zhang, K.; Yang, S.; He, F.; Zhang, Z. PlaD: A transcriptomics database for plant defense responses to pathogens, providing new insights into plant immune system. Genom. Proteom. Bioinform. 2018, 16, 283–293. [Google Scholar] [CrossRef]
- Yang, H.; Luo, P. Changes in photosynthesis could provide important insight into the interaction between wheat and fungal pathogens. Int. J. Mol. Sci. 2021, 22, 8865. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.T.; Kim, S.G.; Hwang, D.H.; Kang, S.Y.; Kim, H.J.; Lee, B.H.; Lee, J.J.; Kang, K.Y. Proteomic analysis of pathogen-responsive proteins from rice leaves induced by rice blast fungus, Magnaporthe grisea. Proteomics 2004, 4, 3569–3578. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Chen, W.; Xin, Y.; Zhang, H.; Yan, C.; Yu, H.; Liu, H.; Xiao, W.; Wang, S.; Zheng, G.; et al. Proteomic analysis of strawberry leaves infected with Colletotrichum fragariae. J. Proteom. 2012, 75, 4074–4090. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manghwar, H.; Hussain, A.; Ali, Q.; Saleem, M.H.; Abualreesh, M.H.; Alatawi, A.; Ali, S.; Munis, M.F.H. Disease severity, resistance analysis, and expression profiling of pathogenesis-related protein genes after the inoculation of Fusarium equiseti in wheat. Agronomy 2021, 11, 2124. [Google Scholar] [CrossRef]
- Li, Y.; Feng, Y.; Lü, Q.; Yan, D.; Liu, Z.; Zhang, X. Comparative proteomic analysis of plant-pathogen interactions in resistant and susceptible poplar ecotypes infected with Botryosphaeria dothidea. Phytopathology 2019, 109, 2009–2021. [Google Scholar] [CrossRef]
- Yang, F.E.N.; Jensen, J.D.; Svensson, B.; Jørgensen, H.J.; Collinge, D.B.; Finnie, C. Secretomics identifies Fusarium graminearum proteins involved in the interaction with barley and wheat. Mol. Plant Pathol. 2012, 13, 445–453. [Google Scholar] [CrossRef]
- Huser, A.; Takahara, H.; Schmalenbach, W.; O’Connell, R. Discovery of pathogenicity genes in the crucifer anthracnose fungus Colletotrichum higginsianum, using random insertional mutagenesis. Mol. Plant-Microbe Interact. 2009, 22, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Reuveni, M.; Sheglov, N.; Eshel, D.; Prusky, D.; Ben-Arie, R. Virulence and the production of endo-1, 4-β-glucanase by Isolates of Alternaria alternata involved in the moldy-core disease of apples. J. Phytopathol. 2007, 155, 50–55. [Google Scholar] [CrossRef]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Silva, M.D.C.; Guerra-Guimarães, L.; Diniz, I.; Loureiro, A.; Azinheira, H.; Pereira, A.P.; Tavares, S.; Batista, D.; Várzea, V. An overview of the mechanisms involved in Coffee-Hemileia vastatrix interactions: Plant and pathogen perspectives. Agronomy 2022, 12, 326. [Google Scholar] [CrossRef]
- Balotf, S.; Wilson, R.; Tegg, R.S.; Nichols, D.S.; Wilson, C.R. Shotgun proteomics as a powerful tool for the study of the proteomes of plants, their pathogens, and plant-pathogen interactions. Proteomes 2022, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Spina, R.; Saliba, S.; Dupire, F.; Ptak, A.; Hehn, A.; Piutti, S.; Poinsignon, S.; Leclerc, S.; Bouguet-Bonnet, S.; Laurain-Mattar, D. Molecular identification of endophytic bacteria in Leucojum aestivum in vitro culture, NMR-based metabolomics study and LC-MS analysis leading to potential Amaryllidaceae alkaloid production. Int. J. Mol. Sci. 2021, 22, 1773. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Fleming’s penicillin producing strain is not Penicillium chrysogenum but P. rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Quinn, R. Rethinking antibiotic research and development: World War II and the penicillin collaborative. Am. J. Public Health 2013, 103, 426–434. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Marine Fungi: Opportunities and Challenges. Encyclopedia 2022, 2, 559–577. [Google Scholar] [CrossRef]
- Chen, H.; Singh, H.; Bhardwaj, N.; Bhardwaj, S.K.; Khatri, M.; Kim, K.H.; Peng, W. An exploration on the toxicity mechanisms of phytotoxins and their potential utilities. Crit. Rev. Environ. Sci. Technol. 2022, 52, 395–435. [Google Scholar] [CrossRef]
- Feussner, I.; Polle, A. What the transcriptome does not tell—Proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 2015, 26, 26–31. [Google Scholar] [CrossRef]
- Tan, K.C.; Ipcho, S.V.; Trengove, R.D.; Oliver, R.P.; Solomon, P.S. Assessing the impact of transcriptomics, proteomics and metabolomics on fungal phytopathology. Mol. Plant Pathol. 2010, 10, 703–771. [Google Scholar] [CrossRef]
- Mashabela, M.D.; Masamba, P.; Kappo, A.P. Metabolomics and Chemoinformatics in Agricultural Biotechnology Research: Complementary Probes in Unravelling New Metabolites for Crop Improvement. Biology 2022, 11, 1156. [Google Scholar] [CrossRef]
- Dickinson, E.; Rusilowicz, M.J.; Dickinson, M.; Charlton, A.J.; Bechtold, U.; Mullineaux, P.M.; Wilson, J. Integrating transcriptomic techniques and k-means clustering in metabolomics to identify markers of abiotic and biotic stress in Medicago truncatula. Metabolomics 2018, 14, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, O.A.H.; Griffin, J.L.; Jung, Y.H.; Shibato, J.; Rakwal, R.; Agrawal, G.K.; Jwa, N.S. Using metabolic profiling to assess plant-pathogen interactions: An example using rice (Oryza sativa) and the blast pathogen Magnaporthe grisea. Eur. J. Plant Pathol. 2011, 129, 539–554. [Google Scholar] [CrossRef]
- Hu, Z.; Chang, X.; Dai, T.; Li, L.; Liu, P.; Wang, G.; Liu, P.; Huang, Z.; Liu, X. Metabolic profiling to identify the latent infection of strawberry by Botrytis cinerea. Evol. Bioinform. 2019, 15, 1176934319838518. [Google Scholar] [CrossRef] [Green Version]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Metabolomic profiling of the host response of tomato (Solanum lycopersicum) following infection by Ralstonia solanacearum. Int. J. Mol. Sci. 2019, 20, 3945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kage, U.; Karre, S.; Kushalappa, A.C.; McCartney, C. Identification and characterization of a fusarium head blight resistance gene TaACT in wheat QTL-2DL. Plant Biotechnol. J. 2017, 15, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, C.B.D.; Santana, M.F. Prediction of the secretomes of endophytic and nonendophytic fungi reveals similarities in host plant infection and colonization strategies. Mycologia 2020, 112, 491–503. [Google Scholar] [CrossRef]
- Garcia, J.F.; Lawrence, D.P.; Morales-Cruz, A.; Travadon, R.; Minio, A.; Hernandez-Martinez, R.; Rolshausen, P.E.; Baumgartner, K.; Cantu, D. Phylogenomics of plant associated Botryosphaeriaceae species. Front. Microbiol. 2021, 12, 587. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Dung, J.K.S.; Johnson, D.A. From pathogen to endophyte: An endophytic population of Verticillium dahliae evolved from a sympatric pathogenic population. New Phytol. 2019, 222, 497–510. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.; Huang, P.W.; Dai, C.C. Endophytism or saprophytism: Decoding the lifestyle transition of the generalist fungus Phomopsis liquidambari. Microbiol. Res. 2018, 206, 99–112. [Google Scholar] [CrossRef]
- Becker, Y.; Eaton, C.J.; Brasell, E.; May, K.J.; Becker, M.; Hassing, B.; Cartwright, G.M.; Reinhold, L.; Scott, B. The fungal cell-wall integrity MAPK cascade is crucial for hyphal network formation and maintenance of restrictive growth of Epichloë festucae in symbiosis with Lolium perenne. Mol. Plant Microbe Interact. 2015, 28, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Segmüller, N.; Kokkelink, L.; Giesbert, S.; Odinius, D.; van Kan, J.; Tudzynski, P. NADPH oxidases are involved in differentiation and pathogenicity in Botrytis cinerea. Mol. Plant Microbe Interact. 2008, 21, 808–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, C.J.; Cox, M.P.; Scott, B. What triggers grass endophytes to switch from mutualism to pathogenism? Plant Sci. 2011, 180, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.M.; Wei, T.; Lou, H.; Shu, X.; Chen, Q. A critical review on communication mechanism within plant-endophytic fungi interactions to cope with biotic and abiotic stresses. J. Fungi 2021, 7, 719. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.J.; Wang, Z.Y.; Jones, M.A.; Smirnoff, N.; Talbot, N.J. Generation of reactive oxygen species by fungal NADPH oxidases is required for rice blast disease. Proc. Natl. Acad. Sci. USA 2007, 104, 11772–11777. [Google Scholar] [CrossRef] [Green Version]
- Lahrmann, U.; Strehmel, N.; Langen, G.; Frerigmann, H.; Leson, L.; Ding, Y.; Scheel, D.; Herklotz, S.; Hilbert, M.; Zuccaro, A. Mutualistic root endophytism is not associated with the reduction of saprotrophic traits and requires a noncompromised plant innate immunity. New Phytol. 2015, 207, 841–857. [Google Scholar] [CrossRef]
| Species | Strain | Host | JGI Project | GenBank Accession Number | Sequencing Platform | References |
|---|---|---|---|---|---|---|
| Diaporthe ampelina | DA912 | Vitis vinifera | NA | LCUC00000000 | Illumina HiSeq | [107] |
| S3MP | Commiphora wightii | - | LWAD00000000 | Illumina HiSeq | [108] | |
| Diaporthe amygdali | CAA958 | Vaccinium corymbosum | - | JAJATV000000000 | Illumina HiSeq | [22] |
| DUCC20226 | Malus sp. | - | JAJJOG000000000 | PacBio Sequel and Illumina | - | |
| Diaporthe aspalathi | MS-SSC91 | Glycine max | - | LJJS00000000 | Illumina HiSeq | [109] |
| Diaporthe batatas | CRI 302-4 | Ipomoea batatas | - | JAHWGW000000000 | Oxford Nanopore and PromethION | [105] |
| Diaporthe capsici | GY-Z16 | Juglans regia | - | WNXA00000000 | PacBio RSII | [65] |
| Diaporthe caulivora | D57 | G. max | - | JAMPTR000000000 | PacBio Sequel | [24] |
| Diaporthe cf. heveae | LGMF1633 | - | 1251927 | - | - | - |
| Diaporthe destruens | CRI305-2 | Ipomoea batatas | - | JACAAM010000000 | Oxford Nanopore and PromethION | [110] |
| Diaporthe citrisiana | ZJUD30 | Citrus unshiu | - | JADAZS000000000 | Illumina HiSeq | [21] |
| - | JADWDH000000000 | |||||
| Diaporthe citrichinensis | ZJUD34 | C. unshiu | - | JADAZR000000000 | Illumina HiSeq | [21] |
| Diaporthe citri | NFHF-8-4 | Citrus sp. | - | JACTAD000000000 | PacBio Sequel | [111] |
| ZJUD2 | C. reticulata | - | JADAZQ000000000 | Illumina HiSeq | [21] | |
| ZJUD14 | C. reticulata | - | JADAZP000000000 | Illumina HiSeq | ||
| Q7 | C. reticulata | - | JADAZO000000000 | Illumina HiSeq | ||
| Diaporthe eres (syn. D. phragmitis) | NJD1 | Actinidia deliciosa | - | JACDXY000000000 | PacBio RS | [112] |
| Diaporthe eres (syn. D. vaccinii) | CBS 160.32 | V. corymbosum | - | JAJATR000000000 | Illumina HiSeq | [22] |
| Diaporthe eres | Phoaprs 18-02 | Malus sp. | - | JAKJXL000000000 | Illumina NovaSeq | [48] |
| Phoaprs 18-03 | Malus sp. | - | JAKJXM000000000 | PacBio Sequel | ||
| Diaporthe helianthi | 7/96 | Helianthus annuus | NA | MAVT02000001 | Illumina MiSeq | [113] |
| Diaporthe ilicicola | FPH2015-502 | Ilex verticillata | - | JALPVH000000000 | Illumina and Oxford Nanopore | [114] |
| Diaporthe inconspicua | LGMF1612 | - | 1251935 | - | - | - |
| Diaporthe longicolla | MSPL 10–6 | G. max | - | AYRD00000000 | Illumina HiSeq | [23] |
| TWH P74 | - | JUJX00000000 | Illumina HiSeq | [115] | ||
| Diaporthe vexans | PV 4 | Solanum melongena | JAJLLZ000000000 | Oxford Nanopore | [116] | |
| Diaporthe vochysiae | LGMF1583 | Vochysia divergens | 1251933 | - | Pacbio | - |
| Diaporthe sp. | DP-2020a | Sequoia sempervirens | - | JACVEP000000000 | Illumina HiSeq | - |
| Diaporthe sp. | HANT25 | Hydnocarpus anthelminthicus | - | JACBFG000000000 | Illumina HiSeq | [117] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hilário, S.; Gonçalves, M.F.M. Mechanisms Underlying the Pathogenic and Endophytic Lifestyles in Diaporthe: An Omics-Based Approach. Horticulturae 2023, 9, 423. https://doi.org/10.3390/horticulturae9040423
Hilário S, Gonçalves MFM. Mechanisms Underlying the Pathogenic and Endophytic Lifestyles in Diaporthe: An Omics-Based Approach. Horticulturae. 2023; 9(4):423. https://doi.org/10.3390/horticulturae9040423
Chicago/Turabian StyleHilário, Sandra, and Micael F. M. Gonçalves. 2023. "Mechanisms Underlying the Pathogenic and Endophytic Lifestyles in Diaporthe: An Omics-Based Approach" Horticulturae 9, no. 4: 423. https://doi.org/10.3390/horticulturae9040423
APA StyleHilário, S., & Gonçalves, M. F. M. (2023). Mechanisms Underlying the Pathogenic and Endophytic Lifestyles in Diaporthe: An Omics-Based Approach. Horticulturae, 9(4), 423. https://doi.org/10.3390/horticulturae9040423