

First Report of the Branched Broomrape (Phelipanche schultzii (Mutel) Pomel.) on Fennel (Foeniculum vulgare Mill.) in Morocco

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
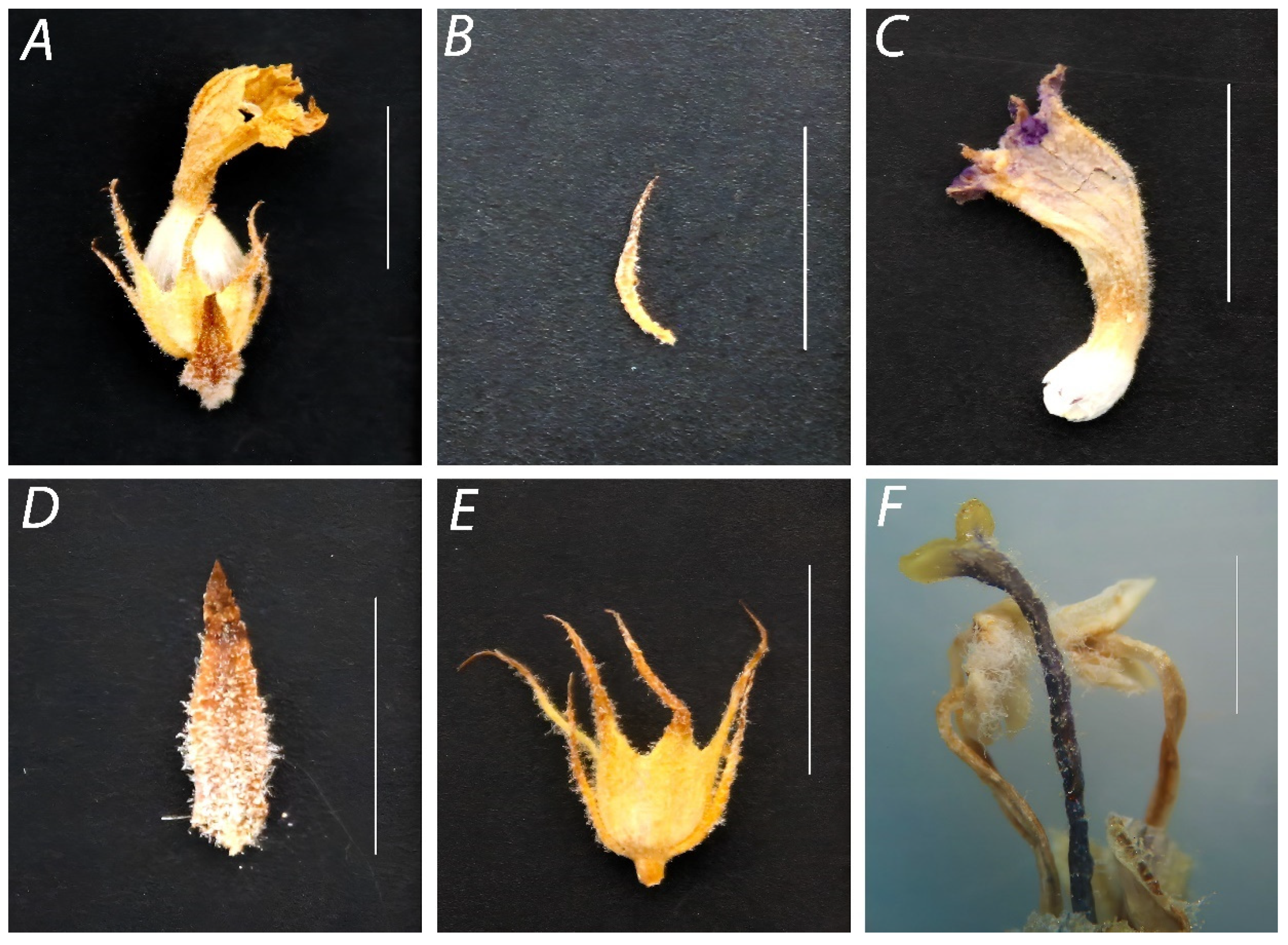
2.2. Morphological Characterization
2.3. DNA Extraction, PCR and Sequencing
2.4. Phylogenetic Analysis
3. Results
3.1. Description and Morphological Identification of Broomrape Species
3.2. Molecular Analysis
3.2.1. DNA Amplification
3.2.2. Sequence Analysis of the ITS-p5 and ITS-u4 rDNA Regions
3.2.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The haustorium, a specialized invasive organ in parasitic plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Westwood, J.H.; Depamphilis, C.W.; Das, M.; Fernández-Aparicio, M.; Honaas, L.A.; Timko, M.P.; Wafula, E.K.; Wickett, N.J.; Yoder, J.I. The parasitic plant genome project: New tools for understanding the biology of Orobanche and Striga. Weed Sci. 2012, 60, 295–306. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Reboud, X.; Gibot-Leclerc, S. Broomrape weeds. Underground mechanisms of parasitism and associated strategies for their control: A review. Front. Plant Sci. 2016, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Delavault, P. Knowing the parasite: Biology and genetics of Orobanche. Helia 2015, 38, 15–29. [Google Scholar] [CrossRef]
- Joel, D.; Hershenhorn, J.; Eizenberg, H.; Aly, R.; Ejeta, G.; Rich, P.; Ransom, J.; Sauerborn, J.; Rubiales, D. Biology and management of weedy root parasites. Hortic. Rev. 2007, 33, 267–349. [Google Scholar]
- Parker, C. Observations on the current status of Orobanche and Striga problems worldwide. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Holub, J. New names inPhanerogamae 6. Folia Geobot. Phytotaxon. 1977, 12, 417–432. [Google Scholar] [CrossRef]
- Sojak, J. Nomenklatorické poznámky (Phanerogamae). Casopis 1972, 140, 127–134. [Google Scholar]
- Teryokhin, E. Weed Broomrapes: Systematics, Ontogenesis, Biology, Evolution; Aufstieg Verlag: Lanshut, Germany, 1997. [Google Scholar]
- Joel, D. The new nomenclature of Orobanche and Phelipanche. Weed Res. 2009, 49, 6–7. [Google Scholar] [CrossRef]
- Schneeweiss, G.M. Correlated evolution of life history and host range in the nonphotosynthetic parasitic flowering plants Orobanche and Phelipanche (Orobanchaceae). J. Evol. Biol. 2007, 20, 471–478. [Google Scholar] [CrossRef]
- Castroviejo, S.; Muñoz Garmendia, F. Flora Ibérica: Plantas Vasculares de la Península Ibérica e Islas Baleares: Vol. IX; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2015; pp. 1–612. [Google Scholar]
- Parker, C. The parasitic weeds of the Orobanchaceae. In Parasitic Orobanchaceae; Springer: Berlin/Heidelberg, Germany, 2013; pp. 313–344. [Google Scholar]
- FAO. The Problem of Orobanche spp. in Africa and Near East. Food and Agriculture Organization of the United Nations, 2014. Available online: http://www.fao.org/agriculture/crops/thematic-sitemap/theme/biodiversity/weeds/issues/oro/en/ (accessed on 19 December 2022).
- Labrada, R. Farmer training on parasitic weed management. In Progress on Farmer Training in Parasitic Weed Management; Labrada, R., Ed.; FAO: Rome, Italy, 2007; pp. 1–5. [Google Scholar]
- Sauerborn, J. Parasitic Flowering Plants: Ecology and Management; Verlag Josef Margraf: Wiekersheim, Germany, 1991. [Google Scholar]
- Stewart, G.R.; Press, M.C. The physiology and biochemistry of parasitic angiosperms. Annu. Rev. Plant Biol. 1990, 41, 127–151. [Google Scholar] [CrossRef]
- Mabsoute, L.; Saadaoui, E. Acquis de recherche sur le parasitisme des légumineuses alimentaires au Maroc: Synthese bibliographique. Al Awamia 1996, 92, 55–67. [Google Scholar]
- Ennami, M.; Briache, F.Z.; Gaboun, F.; Abdelwahd, R.; Ghaouti, L.; Belqadi, L.; Westwood, J.; Mentag, R. Host differentiation and variability of Orobanche crenata populations from legume species in Morocco as revealed by cross-infestation and molecular analysis. Pest Manag. Sci. 2017, 73, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Nabloussi, A.; Velasco, L.; Assissel, N. First report of sunflower broomrape, Orobanche cumana Wallr., in Morocco. Plant Dis. 2018, 102, 457. [Google Scholar] [CrossRef]
- Rubiales, D.; Sadiki, M.; Román, B. First report of Orobanche foetida on common vetch (Vicia sativa) in Morocco. Plant Dis. 2005, 89, 528. [Google Scholar] [CrossRef] [PubMed]
- Vaz Patto, M.; Díaz-Ruiz, R.; Satovic, Z.; Román, B.; Pujadas-Salvà, A.; Rubiales, D. Genetic diversity of Moroccan populations of Orobanche foetida: Evolving from parasitising wild hosts to crop plants. Weed Res. 2008, 48, 179–186. [Google Scholar] [CrossRef]
- Zehhar, N. Recherche des Mécanismes Impliqués dans la Germination d’Orobanche ramosa L. et de Sources et de Mécanismes de Résistance à ce Parasite; Etude Préliminaire de Possibilités de Lutte Chimique Contre Orobanche ramosa et Cuscuta Lupuliformis Krock. Master’s Thesis, Université Sidi Mohamed Ben Abdellah, Fès, Morocco, 2003. [Google Scholar]
- Briache, F.Z.; Ennami, M.; Mbasani-Mansi, J.; Gaboun, F.; Abdelwahd, R.; Fatemi, Z.E.A.; El-Rodeny, W.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Field and controlled conditions screenings of some faba bean (Vicia faba L.) genotypes for resistance to the parasitic plant Orobanche crenata Forsk. and investigation of involved resistance mechanisms. J. Plant Dis. Prot. 2019, 126, 211–224. [Google Scholar] [CrossRef]
- Briache, F.Z.; Ennami, M.; Mbasani-Mansi, J.; Lozzi, A.; Abousalim, A.; Rodeny, W.E.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Effects of salicylic acid and indole acetic acid exogenous applications on induction of faba bean resistance against Orobanche crenata. Plant Pathol. J. 2020, 36, 476–490. [Google Scholar] [CrossRef]
- Ennami, M.; Briache, F.; Mansi, J.; Gaboun, F.; Ghaouti, L.; Belqadi, L.; Mentag, R. Genetic diversity of Moroccan Orobanche crenata populations revealed by sequence-related amplified polymorphism markers. J. Agric. Sci. 2017, 9, 164. [Google Scholar] [CrossRef]
- Ennami, M.; Mbasani-mansi, J.; Briache, F.Z.; Oussible, N.; Gaboun, F.; Ghaouti, L.; Belqadi, L.; Ghanem, M.E.; Aberkani, K.; Westwood, J.; et al. Growth-defense tradeoffs and source-sink relationship during both faba bean and lentil interactions with Orobanche crenata Forsk. Crop Prot. 2020, 127. [Google Scholar] [CrossRef]
- Mbasani-Mansi, J.; Briache, F.Z.; Ennami, M.; Gaboun, F.; Benbrahim, N.; Triqui, Z.E.A.; Mentag, R. Resistance of Moroccan lentil genotypes to Orobanche crenata infestation. J. Crop Improv. 2019, 33, 306–326. [Google Scholar] [CrossRef]
- Abdallah, F.; Kumar, S.; Amri, A.; Mentag, R.; Kehel, Z.; Mejri, R.K.; Triqui, Z.E.A.; Hejjaoui, K.; Baum, M.; Amri, M. Wild Lathyrus species as a great source of resistance for introgression into cultivated grass pea (Lathyrus sativus L.) against broomrape weeds (Orobanche crenata Forsk. and Orobanche foetida Poir.). Crop Sci. 2021, 61, 263–276. [Google Scholar] [CrossRef]
- Briache, F.Z.; El Amri, M.; Ennami, M.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Induction of systemic resistance to Orobanche crenata in lentil by exogenous application of salicylic acid and indole acetic acid. J. Plant Prot. Res. 2023, 63, 83–96. [Google Scholar]
- Bayoumi, T.; Ammar, S.M.; El-Bramawy, M.; Emam, M. Effect of some broomrape control methods on growth and seed yield attributes of faba bean Vicia faba, L. cultivars. J. Plant Prod. Sci. 2013, 1, 17–24. [Google Scholar]
- Fenández-Aparicio, M.; Sillero, J.C.; Rubiales, D. Intercropping with cereals reduces infection by Orobanche crenata in legumes. Crop Prot. 2007, 26, 1166–1172. [Google Scholar] [CrossRef]
- Amsellem, Z.; Barghouthi, S.; Cohen, B.; Goldwasser, Y.; Gressel, J.; Hornok, L.; Kerenyi, Z.; Kleifeld, Y.; Klein, O.; Kroschel, J. Recent advances in the biocontrol of Orobanche (broomrape) species. BioControl 2001, 46, 211–228. [Google Scholar] [CrossRef]
- Amri, M.; Trabelsi, I.; Abbes, Z.; Kharrat, M. Release of a new faba bean variety “Chourouk” resistant to the parasitic plants Orobanche foetida and O. crenata in Tunisia. Int. J. Agric. Biol. 2019, 21, 499–505. [Google Scholar] [CrossRef]
- Amri, M.; Abbes, Z.; Trabelsi, I.; Ghanem, M.E.; Mentag, R.; Kharrat, M. Chlorophyll content and fluorescence as physiological parameters for monitoring Orobanche foetida Poir. infection in faba bean. PLoS ONE 2021, 16, e0241527. [Google Scholar] [CrossRef]
- Lassner, M.W.; Peterson, P.; Yoder, J.I. Simultaneous amplification of multiple DNA fragments by polymerase chain reaction in the analysis of transgenic plants and their progeny. Plant Mol. Biol. Report. 1989, 7, 116–128. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef]
- Swofford, D. PAUP* Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Piwowarczyk, R.; Schneider, A.C.; Góralski, G.; Kwolek, D.; Denysenko-Bennett, M.; Burda, A.; Ruraż, K.; Joachimiak, A.J.; Pedraja, Ó.S. Phylogeny and historical biogeography analysis support Caucasian and Mediterranean centres of origin of key holoparasitic Orobancheae (Orobanchaceae) lineages. PhytoKeys 2021, 174, 165. [Google Scholar] [CrossRef] [PubMed]
- Schneeweiss, G.M.; Colwell, A.; Park, J.-M.; Jang, C.-G.; Stuessy, T.F. Phylogeny of holoparasitic Orobanche (Orobanchaceae) inferred from nuclear ITS sequences. Mol. Phylogenet. Evol. 2004, 30, 465–478. [Google Scholar] [CrossRef]
- Parsai, S.Z.M.; Vaezi, J.; Ejtehadi, H.; Memariani, F.; Joharchi, M.R. Phelipanche pouyanii (Orobanchaceae), a new species from Iran. Nova Biol. Reper. 2021, 7, 457–467. [Google Scholar]
- Wolfe, A.D.; Randle, C.P.; Liu, L.; Steiner, K.E. Phylogeny and biogeography of Orobanchaceae. Folia Geobot. 2005, 40, 115–134. [Google Scholar] [CrossRef]
- Abdellaoui, M.; Kasrati, A.; El Rhaffari, L. The effect of domestication on seed yield, essential oil yield and antioxidant activities of fennel seed (Foeniculum vulgare Mill.) grown in Moroccan oasis. J. Assoc. Arab Univ. Basic Appl. Sci. 2017, 24, 107–114. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Piwowarczyk, R.; Denysenko-Bennett, M.; Góralski, G.; Kwolek, D.; Pedraja, Ó.S.; Cygan, M.; Joachimiak, A. Phylogenetic relationships within Orobanche and Phelipanche (Orobanchaceae) from Central Europe, focused on problematic aggregates, taxonomy, and host ranges. Acta Biol. Cracoviensia Ser. Bot. 2018, 60, 45–64. [Google Scholar]
- Li, X.; Feng, T.; Randle, C.; Schneeweiss, G.M. Phylogenetic relationships in Orobanchaceae inferred from low-copy nuclear genes: Consolidation of major clades and identification of a novel position of the non-photosynthetic Orobanche clade sister to all other parasitic Orobanchaceae. Front. Plant Sci. 2019, 10, 902. [Google Scholar] [CrossRef]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenetics Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef]
- Pomel, A. Nouveaux Matériaux Pour la Flore Atlantique; Savy: Paris, France, 1874. [Google Scholar]
- Qasem, J.R.S. Thesium humile. In Parasitic Weeds of Jordan: Species, Hosts, Distribution and Management; Bentham Science Publishers: Sharjah, United Arab Emirates, 2022; pp. 304–314. [Google Scholar]
- Al-Mayah, A.A.; Al-Asadi, W.M. Two new records of the genus Phelipanche (Orobanchaceae) from Iraq. Glob. J. Biol. Agric. Health Sci. 2017, 6, 4–7. [Google Scholar] [CrossRef]
- Al-Asadi, W.M.; Al-Mayah, A.A. Three new specific records of Phelipanche (Orobanchaceae) to the Flora of Iraq. Calyx (mm) 2016, 5, 970–972. [Google Scholar]
- Conti, F.; Cancellieri, L.; Cangelmi, G.; Filibeck, G.; Rosati, L.; Bartolucci, F. New records of native and alien vascular plants from Abruzzo, Lazio and Molise National Park (Italy)–and additions to the flora of Abruzzo and Molise administrative regions. Ann. Di Bot. 2022, 12, 23–34. [Google Scholar]
- Domina, G.; Marino, P.; Castellano, G. The genus Orobanche (Orobanchaceae) in Sicily. Flora Mediterr. 2011, 21, 205–242. [Google Scholar]
- Raimondo, F.M.; Spadaro, V. Addenda et emendanda to the “A catalogue of plants growing in Sicily”. Flora Mediterr. 2009, 19, 303–312. [Google Scholar]
- Ruiz, L.C.; Gallo, M.L.; Moral, G.M.; Pedraja, O.S. Phelipanche cernua Pomel (Orobanchaceae), a prioritary name for the western mediterranean species recently redescribed as Ph. inexpectata. Flora Montiberica 2013, 54, 75–83. [Google Scholar]
- Castillejo, M.; FERNÁNDEZ-APARICIO, M.; Satovic, Z.; Rubiales, D. Comparative proteomic analysis of Orobanche and Phelipanche species inferred from seed proteins. Weed Res. 2009, 49, 81–87. [Google Scholar] [CrossRef]
- Navarro, J.G.; Gisbert, J.B.P.; Franzi, A.V.; Duato, E.S.; Gimeno, R.R. Las plantas de interés del NE de la Provincia de Albacete e inmediaciones de la Provincia de Valencia, VI. Sabuco: Rev. Estud. Albacet. 2011, 8, 117–142. [Google Scholar]
- Morales, R. Catálogo de plantas vasculares de la Comunidad de Madrid (España)[Catalogue of the vascular plants from Madrid Community (Spain)]. Bot. Complut 2003, 27, 31–70. [Google Scholar]
- Carlón, L.; Gómez Casares, G.; Laínz, M.; Moreno Moral, G.; Sánchez Pedraja, Ó.; Schneeweiss, G.M. Más, a Propósito de Algunas Phelipanche Pomel, Boulardia FW Schultz y Orobanche L. (Orobanchaceae) del Oeste del Paleártico; Jardín Botánico Atlántico: Gijón, Spain, 2008. [Google Scholar]
- Foley, M.Y. Orobanchaceae in the” flora iberica” área: New taxa, excluded-taxa, and typification. In Anales del Jardín Botánico de Madrid; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2001; pp. 223–233. [Google Scholar]
- Qasem, J.R.S. Parasitic Weeds of Jordan: Species, Hosts, Distribution and Management-Part I: Root Parasites; Orobanchaceae, Santalaceae & Cynomoryaceae; Bentham Science Publishers: Sharjah, United Arab Emirates, 2022; p. 360. [Google Scholar]
- Qasem, J.R. Parasitic weeds of the Orobanchaceae family and their natural hosts in Jordan. Weed Biol. Manag. 2009, 9, 112–122. [Google Scholar] [CrossRef]
- Qasem, J.R. Parasitic flowering plants on cultivated plants in Jordan-the present status and management. Pak. J. Weed Sci. Res. 2010, 16, 227–239. [Google Scholar]
- Qasem, J.; Foy, C. Screening studies on the host range of branched broomrape (Orobanche ramosa). J. Hortic. Sci. Biotechnol. 2007, 82, 885–892. [Google Scholar] [CrossRef]
- Babiker, A.; Ahmed, E.; Dawoud, D.; Abdella, N. Orobanche species in Sudan: History, distribution and management. Sudan J. Agric. Res. 2007, 10, 107–114. [Google Scholar]
- Lops, F.; Frabboni, L.; Carlucci, A.; Tarantino, A.; Raimondo, M.L.; Disciglio, G. Management of Branched Broomrape in Field Processing Tomato Crop; IntechOpen: London, UK, 2022; Available online: https://www.intechopen.com/chapters/82794 (accessed on 18 February 2023).
- Conversa, G.; Bonasia, A.; Elia, A. Chemical control of branched broomrape in processing tomato using sulfonylureas in Southern Italy. Ital. J. Agron. 2017, 12, 939. [Google Scholar] [CrossRef]
- Khatri, G.; Jha, P.; Agrawal, V.; Jacobsohn, R. Distribution, host range and phenology of Orobanche spp. in Nepal. In Proceedings of the International Workshop on Orobanche Research, Obermarchtal, Germany, 19–22 August 1989; pp. 18–24. [Google Scholar]
- Foy, C.L.; Jain, R.; Jacobsohn, R. Recent approaches for chemical control of broomrape (Orobanche spp.). Rev. Weed Sci. 1989, 4, 123–152. [Google Scholar]
- Satovic, Z.; Joel, D.M.; Rubiales, D.; Cubero, J.I.; Romn, B. Population genetics in weedy species of Orobanche. Australas. Plant Pathol. 2009, 38, 228–234. [Google Scholar] [CrossRef]
- Darvishzadeh, R. Genetic variability, structure analysis, and association mapping of resistance to broomrape (Orobanche aegyptiaca Pers.) in tobacco. J. Agric. Sci. Technol. 2018, 18, 1407–1418. [Google Scholar]
- Hamrick, J.L.; Godt, M.W. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar]
- Aboukhalaf, A.; El Amraoui, B.; Tabatou, M.; da Rocha, J.M.F.; Belahsen, R. Screening of the antimicrobial activity of some extracts of edible wild plants in Morocco. Funct. Foods Health Dis. 2020, 10, 265–273. [Google Scholar] [CrossRef]
- El Ouariachi, E.; Lahhit, N.; Bouyanzer, A.; Hammouti, B.; Paolini, J.; Majidi, L.; Desjobert, J.; Costa, J. Chemical composition and antioxidant activity of essential oils and solvent extracts of Foeniculum vulgare Mill. from Morocco. J. Chem. Pharm. Res. 2014, 6, 743–748. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Locality | Voucher Information or Reference | GenBank Accession |
|---|---|---|---|
| Phelipanche schultzii Mutel (Pomel.) | Morocco | OA | OM475767 |
| P. schultzii Mutel (Pomel.) | Morocco | OY-4 | OM475768 |
| P. schultzii Mutel (Pomel.) | Spain | [41] | MT026593 |
| P. gratiosa | Spain | herb. Sanchez Pedraja 12463 | EU581785 |
| P. gratiosa | Spain | herb. Sanchez Pedraja 12462 | EU581774 |
| P. cf. aegyptiaca Pers. | Georgia | [42] | AY209327 |
| P. cf. aegyptiaca Pers. | Georgia | [42] | AY209328 |
| P. coelestis (Reut.) G. Beck | Turkey | [42] | AY209329 |
| P. coelestis | Armenia | [41] | MT026616 |
| P. portoilicitana | Spain | [41] | AY209329 |
| P. portoilicitana | Armenia | [41] | MT026675 |
| P. cf. iberica (Beck) Soják | Turkey | WU:Albach 682 | AY960742 |
| P. cf. iberica (Beck) Soják | ? | [43] | LT715380 |
| P. nana (Noe ex Reut.) Nyman | Greece | [42] | AY209316 |
| P. nana (Noe ex Reut.) Nyman | Greece | [42] | AY209313 |
| P. ramosa L. (Pomel.) | Bulgaria | * | MK024289 |
| P. ramosa L. (Pomel.) | Bulgaria | * | MK024288 |
| P. cf. rosmarina | Greece | [42] | AY209335 |
| P. cf. rosmarina | Spain | [42] | AY209334 |
| Digitalis purpurea | ? | [44] | AY911261 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Amri, M.; Amri, M.; Kadir, E.M.; Triqui, Z.E.A.; Khayi, S.; Mentag, R. First Report of the Branched Broomrape (Phelipanche schultzii (Mutel) Pomel.) on Fennel (Foeniculum vulgare Mill.) in Morocco. Horticulturae 2023, 9, 567. https://doi.org/10.3390/horticulturae9050567
El Amri M, Amri M, Kadir EM, Triqui ZEA, Khayi S, Mentag R. First Report of the Branched Broomrape (Phelipanche schultzii (Mutel) Pomel.) on Fennel (Foeniculum vulgare Mill.) in Morocco. Horticulturae. 2023; 9(5):567. https://doi.org/10.3390/horticulturae9050567
Chicago/Turabian StyleEl Amri, Majda, Moez Amri, El Mostafa Kadir, Zine El Abidine Triqui, Slimane Khayi, and Rachid Mentag. 2023. "First Report of the Branched Broomrape (Phelipanche schultzii (Mutel) Pomel.) on Fennel (Foeniculum vulgare Mill.) in Morocco" Horticulturae 9, no. 5: 567. https://doi.org/10.3390/horticulturae9050567
APA StyleEl Amri, M., Amri, M., Kadir, E. M., Triqui, Z. E. A., Khayi, S., & Mentag, R. (2023). First Report of the Branched Broomrape (Phelipanche schultzii (Mutel) Pomel.) on Fennel (Foeniculum vulgare Mill.) in Morocco. Horticulturae, 9(5), 567. https://doi.org/10.3390/horticulturae9050567