
Differences in Dry Matter Accumulation and Distribution Patterns between Pre-Elite Seed and Certified Seed of Virus-Free Potato

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Materials
2.2. Experimental Design and Management
2.3. Plant Sampling and Measurements
2.3.1. Proportion of Dry Matter Accumulation and Distribution
2.3.2. Yield and Its Composition
2.4. Calculation Formula
2.5. Data Analysis
3. Results
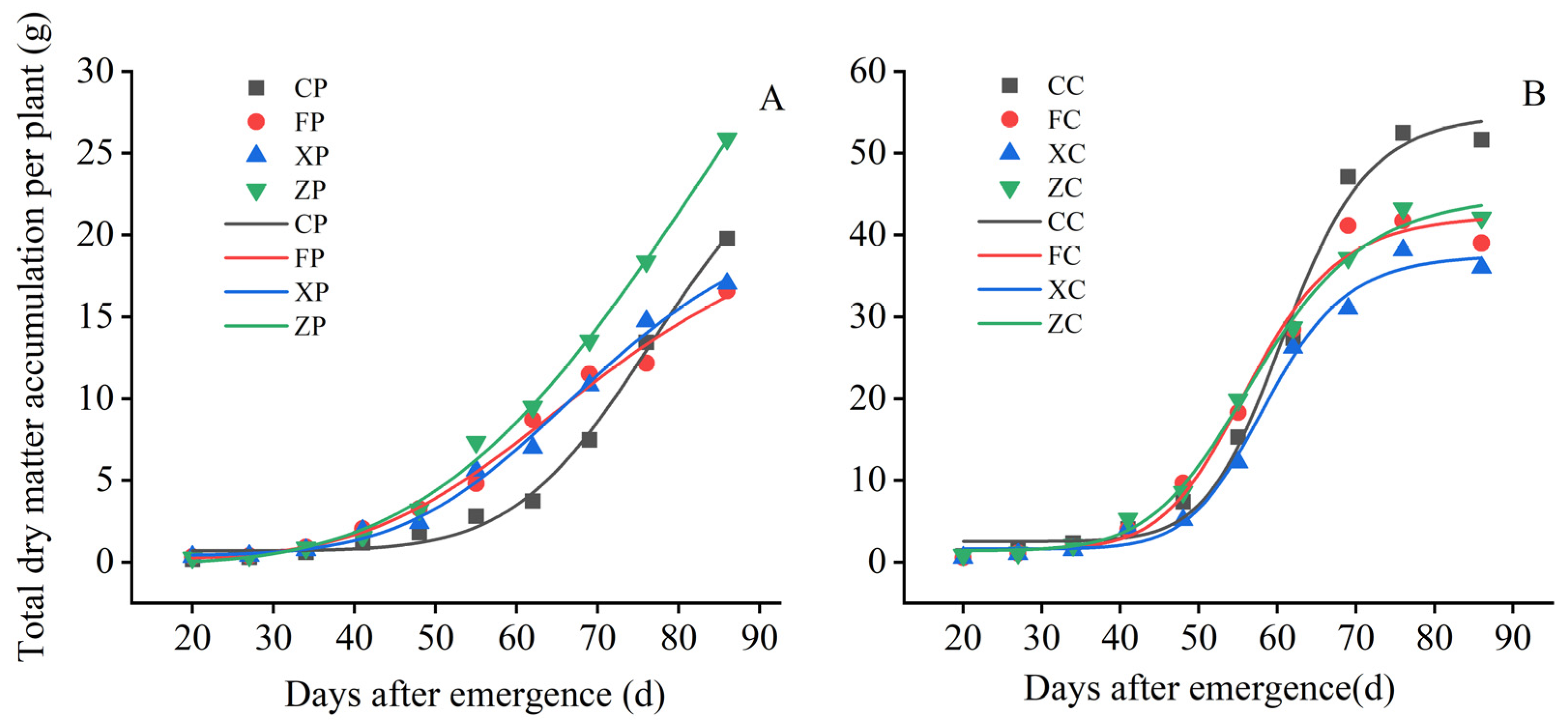
3.1. Comparison of the Growth Process of Pre-Elite and Certified Seeds
3.2. Comparison of the Changing Pattern of Dry Matter Accumulation between the Pre-Elite and Certified Seeds
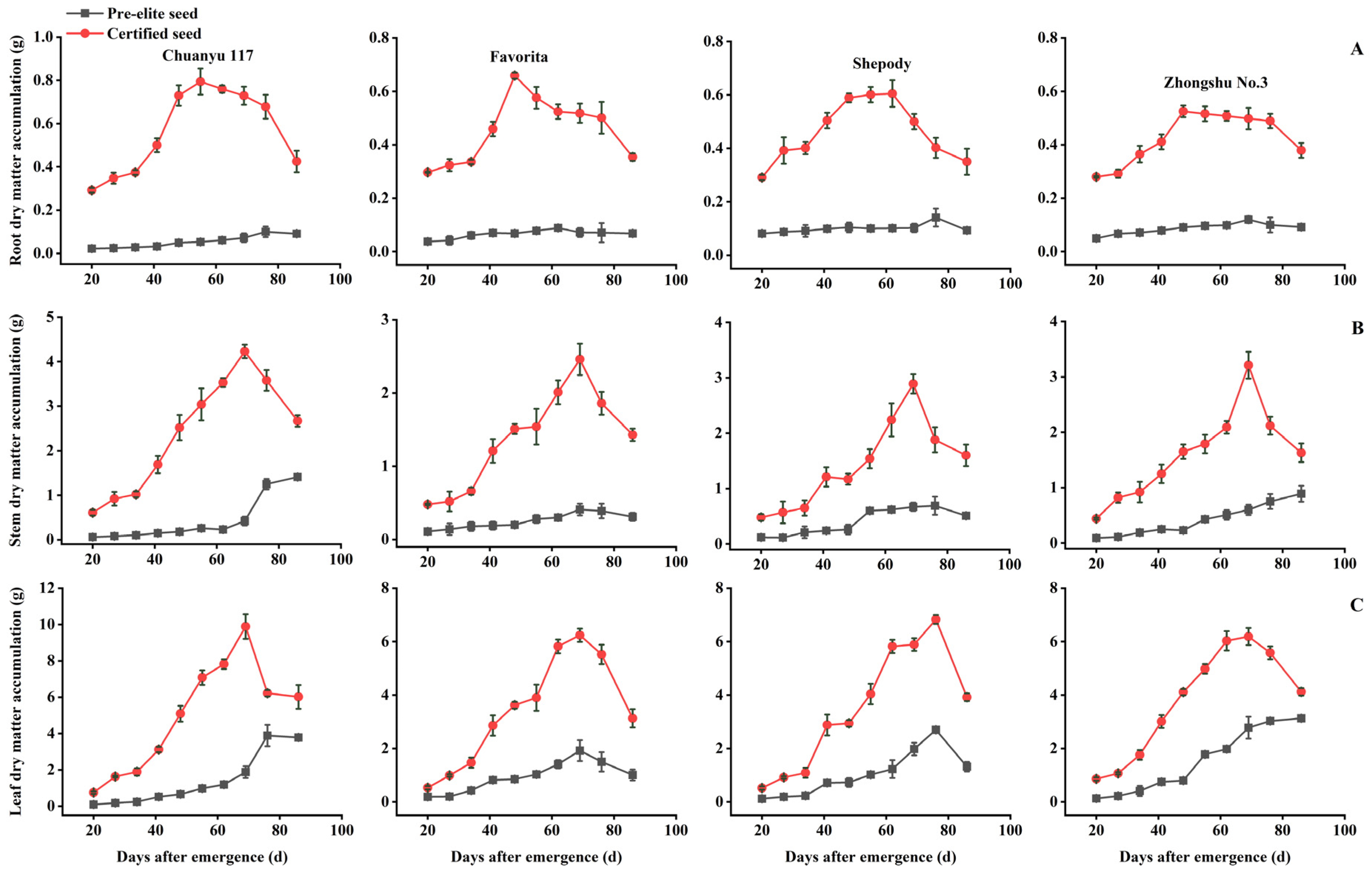
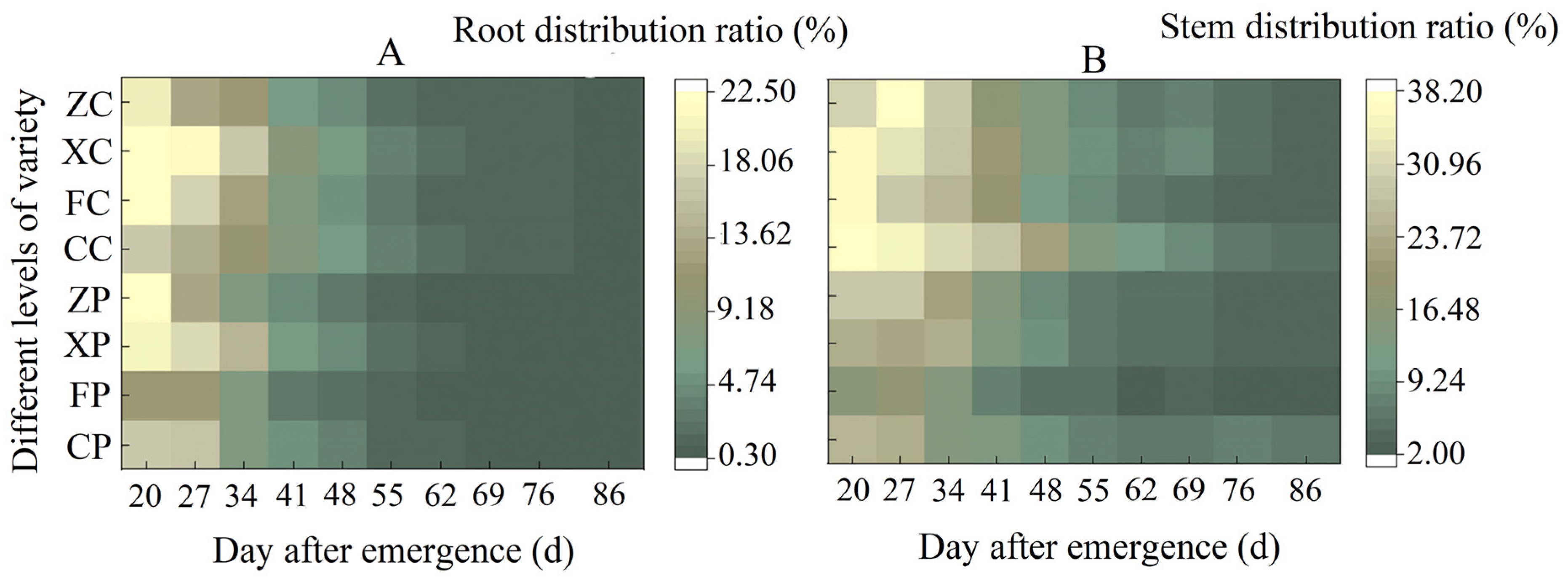
3.2.1. Comparison of the Changing Pattern of Dry Matter Accumulation in Roots, Stems, and Leaves
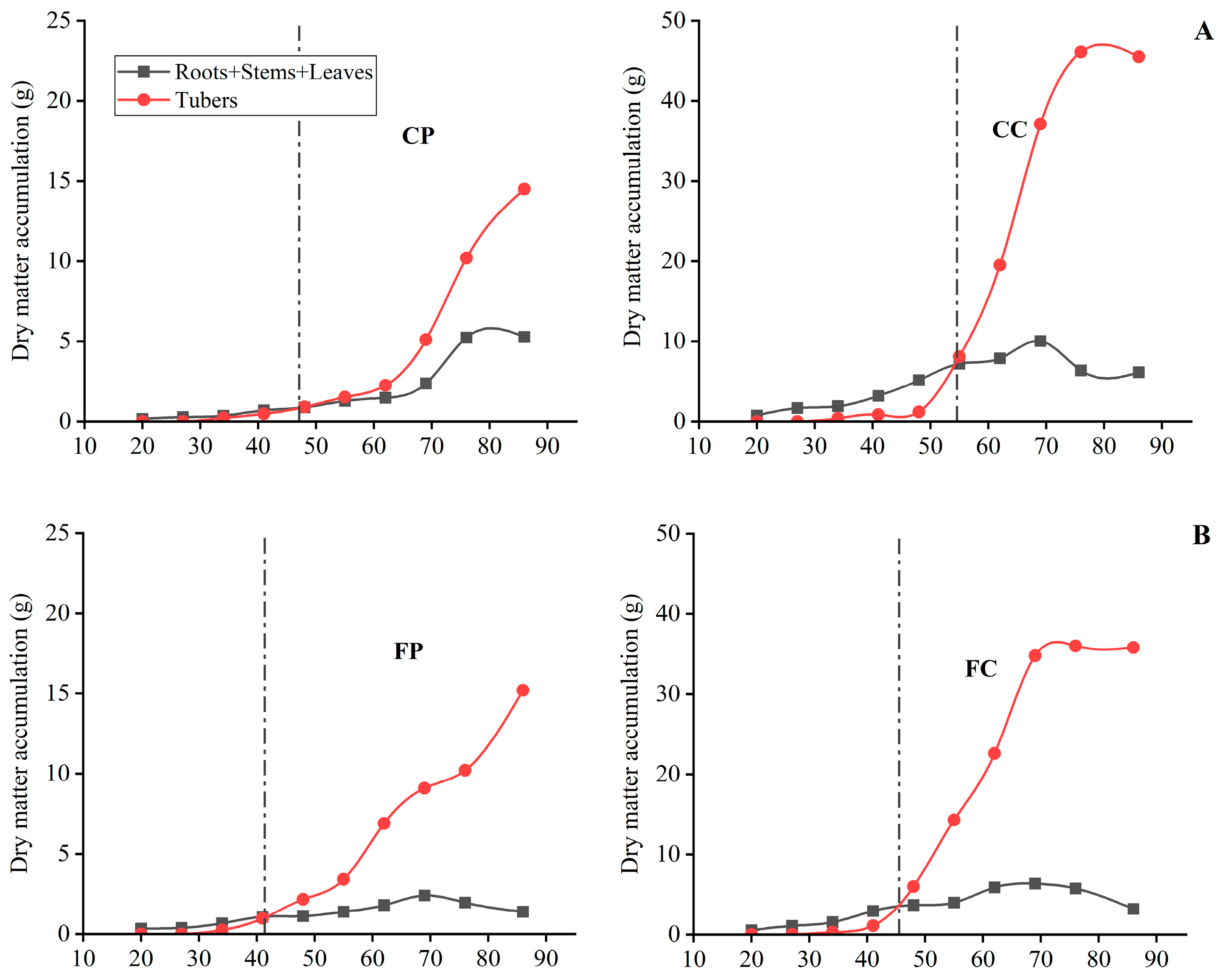
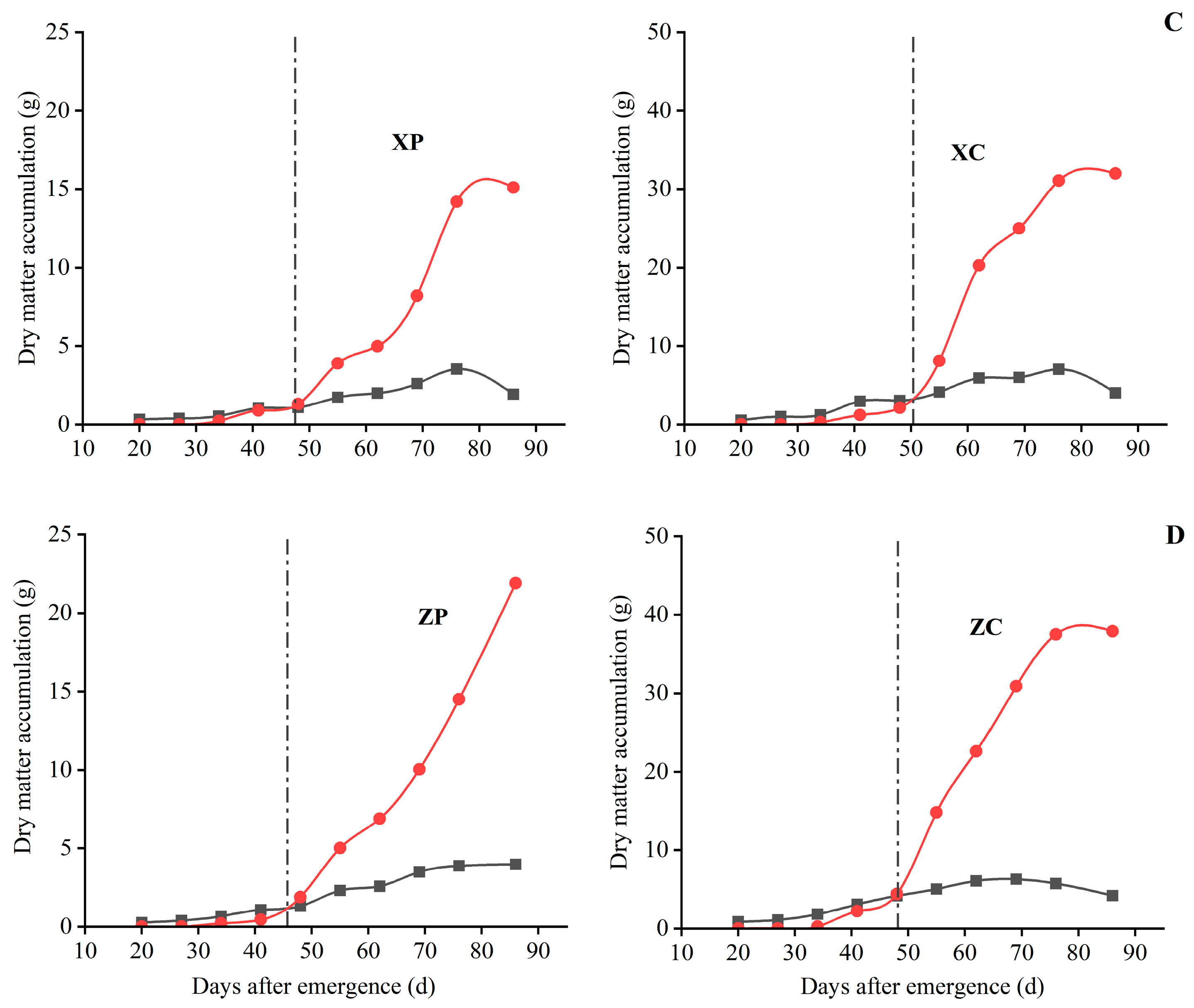
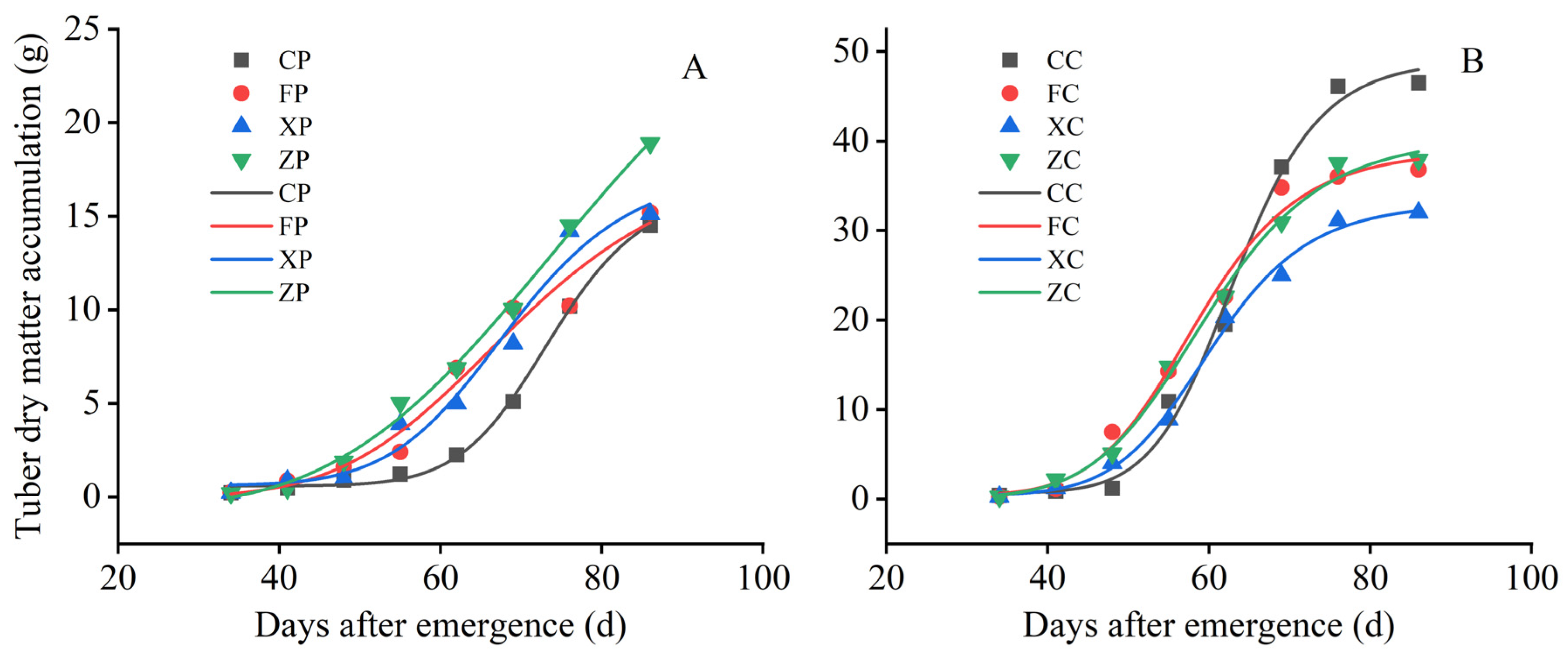
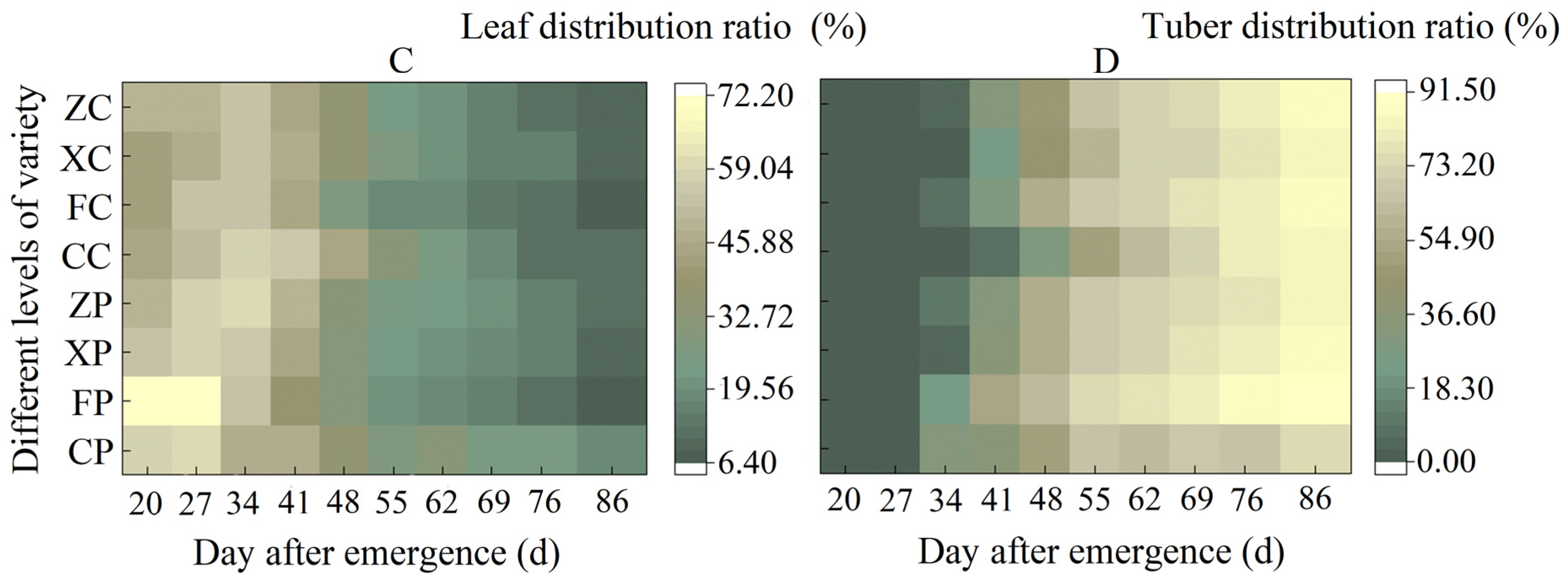
3.2.2. Comparison of the Changing Pattern of Dry Matter Accumulation in Tubers
3.2.3. Comparison of the Changing Pattern of Dry Matter Accumulation in the Whole Plants
3.3. Comparison of Dry Matter Distribution Patterns among Different Organs
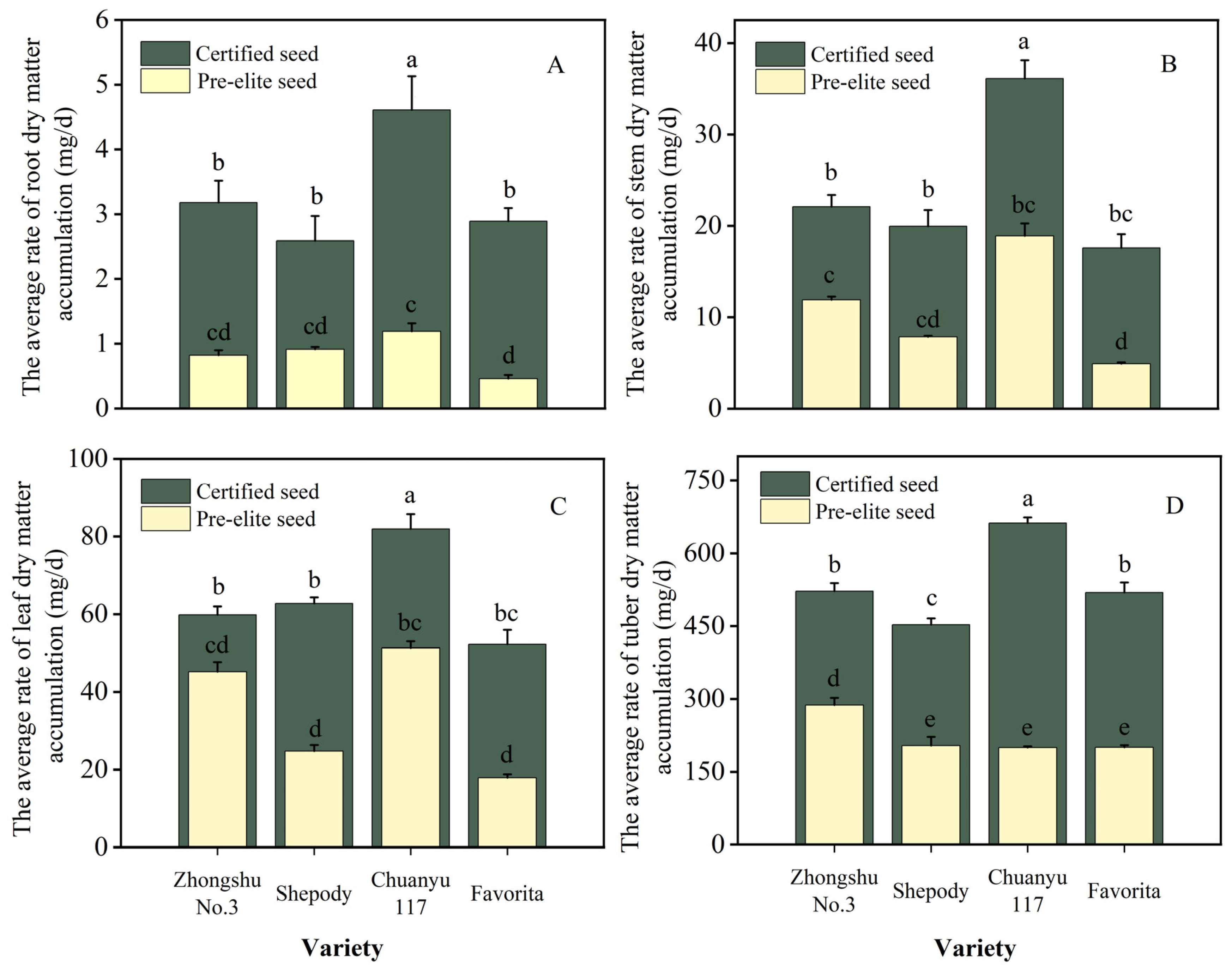
3.4. Comparison of the Dry Matter Accumulation Rate of Different Organs between the Pre-Elite and Certified Seeds
3.5. Differences in Theoretical Yield and Its Constituent Factors between the Pre-Elite and Certified Seeds of Different Varieties
4. Discussion
4.1. Differences in Dry Matter Accumulation Patterns between the Pre-Elite and Certified Seeds
4.2. Differences in Dry Matter Distribution Patterns between the Pre-Elite and Certified Seeds
4.3. Differences in Yields between the Pre-Elite and Certified Seeds
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- André, D.; Jean-Pierre, G.; Peter, K.; Jorge, A.-P.; Vivian, P.; Guy, H. Correction to: The potato of the future: Opportunities and challenges in sustainable agri-food systems. Potato Res. 2021, 64, 681–720. [Google Scholar] [CrossRef]
- Quéré, B. Seeds for the Future = Challenges for Potato. Potato Res. 2017, 60, 327–329. [Google Scholar] [CrossRef]
- Hong, L.; Xinyu, Y.; Yichen, K.; Wenlin, L.; Haojun, L.; Shuhao, Q. Effects of nitrogen, phosphorus and potassium combined fertilisation on the dry matter accumulation, distribution and yield of potato under ridge and furrow film mulch cropping. Potato Res. 2022, 10, 277–281. [Google Scholar] [CrossRef]
- Silva-Díaz, C.; Ramírez, D.A.; Rinza, J.; Ninanya, J.; Loayza, H.; Gómez, R.; Anglin, N.L.; Eyzaguirre, R.; Quiroz, R. Radiation interception, conversion and partitioning efficiency in potato landraces: How Far Are We from the Optimum? Plants 2020, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.I.; Magnitskiy, S.; Rodríguez, L.E. Nitrogen, phosphorus and potassium accumulation and partitioning by the potato group Andigenum in Colombia. Nutr. Cycl. Agroecosyst. 2019, 113, 349–363. [Google Scholar] [CrossRef]
- He, W.; Li, J.; Pu, M.; Xu, Z.-G.; Gan, L. Response of photosynthate distribution in potato plants to different LED spectra. Funct. Plant Biol. 2020, 47, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Aliche, E.B.; Theeuwen, T.P.J.M.; Oortwijn, M.; Visser, R.G.F.; van der Linden, C.G. Carbon partitioning mechanisms in POTATO under drought stress. Plant Physiol. Biochem. 2019, 146, 211–219. [Google Scholar] [CrossRef]
- Golovko, T.K.; Tabalenkova, G.N. Source–Sink Relationships in Potato Plants. Russ. J. Plant Physiol. 2019, 66, 664–671. [Google Scholar] [CrossRef]
- Jia, L.; Wu, L.; Suyala, Q.; Shi, X.; Qin, Y.; Fan, M. Promotion of potato yield under moderate water deficiency at the seedling stage by modifying sink-source relationship. Plant Prod. Sci. 2021, 25, 95–104. [Google Scholar] [CrossRef]
- Aliche, E.B.; Prusova-Bourke, A.; Ruiz-Sanchez, M.; Oortwijn, M.; Gerkema, E.; Van As, H.; Visser, R.G.F.; van der Linden, C.G. Morphological and physiological responses of the potato stem transport tissues to dehydration stress. Planta 2020, 251, 45. [Google Scholar] [CrossRef]
- Fujibayashi, M.; Suzuki, T.; Sano, T. Mechanism underlying potato spindle tuber viroid affecting tomato (Solanum lycopersicum): Loss of control over reactive oxygen species production. J. Gen. Plant Pathol. 2021, 87, 226–235. [Google Scholar] [CrossRef]
- Tatarowska, B.; Plich, J.; Milczarek, D.; Flis, B. Temperature-dependent resistance to potato virus M in potato (Solanum tuberosum). Plant Pathol. 2020, 69, 1445–1452. [Google Scholar] [CrossRef]
- Gerieva, F.; Basiev, S.; Khutinaev, O.; Basieva, A. The growing of minituber using the aerohydroponic method of potato plant cultivating. IOP Conf. Ser. Earth Environ. Sci. 2019, 10, 111–115. [Google Scholar] [CrossRef]
- Tokbergenova, Z.A.; Babayev, S.A.; Togayeva, D.U.; Kudusbekova, D.Z.; Zagurskii, A.V. Efficiency of microtubers application in the production of original potato seeds. Online J. Biol. Sci. 2017, 17, 316–322. [Google Scholar] [CrossRef]
- Ali, O.; Mehmet Emin, Ç. Optimization and economic perspective of planting density and minituber size in potato seed production. Am. J. Potato Res. 2023, 100, 169–183. [Google Scholar] [CrossRef]
- Le Hingrat, Y.; Quéré, B.; Andrivon, D.; Kerlan, M.-C. Report on the workshop “Research challenges for the seed potato sector”. Potato Res. 2017, 60, 331–335. [Google Scholar] [CrossRef]
- Abuhena, M.; Al-Rashid, J.; Azim, M.F.; Khan, M.N.M.; Kabir, M.G.; Barman, N.C.; Rasul, N.M.; Akter, S.; Huq, M.A. Optimization of industrial (3000 L) production of Bacillus subtilis CW-S and its novel application for minituber and industrial-grade potato cultivation. Sci. Rep. 2022, 12, 11153. [Google Scholar] [CrossRef] [PubMed]
- Fulladolsa, A.C.; LaPlant, K.E.; Groves, R.L.; Charkowski, A.O. Potato plants grown from minitubers are delayed in maturity and lower in yield, but are not at a higher risk of potato virus Y infection than plants grown from conventional seed. Am. J. Potato Res. 2017, 95, 45–53. [Google Scholar] [CrossRef]
- Sharifi, M.; Zebarth, B.J.; Hajabbasi, M.A.; Kalbasi, M. Dry matter and Nitrogen accumulation and root morphological characteristics of two clonal selections of ‘Russet Norkotah’ potato as affected by Nitrogen fertilization. J. Plant Nutr. 2005, 25, 2243–2253. [Google Scholar] [CrossRef]
- Ma, Y.; Chai, Y.; Guan, S.; Tuerti, T.; Liu, H.; Zhang, Z.; Diao, M. Effects of different levels of nitrogen supply on key enzyme activities of Nitrogen metabolism and growth stimulation of endive (Cichorium endivia L.). Sustainability 2022, 14, 15159. [Google Scholar] [CrossRef]
- Li, C.; Luo, X.; Li, Y.; Wang, N.; Zhang, T.; Dong, Q.; Feng, H.; Zhang, W.; Siddique, K.H. Ridge planting with transparent plastic mulching improves maize productivity by regulating the distribution and utilization of soil water, heat, and canopy radiation in arid irrigation area. Agric. Water Manag. 2023, 280, 108230. [Google Scholar] [CrossRef]
- Zhai, J.; Zhang, G.; Zhang, Y.; Xu, W.; Xie, R.; Ming, B.; Hou, P.; Wang, K.; Xue, J.; Li, S. Effect of the rate of Nitrogen application on dry matter accumulation and yield formation of densely planted Maize. Sustainability 2022, 14, 14940. [Google Scholar] [CrossRef]
- Cai, F.; Mi, N.; Ming, H.; Zhang, Y.; Zhang, H.; Zhang, S.; Zhao, X.; Zhang, B. Responses of dry matter accumulation and partitioning to drought and subsequent rewatering at different growth stages of maize in Northeast China. Front. Plant Sci. 2023, 14, 2023. [Google Scholar] [CrossRef] [PubMed]
- Woli, K.P.; Sawyer, J.E.; Boyer, M.J.; Abendroth, L.J.; Elmore, R.W. Corn Era Hybrid dry matter and macronutrient accumulation across development stages. Agron. J. 2017, 109, 751–761. [Google Scholar] [CrossRef]
- Fernandes, A.M.; Soratto, R.P.; Pilon, C. Soil phosphorus increases dry matter and Nutrient accumulation and allocation in potato cultivars. Am. J. Potato Res. 2015, 92, 117–127. [Google Scholar] [CrossRef]
- Xia, H.; Wang, L.; Qiao, Y.; Kong, W.; Xue, Y.; Wang, Z.; Kong, L.; Xue, Y.; Sizmur, T. Elucidating the source–sink relationships of zinc biofortification in wheat grains: A review. Food Energy Secur. 2020, 9, 243. [Google Scholar] [CrossRef]
- Jonik, C.; Sonnewald, U.; Hajirezaei, M.R.; Flügge, U.I.; Ludewig, F. Simultaneous boosting of source and sink capacities doubles tuber starch yield of potato plants. Plant Biotechnol. J. 2015, 10, 1088–1098. [Google Scholar] [CrossRef]
- Li, W.; Xiong, B.; Wang, S.; Deng, X.; Yin, L.; Li, H.J.P.O. Regulation effects of water and Nitrogen on the Source-Sink relationship in potato during the tuber bulking stage. PLoS ONE 2016, 11, e0146877. [Google Scholar] [CrossRef]
- Katoh, A.; Ashida, H.; Kasajima, I.; Shigeoka, S.; Yokota, A. Potato yield enhancement through intensification of sink and source performances. Breeding Sci. 2015, 65, 77–84. [Google Scholar] [CrossRef]
- Shaohui, Z.; Junliang, F.; Fucang, Z.; Haidong, W.; Ling, Y.; Xin, S.; Minghui, C.; Houliang, C.; Zhijun, L. Optimizing irrigation amount and potassium rate to simultaneously improve tuber yield, water productivity and plant potassium accumulation of drip-fertigated potato in northwest China. Agric. Water Manag. 2022, 65, 264. [Google Scholar] [CrossRef]
- Vos, J. Split nitrogen application in potato: Effects on accumulation of nitrogen and dry matter in the crop and on the soil nitrogen budget. J. Agric. Sci. 2002, 133, 263–274. [Google Scholar] [CrossRef]
- Lu, J.-W.; Qiu, H.-Z.; Zhang, W.-M.; Wang, D.; Zhang, J.-L.; Zhang, C.-H.; Hou, S.-Y. Characteristics of dry matter and potassium accumulation and distribution in potato plant in semi-arid rainfed areas. J. Appl. Ecol. 2013, 244, 23–30. [Google Scholar]
- Yubi, Y.; Jun, L.; Haiyang, N.; Xiuyun, Z. Collaborative influence of elevated CO2 concentration and high temperature on potato biomass accumulation and characteristics. Open Chem. 2019, 17, 728–737. [Google Scholar] [CrossRef]
- Rogers, C.W.; Dari, B.; Hu, G.; Mikkelsen, R. Dry matter production, nutrient accumulation, and nutrient partitioning of barley. J. Plant Nutr. Soil Sci. 2019, 182, 367–373. [Google Scholar] [CrossRef]
- Delazari, F.T.; Ferreira, M.G.; Silva, G.H.D.; Dariva, F.D.; Freitas, D.S.D.; Nick, C. Eficiência no useo da água e acúmulo de matéria na batata-doce em função de lâminas de ir-rigação. IRRIGA 2018, 45, 826–835. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, J.; Bhardwaj, V.; Sood, S.; Siddappa, S.; Goutam, U.; Dalamu, n.; Kardile, H.B.; Kumar, D.; Kumar, V. Genotypic variations for tuber nutrient content, dry matter and agronomic traits in tetraploid potato germplasm. Physiol. Mol. Biol. Plants 2022, 28, 1233–1248. [Google Scholar] [CrossRef]
- Geremew, E.B.; Steyn, J.M.; Annandale, J.G. Evaluation of growth performance and dry matter partitioning of four processing potato (Solanum tuberosum) cultivars. N. Z. J. Crop Hortic. Sci. 2007, 92, 117–127. [Google Scholar] [CrossRef]
- Soratto, R.P.; Fernandes, A.M. Phosphorus effects on biomass accumulation and nutrient uptake and removal in two potato cultivars. Agron. J. 2016, 108, 1225–1236. [Google Scholar] [CrossRef]
- Obiero, C.O.; Milroy, S.P.; Bell, R.W. Importance of whole plant dry matter dynamics for potato (Solanum tuberosum L.) tuber yield response to an episode of high temperature. Environ. Exp. Bot. 2019, 162, 560–571. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Wubante Negash, A.; Rogers, K.; Melkamu, E. Determination of optimum planting density of very small potato minitubers and appropriate pot size for maximizing seed tuber production in screenhouse conditions. Crop Sci. 2023, 11, 435–437. [Google Scholar] [CrossRef]
- Abbasian, A.; Ahmadi, A.; Abbasi, A.-R.; Darvishi, B. Effect of various phosphorus and calcium concentrations on potato seed tuber production. Plant Nutr. 2018, 14, 1765–1777. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Pre-Elite Seed | Certified Seed | ||||||
|---|---|---|---|---|---|---|---|---|
| Chuanyu 117 | Favorita | Shepody | Zhongshu No. 3 | Chuanyu 117 | Favorita | Shepody | Zhongshu No. 3 | |
| Maturity | Medium | Early | Medium | Early | Medium | Early | Medium | Early |
| Growth period (d) | - | - | - | - | 83 | 65 | 90 | 67 |
| Seed Level | Variety | Wm (g) | a | b | R2 | t0 (d) | t1 (d) | t2 (d) | Δt (d) | Vm (g d−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Pre-elite seed | Zhongshu No. 3 | 19.2224 | 7.4124 | −0.1103 | 0.9918 | 67 | 55 | 79 | 24 | 0.5303 |
| Shepody | 15.8358 | 9.6694 | −0.1458 | 0.9875 | 66 | 57 | 75 | 18 | 0.5774 | |
| Chuanyu 117 | 21.9536 | 9.1719 | −0.1177 | 0.9927 | 78 | 67 | 89 | 22 | 0.6461 | |
| Favorita | 16.8341 | 6.9710 | −0.1050 | 0.9975 | 66 | 54 | 79 | 25 | 0.4417 | |
| Certified seed | Zhongshu No. 3 | 38.2606 | 8.9549 | −0.1512 | 0.9978 | 59 | 51 | 68 | 17 | 1.4463 |
| Shepody | 32.0101 | 10.1848 | −0.1701 | 0.9958 | 60 | 52 | 68 | 15 | 1.3615 | |
| Chuanyu 117 | 48.0480 | 11.4907 | −0.1824 | 0.9949 | 63 | 56 | 70 | 14 | 2.1913 | |
| Favorita | 37.8004 | 9.5113 | −0.1631 | 0.9922 | 58 | 50 | 66 | 16 | 1.5408 |
| Seed Level | Variety | Wm (g) | a | b | R2 | t0 (d) | t1 (d) | t2 (d) | Δt (d) | Vm (g d−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Pre-elite seed | Zhongshu No. 3 | 24.8373 | 6.5261 | −0.0983 | 0.9959 | 66 | 53 | 80 | 27 | 0.6104 |
| Shepody | 18.6422 | 8.5085 | −0.1326 | 0.9795 | 64 | 54 | 74 | 20 | 0.6178 | |
| Chuanyu 117 | 27.6825 | 8.7059 | −0.1150 | 0.9886 | 76 | 64 | 86 | 22 | 0.7957 | |
| Favorita | 18.0456 | 6.4574 | −0.1025 | 0.9989 | 63 | 50 | 76 | 26 | 0.4624 | |
| Certified seed | Zhongshu No. 3 | 46.1289 | 7.2442 | −0.1302 | 0.9953 | 56 | 46 | 66 | 20 | 1.5012 |
| Shepody | 40.1361 | 8.0937 | −0.1427 | 0.9893 | 57 | 48 | 66 | 18 | 1.4314 | |
| Chuanyu 117 | 59.5946 | 7.9193 | −0.1343 | 0.9887 | 59 | 49 | 69 | 20 | 2.0009 | |
| Favorita | 45.0720 | 7.7984 | −0.1411 | 0.9854 | 55 | 46 | 65 | 19 | 1.5896 |
| Seed Level | Variety | Tuber Fresh Weight (g) | Tuber Volume (cm3) | Tuber Number (pcs) | Theoretical Yield (kg ha−1) |
|---|---|---|---|---|---|
| Pre-elite seed | Zhongshu No. 3 | 121.79 ± 23.13 cC | 112.73 ± 20.54 cC | 4.53 ± 0.42 dDE | 11,070.49 ± 2102.3 cC |
| Shepody | 76.54 ± 8.39 dD | 71.20 ± 7.79 dD | 3.27 ± 0.64 eDE | 6957.16 ± 762.58 dD | |
| Chuanyu 117 | 86.56 ± 14.72 dD | 81.20 ± 14.91 dCD | 9.73 ± 0.31 aA | 7868.66 ± 1337.77 dD | |
| Favorita | 85.39 ± 24.31 dD | 79.67 ± 22.85 dCD | 3.00 ± 0.35 eE | 7762.04 ± 2209.70 dD | |
| Certified seed | Zhongshu No. 3 | 183.27 ± 12.82 bB | 170.33 ± 11.55 bB | 6.40 ± 0.72 cC | 16,658.81 ± 1165.36 bB |
| Shepody | 164.80 ± 12.28 bB | 153.07 ± 11.50 bB | 4.80 ± 0.60 dD | 14,980.46 ± 1116.07 bB | |
| Chuanyu 117 | 250.74 ± 23.49 aA | 231.87 ± 29.27 aA | 8.27 ± 0.95 bAB | 22,792.51 ± 2135.14 aA | |
| Favorita | 173.65 ± 21.28 bB | 161.47 ± 27.37 bB | 7.40 ± 0.72 bcBC | 15,784.77 ± 1934.11 bB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Ma, H.; Fang, X.; Liu, R.; Shi, X.; Zhang, K.; Wang, Q.; Zheng, S. Differences in Dry Matter Accumulation and Distribution Patterns between Pre-Elite Seed and Certified Seed of Virus-Free Potato. Horticulturae 2023, 9, 644. https://doi.org/10.3390/horticulturae9060644
Wu C, Ma H, Fang X, Liu R, Shi X, Zhang K, Wang Q, Zheng S. Differences in Dry Matter Accumulation and Distribution Patterns between Pre-Elite Seed and Certified Seed of Virus-Free Potato. Horticulturae. 2023; 9(6):644. https://doi.org/10.3390/horticulturae9060644
Chicago/Turabian StyleWu, Chao, Haiyan Ma, Xiaoting Fang, Ruilin Liu, Xinxin Shi, Kaiqin Zhang, Qiang Wang, and Shunlin Zheng. 2023. "Differences in Dry Matter Accumulation and Distribution Patterns between Pre-Elite Seed and Certified Seed of Virus-Free Potato" Horticulturae 9, no. 6: 644. https://doi.org/10.3390/horticulturae9060644
APA StyleWu, C., Ma, H., Fang, X., Liu, R., Shi, X., Zhang, K., Wang, Q., & Zheng, S. (2023). Differences in Dry Matter Accumulation and Distribution Patterns between Pre-Elite Seed and Certified Seed of Virus-Free Potato. Horticulturae, 9(6), 644. https://doi.org/10.3390/horticulturae9060644