
Medicinal Use, Flower Trade, Preservation and Mass Propagation Techniques of Cymbidium Orchids—An Overview


 , , ,
, , ,
Abstract
:1. Introduction
2. Medicinal Value of Cymbidiums
3. Floristic Significance of Cymbidiums
4. Reproductive Biology in Cymbidiums
5. Seed Biology of Cymbidiums
6. Propagation of Cymbidiums
6.1. Conventional Methods
6.2. Clonal Propagation of Cymbidiums
6.3. Asymbiotic Seed Germination in Cymbidiums
6.4. Ex Vitro Seed Germination
6.5. Seedling Development
6.6. Culture Media
7. Clonal Propagation of Cymbidiums
7.1. Micropropagation of Cymbidiums
7.2. Propagation by Artificial Seeds
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zotz, G. The systematic distribution of vascular epiphytes-a critical update. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.M.; Sharma, M. Dual phase regeneration system for mass propagation of Cymbidium aloifolium (L.) Sw.: A High Value Medicinal Orchid. Plant Tissue Cult. Biotechnol. 2019, 29, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.M.; Goh, N.K.; Chia, L.S.; Chia, T.F. Recent advances in traditional plant drugs and orchids. Acta Pharm. Sin. 2003, 24, 7–21. [Google Scholar]
- Pant, B.; Swar, S. Micropropagation of Cymbidium iridioides Nepal. J. Sci. Technol. 2012, 12, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.K. Morphological diversity of the orchids of Orissa/Sarat Misra. In Orchids: Science and Commerce; Pathak, P., Sehgal, R.N., Shekhar, N., Sharma, M., Sood, A., Eds.; Bishen Singh Mahendra Pal Singh: New Delhi, India, 2001; p. 35. [Google Scholar]
- Teoh, E.S. India: Van Rheede, Caius and Others. In Orchids as aphrodisiac, medicine or food; Teoh, E.S., Ed.; Springer: Singapore, 2019; pp. 195–232. [Google Scholar] [CrossRef]
- Jana, S.K.; Sinha, G.P.; Chauhan, A.S. Ethnobotanical aspects of Orchids in Sikkim. J. Orchid Soc. India 1997, 11, 79–84. [Google Scholar]
- Wang, G.Q. National Chinese Herbal Medicine Collection; People’s Medical Publishing House: Beijing, China, 2014. [Google Scholar]
- Sujin, R.M.; Jeeva, S.; Subin, R.M. Cymbidium aloifolium: A review of its traditional uses, phytochemistry, and pharmacology. Phytochem. Pharmacol. Asp. Ethnomedicinal Plants 2021, 363–371. [Google Scholar]
- Kim, Y.J.; Lee, H.J.; Kim, K.S. Carbohydrate changes in Cymbidium ‘Red Fire’in response to night interruption. Sci. Hortic. 2013, 162, 82–89. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Ito, T.; Iseki, K.; Baba, C.; Imagawa, H.; Yagi, Y.; Morita, H.; Asakawa, Y.; Kawano, S.; Hashimoto, T. Phenanthrene derivatives from Cymbidium Great Flower Marie Laurencin and their biological activities. J. Nat. Prod. 2012, 75, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.M.; Yang, M.; Jin, Y.; Kim, E.M.; Ko, J.; Lee, J. Identification of major flavone C-glycosides and their optimized extraction from Cymbidium kanran using deep eutectic solvents. Molecules 2017, 22, 2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.S.; Fu, Y.; Chen, J.; Jiao, Y.; Chen, S.Q. Six phenanthrenes from the roots of Cymbidium faberi Rolfe. and their biological activities. Nat. Prod. Res. 2020, 1–12. [Google Scholar] [CrossRef]
- Won, J.H.; Kim, J.Y.; Yun, K.J.; Lee, J.H.; Back, N.I.; Chung, H.G.; Chung, S.A.; Jeong, T.S.; Choi, M.S.; Lee, K.T. Gigantol isolated from the whole plants of Cymbidium goeringii inhibits the LPS-induced iNOS and COX-2 expression via NF-kappaB inactivation in RAW 264.7 macrophages cells. Planta Med. 2006, 72, 1181–1187. [Google Scholar] [CrossRef]
- Rampilla, V.; Khasim, S.M. GC-MS analysis of organic extracts of Cymbidium aloifolium (L.) Sw. (Orchidaceae) leaves from Eastern Ghats of India. In Orchid Biology; Recent Trends & Challenges; Springer: Singapore, 2020; pp. 507–517. [Google Scholar]
- Chowdappa, S.; Jagannath, S.; Konappa, N.; Udayashankar, A.C.; Jogaiah, S. Detection and characterization of antibacterial siderophores secreted by endophytic fungi from Cymbidium aloifolium. Biomolecules 2020, 10, 1412. [Google Scholar] [CrossRef]
- Hinsley, A.; De Boer, H.J.; Fay, M.F.; Gale, S.W.; Gardiner, L.M.; Gunasekara, R.S.; Kumar, P.; Masters, S.; Metusala, D.; Roberts, L.R.; et al. A review of the trade in orchids and its implications for conservation. Bot. J. Linn. Soc. 2018, 186, 435–455. [Google Scholar] [CrossRef]
- Park, P.H.; Ramya, M.; An, H.R.; Park, P.M.; Lee, S.Y. Breeding of Cymbidium ‘Sale Bit’with bright yellow flowers and floral scent. Korean Soc. Breed. Sci. 2019, 51, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Kostenyuk, I.; Oh, B.J.; So, I.S. Induction of early flowering in Cymbidium niveo-marginatum Mak in vitro. Plant Cell Rep. 1999, 19, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Chen, G.; Ma, Y.P.; Wang, C.G.; Lin, B.; Yang, Y.Q.; Li, W.; Koike, K.; Hou, Y.; Li, N. Isolation, structural elucidation, optical resolution, and antineuroinflammatory activity of phenanthrene and 9,10-dihydrophenanthrene derivatives from Bletilla striata. J. Nat. Prod. 2019, 82, 2238–2245. [Google Scholar] [CrossRef] [PubMed]
- An, H.R.; Kim, Y.J.; Kim, K.S. Flower initiation and development in Cymbidium by night interruption with potassium and nitrogen. Hortic. Environ. Biotechnol. 2012, 53, 204–211. [Google Scholar] [CrossRef]
- Barman, D.; Bharathi, T.U.; Medhi, R.P. Effect of media and nutrition on growth and flowering of Cymbidium hybrid ‘HC Aurora’. Indian J. Hortic. 2012, 69, 395–398. [Google Scholar]
- Lee, N.; Lee, C.Z. Growth and flowering of Cymbidium ensifolium var. misericors as influenced by temperature. Acta Horticulturae 1991, 337, 123–130. [Google Scholar] [CrossRef]
- Yu, H.; Goh, C.J. Molecular Genetics of Reproductive Biology in Orchids. Plant Physiol. 2001, 127, 1390–1393. [Google Scholar] [CrossRef]
- Yang, W.K.; Li, T.Q.; Wu, S.M.; Finnegan, P.M.; Gao, J.Y. Ex situ seed baiting to isolate germination-enhancing fungi for assisted colonization in Paphiopedilum spicerianum, a critically endangered orchid in China. Glob. Ecol. Conserv. 2020, 23, e01147. [Google Scholar] [CrossRef]
- Li, X.; Jin, F.; Jin, L.; Jackson, A.; Ma, X.; Shu, X.; Wu, D.; Jin, G. Characterization and comparative profiling of the small RNA transcriptomes in two phases of flowering in Cymbidium ensifolium. BMC Genom. 2015, 16, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Sugiura, N. Specialized pollination by honeybees in Cymbidium dayanum, a fall–winter flowering orchid. Plant Species Biol. 2019, 34, 19–26. [Google Scholar] [CrossRef]
- Suetsugu, K. Autonomous self-pollination and insect visitors in partially and fully mycoheterotrophic species of Cymbidium (Orchidaceae). J. Plant Res. 2015, 128, 115–125. [Google Scholar] [CrossRef]
- Du Puy, D.; Cribb, P. The genus Cymbidium. In Surrey, Royal Botanic Gardens, 2nd ed.; Kew Publishing: London, UK, 2007. [Google Scholar]
- Davies, K.L.; Stpiczyńska, M.; Turner, M.P. A rudimentary labellar speculum in Cymbidium lowianum (Rchb. f.) Rchb. f. and Cymbidium devonianum Paxton (Orchidaceae). Ann. Bot. 2006, 97, 975–984. [Google Scholar] [CrossRef] [Green Version]
- Thummavongsa, T. Taxonomy, Reproductive Biology and Seed Germination of Habenaria rhodocheila Hance complex (Orchidaceae). Ph.D. Dissertation, School of Biology Institute of Science, Suranaree University of Technology, Nakhon Ratchasima, Thailand, 2021. [Google Scholar]
- Balilashaki, K.; Vahedi, M.; Ho, T.T.; Niu, S.C.; Cardoso, J.C.; Zotz, G.; Khodamzadeh, A.A. Biochemical, cellular and molecular aspects of Cymbidium orchids: An ecological and economic overview. Acta Physiol. Plant. 2022, 44, 24. [Google Scholar] [CrossRef]
- Kjellsson, G.; Rasmussen, F.N.; Dupuy, D. Pollination of Dendrobium infundibulum, Cymbidium insigne (Orchidaceae) and Rhododendron lyi (Ericaceae) by Bombus eximius (Apidae) in Thailand: A possible case of floral mimicry. J. Trop. Ecol. 1985, 1, 289–302. [Google Scholar] [CrossRef]
- Nanekar, V.; Shriram, V.; Kumar, V.; Kishor, P.K. Asymbiotic in vitro seed germination and seedling development of Eulophia nuda Lindl., an endangered medicinal orchid. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2014, 84, 837–846. [Google Scholar] [CrossRef]
- Puspitaningtyas, D.M.; Handini, E. Ex-situ conservation of Cymbidium finlaysonianum by seed storage. Biodiversitas J. Biol. Divers. 2020, 21. [Google Scholar] [CrossRef]
- Gantait, S.; Mitra, M. Applications of synthetic seed technology for propagation, storage, and conservation of orchid germplasms. In Synthetic Seeds: Germplasm Regeneration, Preservation and Prospects; Springer: Cham, Switzerland, 2019; pp. 301–321. [Google Scholar]
- Garg, R.; Maheshwari, S. Synthetic seed technology, application and future trends. EPH-Int. J. Agric. Environ. Res. 2023, 9, 1–10. [Google Scholar] [CrossRef]
- Patavardhan, S.S.; Ignatius, S.; Thiyam, R.; Lasrado, Q.; Karkala, S.; D’Souza, L.; Nivas, S.K. Asymbiotic seed germination and in vitro development of orchid Papilionanthe Miss Joaquim. Ornam. Hortic. 2022, 28, 246–255. [Google Scholar] [CrossRef]
- Seaton, P.; Kendon, J.P.; Pritchard, H.W.; Puspitaningtyas, D.M.; Marks, T.R. Orchid conservation: The next ten years. Lankesteriana 2013, 13, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Whigham, D.F.; O’Neill, J.P.; Rasmunssen, H.N.; Caldwell, B.A.; McCormick, M.K. Seed longevity in terrestrial orchids-Potential for persistent in-situ seed banks. Biol. Conserv. 2006, 129, 2–30. [Google Scholar] [CrossRef]
- Suzuki, R.M.; Moreira, V.C.; Pescador, R.; de Melo Ferreira, W. Asymbiotic seed germination and in vitro seedling development of the threatened orchid Hoffmannseggella cinnabarina. In vitro Cell. Dev. Biol. -Plant 2012, 48, 500–511. [Google Scholar] [CrossRef]
- Paudel, M.; Pradhan, S.; Pant, B. In vitro seed germination and seedling development of Esmeralda clarkei Rchb. f. (Orchidaceae). Plant Tissue Cult. Biotechnol. 2012, 22, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, J.C.; Zanello, C.A.; Chen, J.-T. An Overview of Orchid Protocorm-Like Bodies: Mass Propagation, Biotechnology, Molecular Aspects, and Breeding. Int. J. Mol. Sci. 2020, 21, 985. [Google Scholar] [CrossRef] [Green Version]
- Udomdee, W.; Wen, P.J.; Lee, C.Y.; Chin, S.W.; Chen, F.C. Effect of sucrose concentration and seed maturity on in vitro germination of Dendrobium nobile hybrids. Plant Growth Regul. 2014, 72, 249–255. [Google Scholar] [CrossRef]
- Hossain, M.M.; Dey, R. Multiple regeneration pathways in Spathoglottis plicata Blume—A study in vitro. S. Afr. J. Bot. 2013, 85, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Vudala, S.M.; Ribas, L.L.F. Seed storage and asymbiotic germination of Hadrolaelia grandis (Orchidaceae). S. Afr. J. Bot. 2017, 108, 1–7. [Google Scholar] [CrossRef]
- Huh, Y.S.; Lee, J.K.; Nam, S.Y.; Hong, E.Y.; Paek, K.Y.; Son, S.W. Effects of altering medium strength and sucrose concentration on in vitro germination and seedling growth of Cypripedium macranthos Sw. J. Plant Biotechnol. 2016, 43, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S.; Zhang, Y.; Teixeira da Silva, J.A.; Wu, K.; Zhang, J.; Duan, J. Seed biology and in vitro seed germination of Cypripedium. Crit. Rev. Biotechnol. 2014, 34, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Yan, S.; Zhang, Y. Factors affecting germination and propagators of artificial seeds of Dendrobium candidum. In Proceedings of the International Conference on Agricultural and Biosystems Engineering, Amsterdam, Netherlands, 13–15 July 2011; pp. 1–2. [Google Scholar]
- Lu, Y.X.; Li, C.J.; Zhang, F.S. Transpiration, potassium uptake and flow in tobacco as affected by nitrogen forms and nutrient levels. Ann. Bot. 2005, 95, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Xiang, M.A.; Zheng, F.W.; Li, Y.; Liu, L.; Wu, J. Symbiotic seed germination and seedling growth promoted by Rhizoctonia fungi in Cymbidium mastersii, an endangered orchid species endemic to Southwest of China. In Proceedings of the 18th EOCCE-What Future for Orchids, Paris, France, 24 March 2018. [Google Scholar]
- Teixeira da Silva, J.A.; Norikane, A.; Tanaka, M. Cymbidium: Successful in vitro growth and subsequent acclimatization. Acta Hort. 2015, 748, 207–214. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, H.J.; Kim, K.S. Night interruption promotes vegetative growth and flowering of Cymbidium. Sci. Hortic. 2011, 130, 887–893. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Yam, T.; Fukai, S.; Nayak, N.; Tanaka, M. Establishment of optimum nutrient media for in vitro propagation of Cymbidium Sw. (Orchidaceae) using protocorm-like body segments. Propag. Ornam. Plants 2016, 5, 129–136. [Google Scholar]
- Chand, K.; Shah, S.; Sharma, J.; Paudel, M.R.; Pant, B. Isolation, characterization, and plant growth-promoting activities of endophytic fungi from a wild orchid Vanda cristata. Plant Signal. Behav. 2020, 15, 174–294. [Google Scholar] [CrossRef] [PubMed]
- Acemi, A.; Özen, F. Optimization of in vitro asymbiotic seed germination protocol for Serapias vomeracea. EuroBiotech J. 2019, 3, 143–151. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Rasmussen, F.N. Seedling mycorrhiza: A discussion of origin and evolution in Orchidaceae. Bot. J. Linn. Soc. 2014, 175, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Yam, T.W.; Arditti, J. History of orchid propagation: A mirror of the history of biotechnology. Plant Biotechnol. Rep. 2009, 3, 1–56. [Google Scholar] [CrossRef] [Green Version]
- Valadares, R.B.S.; Perotto, S.; Santos, E.C.; Lambais, M.R. Proteome changes in Oncidium sphacelatum (Orchidaceae) at different trophic stages of symbiotic germination. Mycorrhiza 2014, 24, 349–360. [Google Scholar] [CrossRef]
- Rafter, M.; Yokoya, K.; Schofield, E.J.; Zettler, L.W.; Sarasan, V. Non-specific symbiotic germination of Cynorkis purpurea (Thouars) Kraezl a habitat-specific terrestrial orchid from the Central Highlands of Madagascar. Mycorrhiza 2016, 26, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Nikabadi, S.; Bunn, E.; Stevens, J.; Newman, B.; Turner, S.R.; Dixon, K.W. Germination responses of four native terrestrial orchids from south-west Western Australia to temperature and light treatments. Plant Cell Tissue Organ Cult. 2014, 118, 559–569. [Google Scholar] [CrossRef]
- Mala, B.; Kuegkong, K.; Sa-Ngiaemsri, N.; Nontachaiyapoom, S. Effect of germination media on in vitro symbiotic seed germination of three Dendrobium orchids. S. Afr. J. Bot. 2017, 112, 521–526. [Google Scholar] [CrossRef]
- Yang, Q.; Xu, L.; Xia, W.; Liang, L.; Bai, X.; Li, L.; Xu, L.; Liu, L. Mycorrhizal compatibility and germination-Promoting activity of Tulasnella species in two species of orchid (Cymbidium mannii and Epidendrum radicans). Horticulturae 2021, 7, 472. [Google Scholar] [CrossRef]
- Shao, S.C.; Burgess, K.S.; Cruse-Sanders, J.M.; Liu, Q.; Fan, X.L.; Huang, H.; Gao, J.Y. Using in situ symbiotic seed germination to restore over-collected medicinal orchids in Southwest China. Front. Plant Sci. 2017, 8, 888. [Google Scholar] [CrossRef] [Green Version]
- Smith, Z.F.; James, E.A.; Mclean, C.B. Experimental reintroduction of the threatened terrestrial orchid Diuris fragrantissima. Lankesteriana Int. J. Orchid. 2007, 7, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, F.; Liu, H.X.; Jin, H.; Luo, Y.B. Symbiosis between fungi and the hybrid Cymbidium and its mycorrhizal microstructures. For. Stud. China 2008, 10, 41–44. [Google Scholar] [CrossRef]
- Liu, S.; Liu, M.; Liao, Q.G.; Lü, F.B.; Zhao, X.L. Effects of inoculated mycorrhizal fungi and non-mycorrhizal beneficial microorganisms on plant traits, nutrient uptake and root associated fungal community composition of the Cymbidium hybrid in greenhouse. J. Appl. Microbiol. 2021, 131, 413–424. [Google Scholar] [CrossRef]
- Gogoi, K.; Kumaria, S.; Tandon, P. Ex situ conservation of Cymbidium eburneum Lindl.: A threatened and vulnerable orchid, by asymbiotic seed germination. 3 Biotech. 2012, 2, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Herrera, H.; Valadares, R.; Contreras, D.; Bashan, Y.; Arriagada, C. Mycorrhizal compatibility and symbiotic seed germination of orchids from the Coastal Range and Andes in south central Chile. Mycorrhiza 2017, 27, 175–188. [Google Scholar] [CrossRef]
- Zi, X.M.; Sheng, C.L.; Goodale, U.M.; Shao, S.C.; Gao, J.Y. In situ seed baiting to isolate germination-enhancing fungi for an epiphytic orchid, Dendrobium aphyllum (Orchidaceae). Mycorrhiza 2014, 24, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Sheng, C.L.; Lee, Y.; Gao, J.Y. Ex situ symbiotic seed germination, isolation and identification of effective symbiotic fungus in Cymbidium mannii (Orchidaceae). Chin. J. Plant Ecol. 2012, 36, 859. [Google Scholar] [CrossRef]
- Prutsch, J.; Schardt, A.; Schill, R. Adaptations of an orchid seed to water uptake and-storage. Plant Syst. Evol. 2000, 220, 69–75. [Google Scholar] [CrossRef]
- Chang, C.; Shiu, Q.J. January. Pollination, Seed Development and In vitro Germination of Cymbidium sinense. I Int. Orchid Symp. 2010, 878, 251–259. [Google Scholar]
- Tandon, P.; Kumaria, S. Prospects of plant conservation biotechnology in India with special reference to Northeastern region. In Biodiversity: Status and Prospects; Tandon, P., Sharma, M., Swarup, R., Eds.; Narosa Publshing House: New Delhi, India, 2005; pp. 79–91. [Google Scholar]
- Mohanty, P.; Paul, S.; Das, M.C.; Kumaria, S.; Tandon, P. A simple and efficient protocol for the mass propagation of Cymbidium mastersii: An ornamental orchid of Northeast India. AoB Plants 2012, 2012, pls023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deb, C.R.; Pongener, A. Asymbiotic seed germination and in vitro seedling development of Cymbidium aloifolium (L.) Sw: A multipurpose orchid. J. Plant Biochem. Biotechnol. 2011, 20, 90–95. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dohling, S.; Kumaria, S.; Tandon, P. Optimization of nutrient requirements for asymbiotic seed germination of D. longicornu and D. formosumi Roxb. Proc. Indian Natl. Sci. Acad. 2008, 74, 167–171. [Google Scholar]
- Utami, E.S.W.; Hariyanto, S. Organic compounds: Contents and their role in improving seed germination and protocorm development in orchids. Int. J. Agron. 2020, 2020, 2795108. [Google Scholar] [CrossRef]
- Dutra, D.; Johnson, T.R.; Kauth, P.J.; Stewart, S.L.; Kane, M.E.; Richardson, L. Asymbiotic seed germination, in vitro seedling development, and greenhouse acclimatization of the threatened terrestrial orchid Bletia purpurea. Plant Cell Tissue Organ Cult. 2008, 94, 11–21. [Google Scholar] [CrossRef]
- Hajong, S.; Kumaria, S.; Tandon, P. In vitro propagation of medicinal orchid Dendrobium chrysanthum. Proc. Indian Natl. Sci. Acad. 2010, 76, 67–70. [Google Scholar]
- Bhowmik, T.K.; Rahman, M.M. Effect of different basal media and PGRs on in vitro seed germination and seedling development of medicinally important orchid Cymbidium aloifolium (L.) Sw. J. Pharmacogn. Phytochem. 2017, 6, 167–172. [Google Scholar]
- Bhattacharjee, D.K.; Hossain, M.M. Effect of plant growth regulators and explants on propagation on a monopodial and sympodial orchid: A study in vitro. J. Orchid Soc. India 2015, 29, 91–102. [Google Scholar]
- Teixeira da Silva, J.A.; Tanaka, M. Multiple regeneration pathways via Thin Cell Layers in hybrid Cymbidium (Orchidaceae). J. Plant Growth Regul. 2006, 25, 203. [Google Scholar] [CrossRef]
- Morel, G.M. A new means of clonal propagation of orchids. Am. Orchid Soc. Bull. 1964, 31, 437–477. [Google Scholar]
- Chang, C.; Chang, W.C. Micropropagation of Cymbidium ensifolium var. Misericors through Callus-Derived Rhizomes. In vitro Cellular & Developmental Biology. Plant 2000, 36, 517–520. [Google Scholar]
- Wimber, D.E. Clonal multiplication of Cymbidiums through tissue culture of the shoot meristem. Am. Orchid Soc. Bull. 1963, 32, 105–107. [Google Scholar]
- Malabadi, R.B.; Teixeira da Silva, J.A.; Nataraja, K.; Mulgund, G.S. Shoot tip transverse thin cell layers and 2,4-epibrassinolide in the micropropagation of Cymbidium bicolor Lindl. Floric. Ornam. Biotech. 2008, 2, 44–48. [Google Scholar]
- Begum, A.A.; Tamaki, M.; Kako, S. Formation of protocorm-like bodies (PLBs) and shoot development through in vitro culture of outer tissue of Cymbidium PLB. J. Jpn. Soc. Hort. Sci. 1994, 63, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.M.; Sharma, M.; Pathak, P. In vitro propagation of Dendrobium aphyllum (Orchidaceae)-seed germination to flowering. J. Plant Biochem. Biotechnol. 2013, 22, 157–167. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Singh, N.; Tanaka, M. Priming biotic factors for optimal protocorm-like body and callus induction in hybrid Cymbidium (Orchidaceae), and assessment of cytogenetic stability in regenerated plantlets. Plant Cell Tissue Organ Cult. 2006, 84, 135–144. [Google Scholar] [CrossRef]
- Shimasaki, K.; Uemoto, S. Micropropagation of a terrestrial Cymbidium species using rhizomes developed from seeds and pseudobulbs. Plant Cell Tissue Organ Cult. 1990, 22, 237–244. [Google Scholar] [CrossRef]
- Chugh, S.; Guha, S.; Rao, I.U. Micropropagation of orchids: A review on the potential of different explants. Sci. Hortic. 2009, 122, 507–520. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sharma, M.; Pathak, P. In vitro mass propagation of an economically important orchid, Cymbidium aloifolium (L.) Sw. J. Orchid Soc. 2008, 22, 91–95. [Google Scholar]
- Yasugi, S.; Sakamoto, K.; Onodera, K.; Tamashiro, M. Plantlet regeneration in root segment culture of Cymbidium Kenny ‘Wine Color’. Plant Tissue Cult. Lett. 1994, 11, 150–152. [Google Scholar] [CrossRef]
- Begum, A.A.; Tamaki, M.; Tahara, M.; Kako, S. Somatic embryogenesis in Cymbidium through in vitro culture of inner tissue of protocorm-like bodies. J. Jpn. Soc. Hort. Sci. 1994, 63, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Huan, L.V.T.; Takamura, T.; Tanaka, M. Callus formation and plant regeneration from callus through somatic embryo structures in Cymbidium orchid. Plant Sci. 2004, 166, 1443–1449. [Google Scholar] [CrossRef]
- Das, M.C.; Kumeria, S.; Tandon, P. Protocorm regeneration, multiple shoot induction and ex vitro establishment of Cymbidium devonianum Paxt. Asian J. Plant Sci. 2007, 6, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, S.; Tiruwa, B.; Subedee, B.R.; Pant, B. In vitro germination and propagation of a threatened medicinal orchid, Cymbidium aloifolium (L.) Sw. through artificial seed. Asian Pac. J. Trop. Biomed. 2014, 4, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Rihan, H.Z.; Kareem, F.; El-Mahrouk, M.E.; Fuller, M.P. Artificial seeds (principle, aspects and applications). Agronomy 2017, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.A.T. Production of synseed for hybrid Cymbidium using protocorm-like bodies. J. Fruit Ornam. Plant Res. 2012, 20, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Deb, C.R.; Pongener, A. Studies on the in vitro regenerative competence of aerial roots of two horticultural important Cymbidium species. J. Plant Biochem. Biotechnol. 2012, 21, 235–241. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sharma, M.; Pathak, P. Cost effective protocol for in vitro mass propagation of Cymbidium aloifolium (L.) Sw.—A medicinally important orchid. Eng. Life Sci. 2009, 9, 444–453. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sharma, M.; da Silva, J.A.T.; Pathak, P. Seed germination and tissue culture of Cymbidium giganteum Wall. ex Lindl. Sci. Hortic. 2010, 123, 479–487. [Google Scholar] [CrossRef]
- Nahar, S.J.; Shimasaki, K.; Haque, S.M. Effect of different light and two polysaccharides on the proliferation of protocorm-like bodies of Cymbidium cultured in vitro. Acta Hortic. 2012, 307–314. [Google Scholar] [CrossRef]
- Parmar, G.; Pant, B. In vitro seed germination and seedling development of the orchid Coelogyne stricta (D. Don) Schltr. Afr. J. Biotechnol. 2016, 15, 105–109. [Google Scholar]
- Pradhan, S.; Tiruwa, B.L.; Subedee, B.R.; Pant, B. Efficient plant regeneration of Cymbidium aloifolium (L.) Sw., a threatened orchid of Nepal through artificial seed technology. Am. J. Plant Sci. 2016, 7, 1964–1974. [Google Scholar] [CrossRef] [Green Version]
- Philip Robinson, J.; Jyoti, P.K.; Sebastinraj, J.; Suriya, K. In vitro seed germination of Cymbidium aloifolium (L.) Sw., a potential medicinal orchid from Eastern Ghats of Tamil Nadu, India. J. Plant Biotechnol. 2017, 44, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.; Islam, T.; Sarker, R.H.; Hoque, M.I. In vitro mass propagation of Cymbidium aloifolium (L.) Sw. Plant Tissue Cult. Biotechnol. 2019, 29, 73–79. [Google Scholar] [CrossRef]



{kind=link}
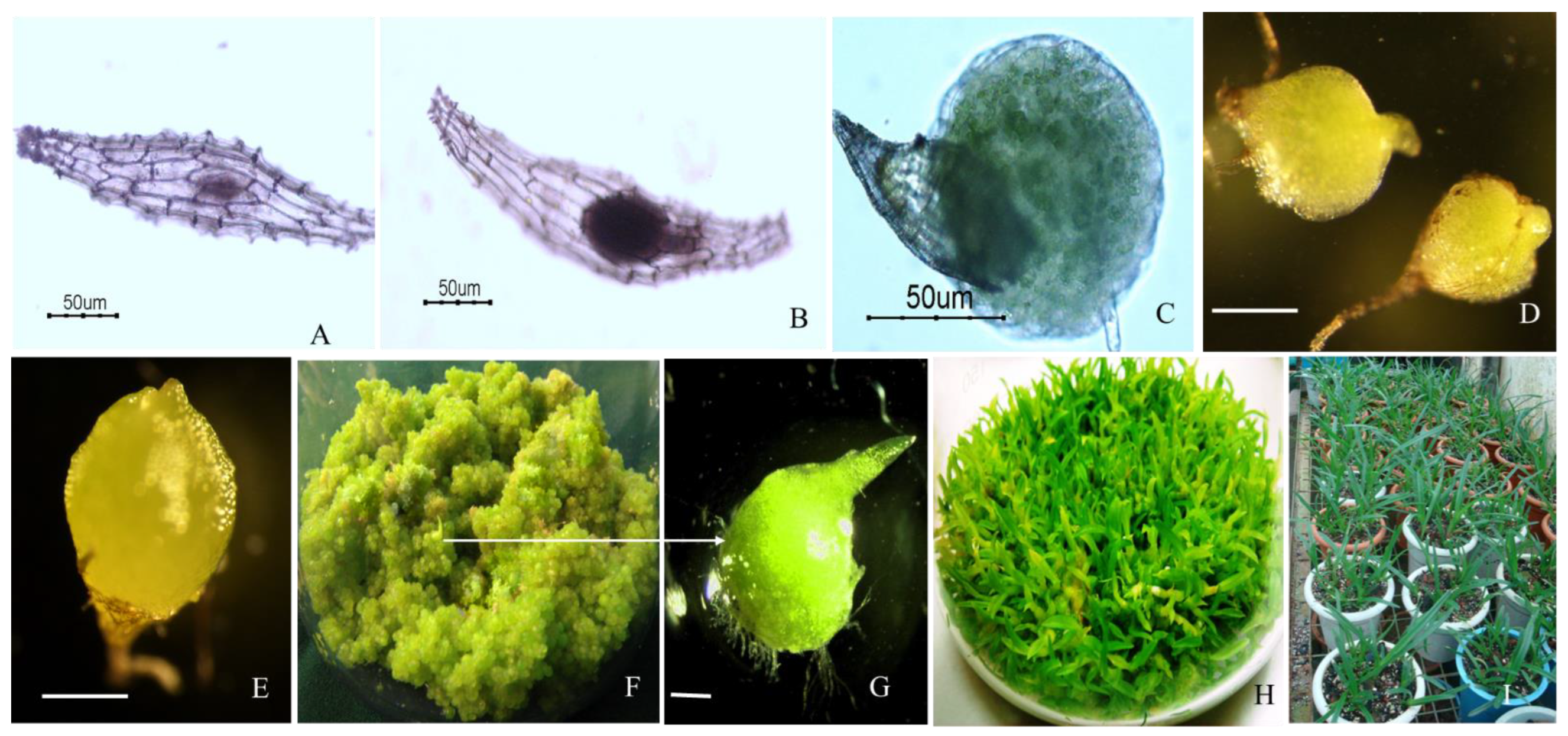{kind=link}
| Researchers | Year | Subject | Results |
|---|---|---|---|
| Deb and Pongener [104] | 2002 | Studies on the in vitro regenerative competence of aerial roots of two horticulturally important Cymbidium species | Of the three basal media tested, MS medium supported optimum regeneration and culture proliferation in both the species. In C. aloifolium ~12, shoot buds developed on medium nourished with sucrose 3% and benzyl adenine (BA) 3 μM but in C. iridioides optimum regeneration was achieved when medium supplemented with sucrose 3%, coconut water (CW) 15%, casein hydrolysate (CH) 100 mg/L and ~20 shoot buds formed per subculture |
| Hossain et al. [105] | 2009 | Cost-effective protocol for in vitro mass propagation of C. aloifolium (L.) Sw.–a medicinally important orchid | Mitra medium supplemented with 2.0 g/L activated charcoal (AC) showed 100% seed germination and effective for induction of significantly large-size protocorms (1.64 mm in dia.) |
| Hossain et al. [106] | 2010 | Seed germination and tissue culture of C. giganteum Wall. ex Lindl | The effects of peptone, AC and two-plant growth regulators [6-benzylaminopurine (BAP) and 2,4-Dichlorophenoxyacetic acid (2,4-D)] were also studied. Both M and PM supplemented with 2.0 g/L peptone or 1.0 mg/L BAP resulted in ∼100% seed germination. |
| Deb and Pongener [78] | 2011 | Asymbiotic seed germination and in vitro seedling development of C. aloifolium (L.) Sw.: a multipurpose orchid | Immature embryos of 9 months after pollination were successfully germinated on MS medium containing sucrose (2%) (w/v) and α-naphthalene acetic acid (NAA) and BA (3 and 6 μM, respectively, in combination) within 45 days of culture where 90% germination was recorded. The germinated seeds formed PLBs on the optimum germination medium within two passages. |
| Nahar et al. [107] | 2012 | Effect of different light and two polysaccharides on the proliferation of protocorm-like bodies of Cymbidiums cultured in vitro | The highest protocorm-like bodies (PLBs) formation, shoot formation rate (90%) and root formation rate (50%) were found among explants cultured on medium supplemented with 0.1 mg/L Chitosan H under green light. After 11 weeks of culture, fresh weight of PLBs was higher (241.3 mg) at HA9 (1 mg/L) treatment with green light. The average number of PLBs (5.7) was higher under green light at HA9 treatment. PLBs under white light showed the highest number of shoot (1.2) at Chitosan H treatment. |
| Parmar and Pant [108] | 2016 | In vitro seed germination and seedling development of the orchid Coelogyne stricta (D. Don) Schltr | MS medium supplemented with 1 mg/L BAP and 1 mg/L NAA was found to be the best condition for the development. The germination started after 7 weeks of culture and complete seedlings were obtained after 23 weeks of culture on the medium supplemented with 1 mg/L BAP and 1 mg/L NAA suggesting the usefulness of both hormones in root induction. In the hormone, free MS medium germination started after 5 weeks, but root initials were not developed even after 32 weeks of culture. |
| Pradhan et al. [109] | 2016 | Efficient plant regeneration of C. aloifolium (L.) Sw., a threatened orchid of Nepal through artificial seed technology | Full strength of MS medium without plant growth regulators was found to be the most favourable condition for efficient plantlet regeneration of C. aloifolium (9.83 shoot and 2.66 roots per culture). |
| Bhowmik and Rahman [84] | 2017 | Effect of different basal media and PGRs on in vitro seed germination and seedling development of medicinally important orchid C. aloifolium (L.) | Medium supplemented with hormones favored optimum condition for the germination (approx. 95%) of seeds followed by full strength and half strength on KC, MS, PM and VW media. MS medium supplemented with 0.5 mg/L BAP and 0.5 mg/L NAA showed comparatively better response within 6 weeks of culture than other conditions of MS medium as well as KC, PM and VW media. |
| Philip Robinson et al. [110] | 2017 | In vitro seed germination of C. aloifolium (L.) Sw., a potential medicinal orchid from Eastern Ghats of Tamil Nadu, India | The highest seed germination of 90% was observed KC basal media after30th days whereas germination percentages were 40% and 30% on 1/2 MS and VW media respectively. |
| Paul et al. [111] | 2019 | In vitro mass propagation of C. aloifolium (L.) Sw | Developing an efficient protocol for rapid propagation of C. aloifolium starting with in vitro asymbiotic seed germination, leading to protocorm induction followed by plantlet development and successful ex vitro acclimation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balilashaki, K.; Martinez-Montero, M.E.; Vahedi, M.; Cardoso, J.C.; Silva Agurto, C.L.; Leiva-Mora, M.; Feizi, F.; Musharof Hossain, M. Medicinal Use, Flower Trade, Preservation and Mass Propagation Techniques of Cymbidium Orchids—An Overview. Horticulturae 2023, 9, 690. https://doi.org/10.3390/horticulturae9060690
Balilashaki K, Martinez-Montero ME, Vahedi M, Cardoso JC, Silva Agurto CL, Leiva-Mora M, Feizi F, Musharof Hossain M. Medicinal Use, Flower Trade, Preservation and Mass Propagation Techniques of Cymbidium Orchids—An Overview. Horticulturae. 2023; 9(6):690. https://doi.org/10.3390/horticulturae9060690
Chicago/Turabian StyleBalilashaki, Khosro, Marcos Edel Martinez-Montero, Maryam Vahedi, Jean Carlos Cardoso, Catherine Lizzeth Silva Agurto, Michel Leiva-Mora, Fatemeh Feizi, and Mohammad Musharof Hossain. 2023. "Medicinal Use, Flower Trade, Preservation and Mass Propagation Techniques of Cymbidium Orchids—An Overview" Horticulturae 9, no. 6: 690. https://doi.org/10.3390/horticulturae9060690
APA StyleBalilashaki, K., Martinez-Montero, M. E., Vahedi, M., Cardoso, J. C., Silva Agurto, C. L., Leiva-Mora, M., Feizi, F., & Musharof Hossain, M. (2023). Medicinal Use, Flower Trade, Preservation and Mass Propagation Techniques of Cymbidium Orchids—An Overview. Horticulturae, 9(6), 690. https://doi.org/10.3390/horticulturae9060690