
Fine Mapping and Candidate Gene Analysis of the Gv1 Locus Controlling Green-Peel Color in Eggplant (Solanum melongena L.)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
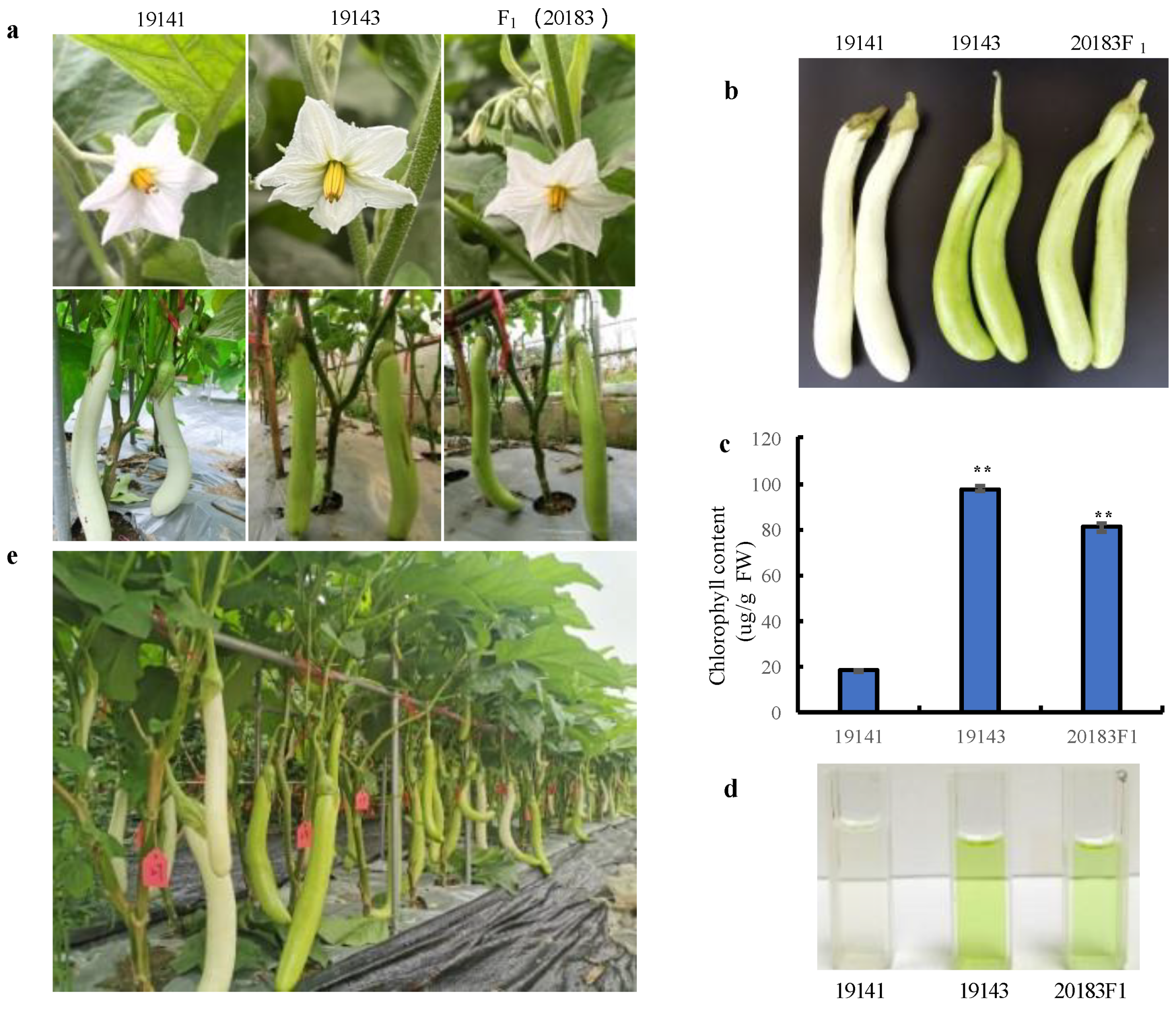
3.1. Inheritance of Peel Color in Eggplant Controlled by Gv1
3.2. Identification of the Candidate Region Associated with Gv1 by Whole Genome Re-Sequencing
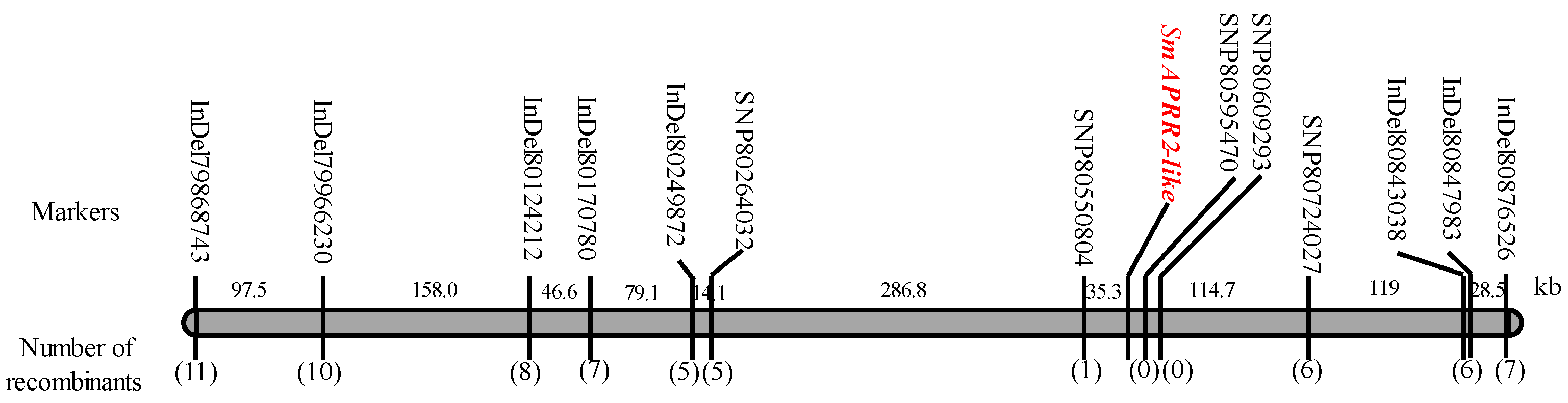
3.3. Fine Mapping of Gv1 with InDel and SNP Markers
- Annotation and screening of candidate genes
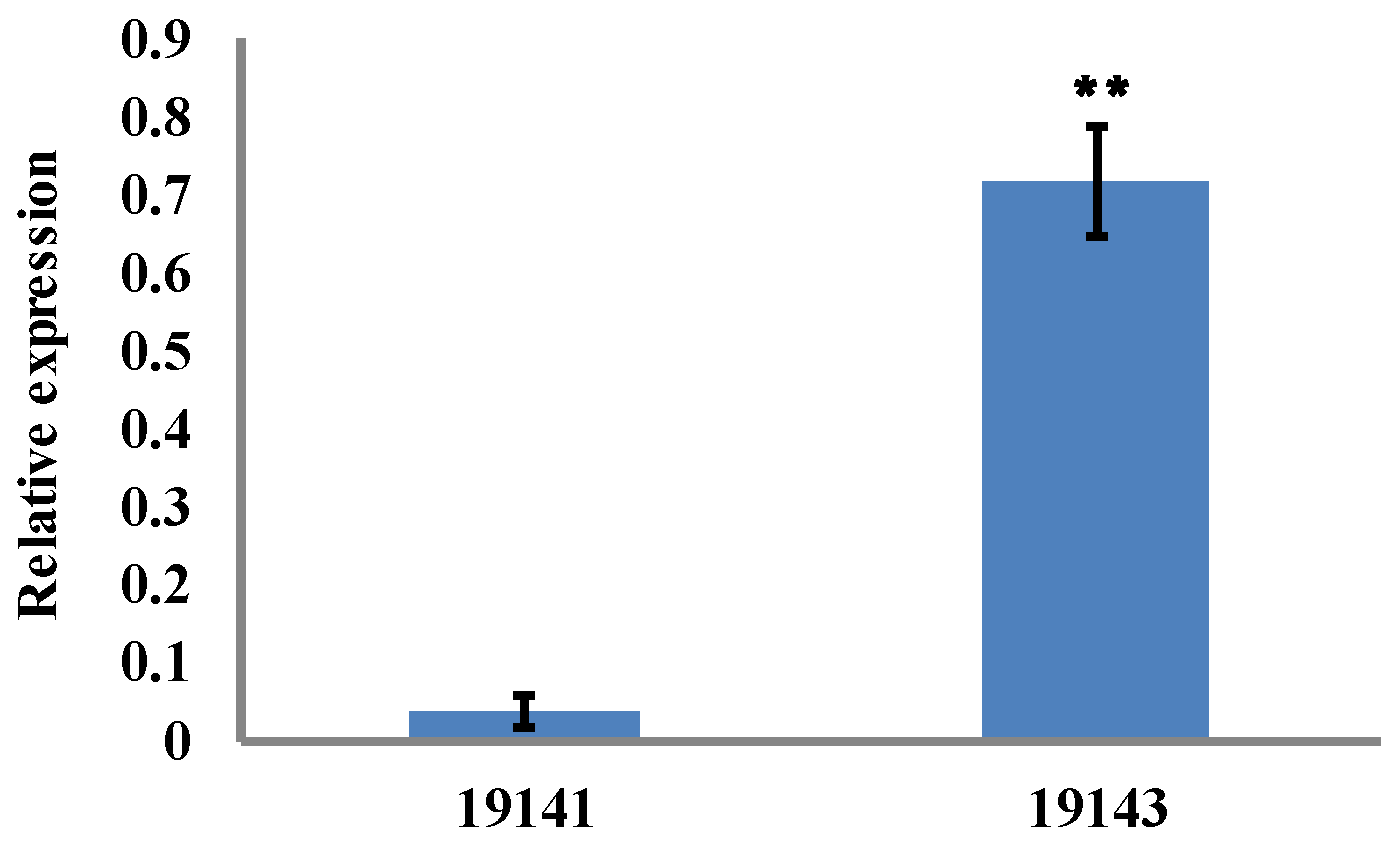
3.4. Cloning of SmAPRR2-like
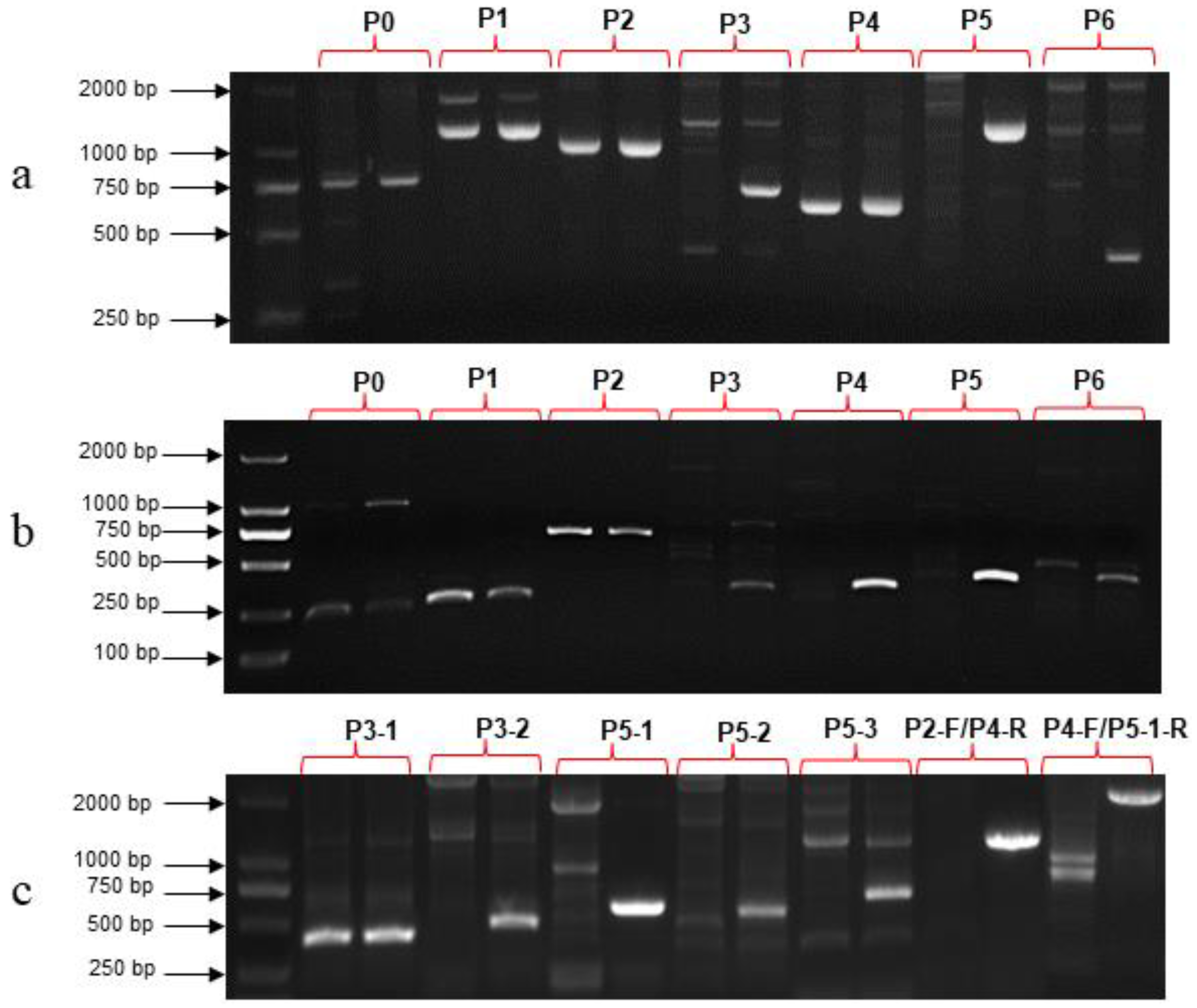
3.5. Development of Molecular Marker Based on the SmAPRR2-like Gene
3.6. Detection of Genetic Variation in SmAPRR2-like in Natural Population
3.7. Mining other Genetic Variations of SmAPRR2-like
4. Discussion
4.1. Mapping of Genes Controlling Green Fruit Color in Edible Fruit and Vegetables
4.2. APRR2-like Gene Related to Fruit Color
4.3. Development and Utilization of Molecular Markers for Eggplant Peel Color
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Daunay, M.C.; Aubert, S.; Frary, A.; Doganlar, S.; Lester, R.N.; Barendse, G.; van der Weerden, G.; Hennart, J.W.; Haanstra, J.; Dauphin, F.; et al. Eggplant (Solanum melongena L.) fruit color: Pigments, measurements and genetics. In Proceedings of the 12 Eucarpia Meeting on Genetics and Breeding of Capsicum and Eggplant, Noordwijkerhout, The Netherlands, 17–19 May 2004; pp. 108–116. [Google Scholar]
- Pan, Y.; Bradley, G.; Pyke, K.; Ball, G.; Lu, C.; Fray, R.; Marshall, A.; Jayasuta, S.; Baxter, C.; van Wijk, R.; et al. Network inference analysis identifies an APRR2-like gene linked to pigment accumulation in tomato and pepper fruits. Plant Physiol. 2013, 161, 1476–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.Y.; Miao, H.; Zhang, S.P.; Liu, M.M.; Wang, Y.; Gu, X.F. Genetic analysis and gene mapping of white fruit skin in cucumber (Cucumis stativus L.). Acta Bota Borea Occident. Sin. 2012, 32, 2177–2181. (In Chinese) [Google Scholar]
- Dou, W.W.; Shen, D.; Dong, H.X.; Qiu, Y.; Li, X.X. QTL mapping of immature fruit basal color in Cucumis sativus L. J. Plant Genet. Resour. 2018, 19, 1138–1142. (In Chinese) [Google Scholar] [CrossRef]
- Hu, K.L.; Fu, Q.M.; Wang, G.P.; Hu, Z.Q. Study on the heredity of fruit colour of Momordica charantia. China Veg. 2002, 6, 11–12. (In Chinese) [Google Scholar]
- Deng, J.Y.; Wang, Z.L.; Wu, P.; Liu, C.A.; Deng, J.L.; Li, L.Z.; Zhou, Y.X. Study on the inheritance of fruit color in wax guard. J. Anhui Agr. Sci. 2015, 43, 40–41. (In Chinese) [Google Scholar]
- Behera, T.K.; Satyavati, C.T.; Pal, A. Generation mean analysis of yield related traits and inheritance of fruit colour and surface in bitter gourd. Indian J. Hortic. 2012, 69, 65–69. [Google Scholar]
- Sun, X.D.; Shang, Q.M.; Qin, Z.W. Genetic regularity of white skin color and its AFLP markers in cucumber tender fruit. North. Hortic. 2011, 3, 135–140. [Google Scholar]
- Liu, H.Q.; Jiao, J.Q.; Liang, X.J.; Liu, J.; Meng, H.W.; Chen, S.X.; Li, Y.H.; Cheng, Z.H. Map-based cloning, identification and characterization of the w gene controlling white immature fruit color in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2016, 129, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Tatebe, T. On inheritance of color in Solanum melongena L. Jpn. J. Genet. 1939, 15, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Sambandam, C.N. Guide chart for color combinations in hybrid eggplants. Econ. Bot. 1967, 21, 309–311. [Google Scholar] [CrossRef]
- Doganlar, S.; Frary, A.; Daunay, M.C.; Leste, R.N.; Tanksley, S.D. Conservation of gene function in the solanaceae as revealed by comparative mapping of domestication traits in eggplant. Genetics 2002, 161, 1713–1726. [Google Scholar] [CrossRef]
- Sun, B.J.; Wang, R.; Sun, G.W.; Wang, Y.K.; Li, T.; Gong, C.; Heng, Z.; You, Q.; Li, Z.L. Transcriptome and metabolome integrated analysis of epistatic genetics effects on eggplant peel color. Sci. Agric. Sin. 2022, 55, 3997–4010. (In Chinese) [Google Scholar]
- Wang, P.J.; Gu, M.Y.; Shao, S.X.; Chen, X.M.; Hou, B.H.; Ye, N.X.; Zhang, X.T. Changes in Non-Volatile and Volatile Metabolites Associated with Heterosis in Tea Plants (Camellia sinensis). J. Agric. Food Chem. 2022, 70, 3067–3078. [Google Scholar] [CrossRef]
- Wei, Q.Z.; Wang, J.L.; Wang, W.H.; Hu, T.H.; Hu, H.J.; Bao, C.L. A high-quality chromosome-level genome assembly reveals genetics for important traits in eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Mckenna, A.; Hanna, M.E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; Depristo, M.A. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Dhillon, S.; Ke, Y.X.; Collins, A.R.; Day, I.N.M. An efficient procedure for genotyping single nucleotide polymorphisms. Nucleic Acids Res. 2001, 29, e88. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Qian, J.; Li, W.L.; Yu, N.; Gan, G.Y.; Jiang, Y.Q.; Li, W.J.; Liang, X.Y.; Chen, R.Y.; Mo, Y.C.; et al. A high-quality genome assembly of the eggplant provides insights into the molecular basis of disease resistance and chlorogenic acid synthesis. Mol. Ecol. Resour. 2021, 21, 1274–1286. [Google Scholar] [CrossRef]
- Barchi, L.; Rabanus-Wallace, M.T.; Prohens, J.; Toppino, L.; Padmarasu, S.; Portis, E.; Rotino, G.L.; Stein, N.; Lanteri, S.; Giuliano, G. Improved genome assembly and pan-genome provide key insights into eggplant domestication and breeding. Plant J. 2021, 107, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.L.; Nguyen, C.V.; Hill, T.; Cheng, K.L.; Figueroa-Balderas, R.; Aktas, H.; Ashrafi, H.; Pons, C.; Fernández-Muñoz, R.; Vicente, A.; et al. Uniform ripening encodes a Golden 2-like transcription factor regulating tomato fruit chloroplast development. Science 2012, 336, 1711–1715. [Google Scholar] [CrossRef] [Green Version]
- Jia, T.; Cheng, Y.T.; Khan, I.; Zhao, X.; Gu, T.Y.; Hu, X.Y. Progress on understanding transcriptional regulation of chloroplast development in fleshy fruit. Int. J. Mol. Sci. 2020, 21, 6951. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Pang, X.Q.; Liu, W.J.; Wang, R.; Su, D.D.; Gao, Y.S.; Wu, M.B.; Deng, W.; Liu, Y.D.; Li, Z.G. SlZHD17 is involved in the control of chlorophyll and carotenoid metabolism in tomato fruit. Hortic. Res. 2021, 8, 259. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.; Borovsky, Y.; Meir, S.; Rogachev, I.; Aharoni, A.; Paran, I. pc8.1, a major QTL for pigment content in pepper fruit, is associated with variation in plastid compartment size. Planta 2012, 235, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.B.; Jiang, S.J.; Kang, M.Y.; Kim, S.; Kwon, J.K.; Kang, B.C. Candidate gene analysis reveals that the fruit color locus C1 corresponds to PRR2 in pepper (Capsicum frutescens). Front. Plant Sci. 2020, 11, 399. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Song, M.F.; Wei, Q.Z.; Wang, J.; Chen, J.F.; Lou, Q.F. Mapping and prediction of candidate gene w controlling white-green fruit color in cucumber. J. Nanjiang Agric. Univ. 2018, 41, 1003–1008. (In Chinese) [Google Scholar]
- Arrones, A.; Mangino, G.; Alonso, D.; Plazas, M.; Prohens, J.; Portis, E.; Barchi, L.; Giuliano, G.; Vilanova, S.; Gramazio, P. Mutations in the SmAPRR2 transcription factor suppressing chlorophyll pigmentation in the eggplant fruit peel are key drivers of a diversified colour palette. Front. Plant Sci. 2022, 13, 1025951. [Google Scholar] [CrossRef]
- Fang, H.; Wang, P.; Wang, W.; Peng, J.; Zheng, J.; Zhu, G.; Zhong, C.; Yu, W. Fine mapping and identification of SmAPRR2 regulating rind color in eggplant (Solanum melongena L.). Int. J. Mol. Sci. 2023, 24, 3059. [Google Scholar] [CrossRef]
- Hwang, I.; Chen, H.C.; Sheen, J. Two-component signal transduction pathways in Arabidopsis. Plant Physiol. 2002, 129, 500–515. [Google Scholar] [CrossRef] [Green Version]
- Lohrmann, J.; Harter, K. Plant two-component signaling systems and the role of response regulators. Plant Physiol. 2002, 128, 363–369. [Google Scholar] [CrossRef]
- Xu, X.Y.; Shen, J.; Zhang, Y.J.; Li, J.G.; Niu, X.W.; Shou, W.G. Fine mapping of an immature rind color gene GR in melon. Sci. Acricultura Sin. 2021, 54, 3308–3319. [Google Scholar]
- Borovsky, Y.; Monsonego, N.; Mohan, V.; Shabtai, S.; Kamara, I.; Faigenboim, A.; Hill, T.; Chen, S.; Stoffel, K.; Van Deynze, A.; et al. The zinc-finger transcription factor CcLOL1 controls chloroplast development and immature pepper fruit color in Capsicum chinense and its function is conserved in tomato. Plant J. 2019, 99, 41–55. [Google Scholar] [CrossRef]
- Tigchelaar, E.C.; Janick, J.; Erickson, H.T. The genetics of anthocyanin coloration in eggplant (Solanum melongena L.). Genetics 1968, 60, 475–491. [Google Scholar] [CrossRef]
- You, Q.; Li, H.M.; Wu, J.; Li, T.; Wang, Y.K.; Sun, G.W.; Li, Z.L.; Sun, B.J. Mapping and validation of the epistatic D and P genes controlling anthocyanin biosynthesis in the peel of eggplant (Solanum melongena L.) fruit. Hortic. Res. 2023, 10, uhac268. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Total Plants | Number of Green-Peel Plants | Number of White-Peel Plants | Expected Ratio | χ2a |
|---|---|---|---|---|---|
| 19141 (female parent) | 30 | - | 30 | - | - |
| 19143 (male parent) | 30 | 30 | - | - | - |
| F1 (20183) | 30 | 30 | 0 | - | - |
| F2 population | 267 | 199 | 68 | 3:1 | 0.03 |
| Code | Gene Name | Start Position | Stop Position | Nr Annotation |
|---|---|---|---|---|
| 1 | Smechr0802018 | 80,586,146 | 80,589,692 | PREDICTED: two-component response regulator-like APRR2 [Solanum tuberosum] |
| 2 | Smechr0802019 | 80,620,395 | 80,626,917 | putative methyltransferase PMT17 [Capsicum chinense] |
| 3 | Smechr0802020 | 80,628,319 | 80,638,536 | uncharacterized protein LOC107027127 [Solanum pennellii] |
| 4 | Smechr0802021 | 80,641,088 | 80,645,293 | vacuolar protein sorting-associated protein 55 homolog [Solanum pennellii] |
| 5 | Smechr0802022 | 80,646,220 | 80,653,320 | PREDICTED: phosphoacetylglucosamine mutase [Solanum tuberosum] |
| 6 | Smechr0802023 | 80,656,128 | 80,656,656 | PREDICTED: uncharacterized protein LOC102601084 [Solanum tuberosum] |
| 7 | Smechr0802024 | 80,658,611 | 80,661,728 | transcription factor MAMYB [Solanum lycopersicum] |
| 8 | Smechr0802025 | 80,662,057 | 80,666,530 | PREDICTED: UMP-CMP kinase 3-like [Solanum tuberosum] |
| 9 | Smechr0802026 | 80,674,673 | 80,688,072 | SUPPRESSOR OF ABI3-5 isoform X1 [Solanum lycopersicum] |
| 10 | Smechr0802027 | 80,689,088 | 80,691,353 | PREDICTED: pentatricopeptide repeat-containing protein At3g29230-like [Solanum tuberosum] |
| 11 | Smechr0802028 | 80,692,128 | 80,693,721 | PREDICTED: expansin-like B1 isoform X1 [Capsicum annuum] |
| 12 | Smechr0802029 | 80,699,208 | 80,706,124 | PREDICTED: haloacid dehalogenase-like hydrolase domain-containing protein Sgpp isoform X1 [Solanum tuberosum] |
| 13 | Smechr0802030 | 80,702,195 | 80,709,174 | PREDICTED: dynamin-related protein 1E-like [Solanum tuberosum] |
| 14 | Smechr0802031 | 80,712,567 | 80,714,826 | PREDICTED: probable purine permease 11 [Solanum tuberosum] |
| 15 | Smechr0802032 | 80,717,650 | 80,719,902 | PREDICTED: probable purine permease 11 [Solanum tuberosum] |
| 16 | Smechr0802033 | 80,722,938 | 80,729,560 | PREDICTED: uncharacterized mitochondrial protein ymf11 [Solanum tuberosum] |
| Gene Name | Average RPKM Value of 19143 | Average RPKM Value of 19141 | log2 (Fold Change) | Description |
|---|---|---|---|---|
| Smechr0802018 | 448.6 | 95.5 | 2.2 | PREDICTED: two-component response regulator-like APRR2 [Solanum tuberosum] |
| Smechr0802030 | 72.6 | 144.2 | −1.0 | PREDICTED: dynamin-related protein 1E-like [Solanum tuberosum] |
| Smechr0802033 | 149.2 | 221.7 | −0.6 | PREDICTED: uncharacterized mitochondrial protein ymf11 [Solanum tuberosum] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.; Jin, Q.; Li, Z.; Li, T.; Wang, Y.; You, Q.; Gong, C.; Heng, Z.; Sun, B. Fine Mapping and Candidate Gene Analysis of the Gv1 Locus Controlling Green-Peel Color in Eggplant (Solanum melongena L.). Horticulturae 2023, 9, 888. https://doi.org/10.3390/horticulturae9080888
Lv Z, Jin Q, Li Z, Li T, Wang Y, You Q, Gong C, Heng Z, Sun B. Fine Mapping and Candidate Gene Analysis of the Gv1 Locus Controlling Green-Peel Color in Eggplant (Solanum melongena L.). Horticulturae. 2023; 9(8):888. https://doi.org/10.3390/horticulturae9080888
Chicago/Turabian StyleLv, Zijian, Qingmin Jin, Zhiliang Li, Tao Li, Yikui Wang, Qian You, Chao Gong, Zhou Heng, and Baojuan Sun. 2023. "Fine Mapping and Candidate Gene Analysis of the Gv1 Locus Controlling Green-Peel Color in Eggplant (Solanum melongena L.)" Horticulturae 9, no. 8: 888. https://doi.org/10.3390/horticulturae9080888
APA StyleLv, Z., Jin, Q., Li, Z., Li, T., Wang, Y., You, Q., Gong, C., Heng, Z., & Sun, B. (2023). Fine Mapping and Candidate Gene Analysis of the Gv1 Locus Controlling Green-Peel Color in Eggplant (Solanum melongena L.). Horticulturae, 9(8), 888. https://doi.org/10.3390/horticulturae9080888