

Cost of Transport of Undulating Fin Propulsion

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Single-Finned Robot—Mechanical Design
2.2. Single-Finned Robot—Control
2.3. Double-Finned Robot—Mechanical Design
2.4. Experiments
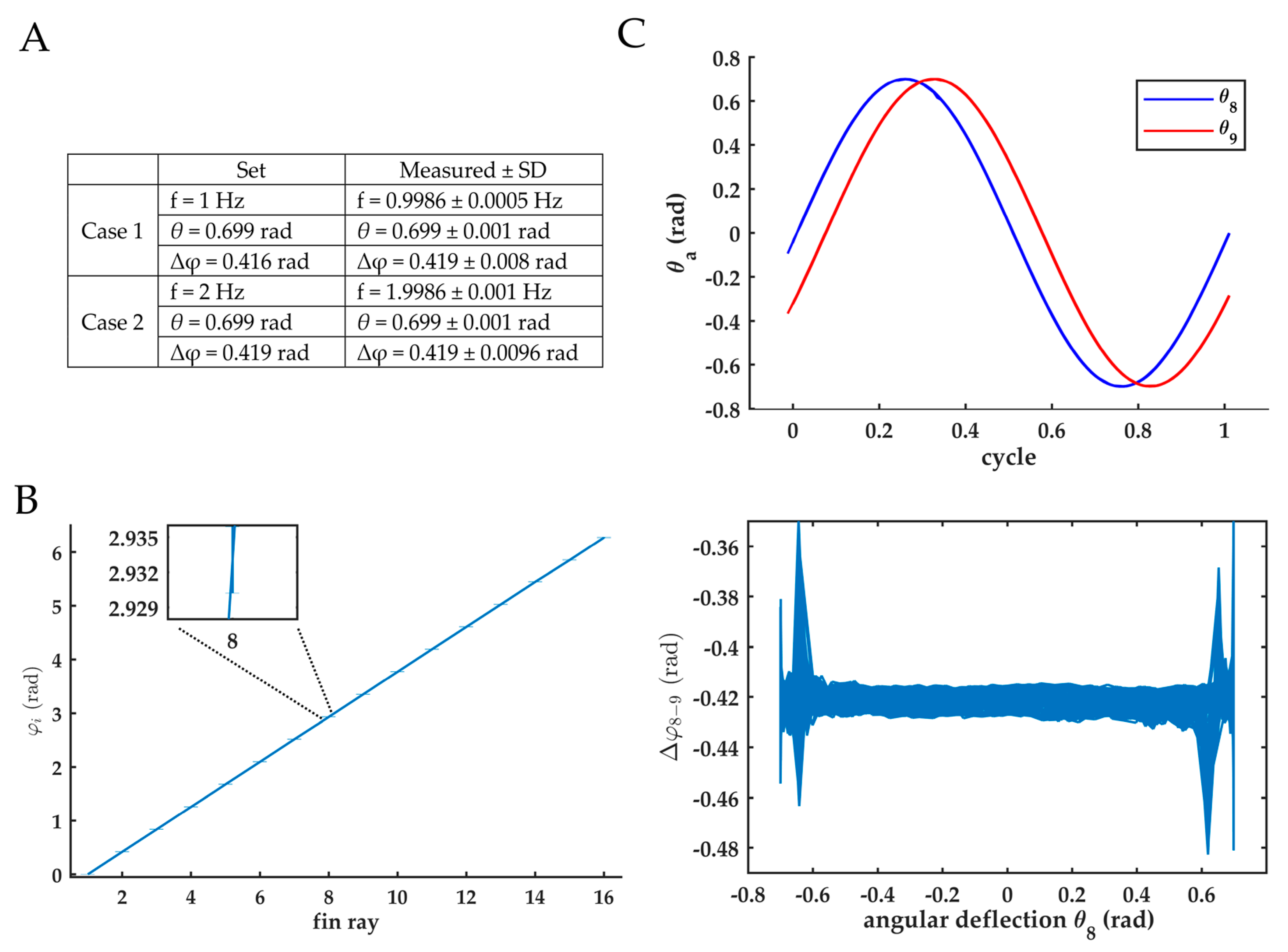
2.4.1. Accuracy of Fin Kinematics
2.4.2. Free-Swimming Measurements with a Single-Finned Robot
2.4.3. Free-Swimming Measurements with a Double-Finned Robot
3. Results
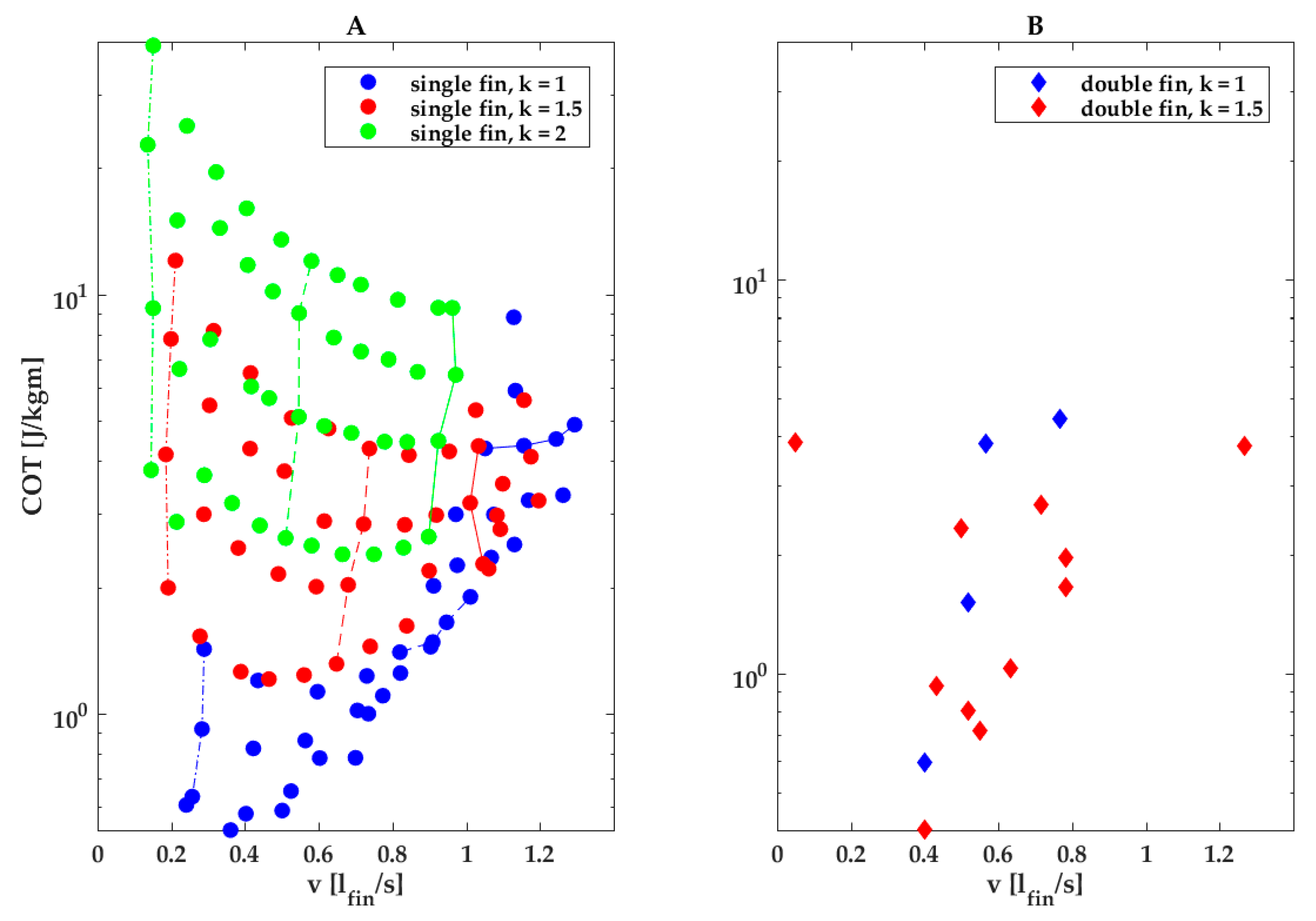
3.1. Single-Finned Robot Performance
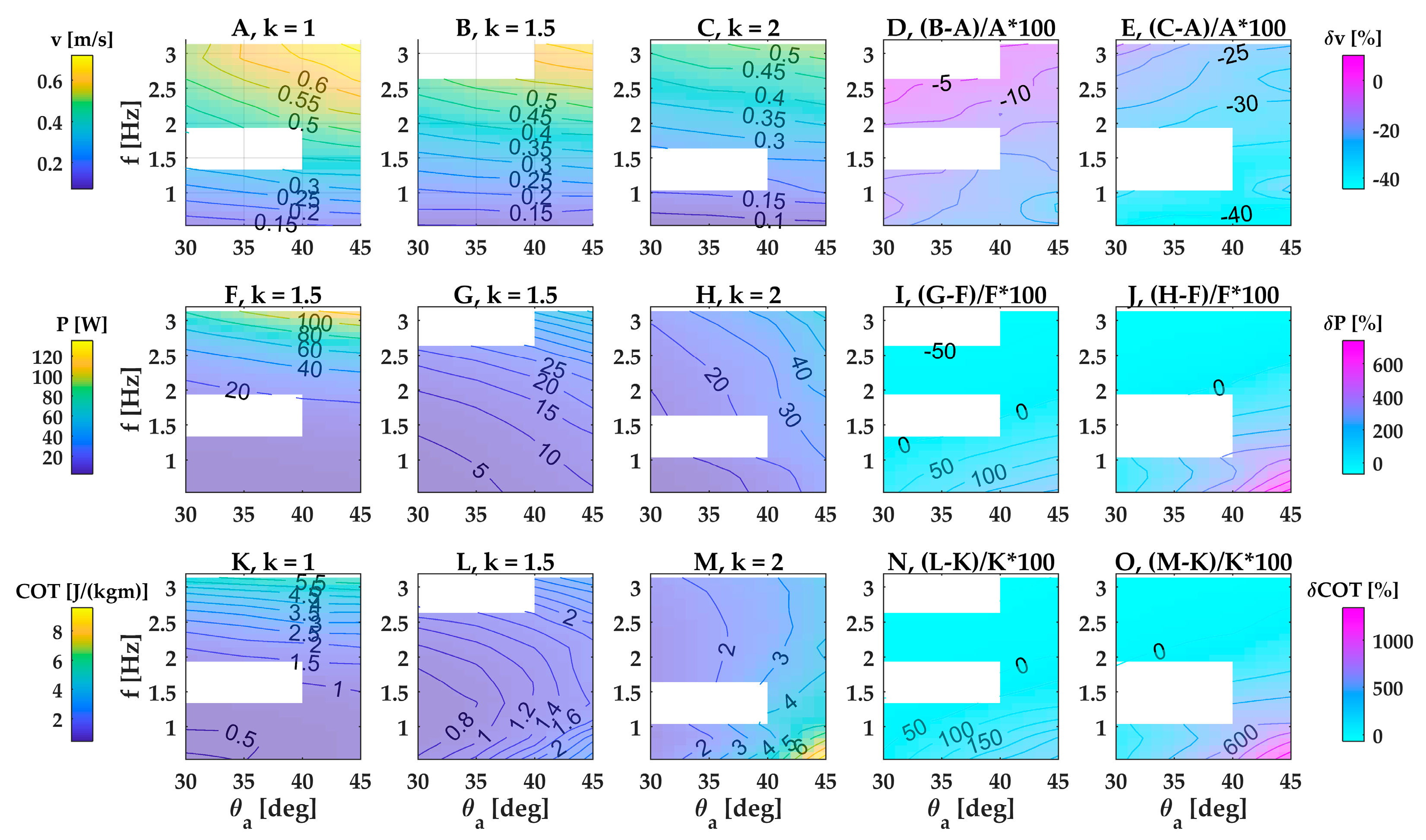
3.1.1. Velocity
3.1.2. Power Consumption
3.1.3. Cost of Transport
3.2. Velocity, Power Consumption and Cost of Transport of the Double-Finned Robot
4. Discussion
4.1. Comparing Kinematics and Swimming Speed among Robotic and Animal Swimmers
4.2. Comparing Power and Cost of Transport among Robotic and Animal Swimmers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Symbols and Experimental Parameters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Definition | Unit |
|---|---|---|
| Θi | Angular deflection | [deg] |
| θa | Angular amplitude | [deg] |
| θ0 | Offset angle | [deg] |
| λ | Wavelength | [mm] |
| φ | Phase angle | [deg] |
| COT | Cost of transport | [W/(kg m/s)] |
| f | Frequency | [Hz] |
| i | Fin ray number | [-] |
| I | Local current | [A] |
| k | Wavenumber | [-] |
| lf | Fin length | [mm] |
| m | Robot mass | [kg] |
| P | Power consumption | [W] |
| U | Local voltage | [V] |
| v | Steady average swimming speed | [m/s] |
| x | Position along fin base | [mm] |
| Parameter | Symbol | Values | Unit |
|---|---|---|---|
| fin height | — | 60, 100, 140 mm | mm |
| fin beat frequency | f | 0.55 to 3.19 Hz in steps of 0.264 Hz | Hz |
| fin beat amplitude | θa | 30° to 45° in steps of 5° | deg |
| wavenumber | k | 1, 1.5, 2 | — |
Appendix B. Accuracy of Fin Kinematics

Appendix C. Free-Swimming Performance of Small, Medium and Elliptic Fin




Appendix D. Velocity, Power and Cost of Transport of a Double-Finned Robot

References
- Sefati, S.; Neveln, I.D.; Roth, E.; Mitchell, T.R.; Snyder, J.B.; MacIver, M.A.; Fortune, E.S.; Cowan, N.J. Mutually opposing forces during locomotion can eliminate the tradeoff between maneuverability and stability. Proc. Natl. Acad. Sci. USA 2013, 110, 18798–18803. [Google Scholar] [CrossRef] [PubMed]
- Jastrebsky, R.A.; Bartol, I.K.; Krueger, P.S. Turning performance of brief squid Lolliguncula brevis during attacks on shrimp and fish. J. Exp. Biol. 2017, 220, 908–919. [Google Scholar] [PubMed]
- Bartol, I.K.; Krueger, P.S.; York, C.A.; Thompson, J.T. New approaches for assessing squid fin motions: Coupling proper orthogonal decomposition with volumetric particle tracking velocimetry. J. Exp. Biol. 2018, 221, jeb176750. [Google Scholar] [CrossRef] [PubMed]
- Toda, Y.; Suzuki, T.; Uto, S.; Tanaka, N. Fundamental study of a fishlike body with two undulating side-fins. In Bio-Mechanisms of Swimming and Flying; Springer: Berlin/Heidelberg, Germany, 2004; pp. 93–110. [Google Scholar]
- Simons, D.; Bergers, M.; Henrion, S.; Hulzenga, J.; Jutte, R.; Pas, W.; Van Schravendijk, M.; Vercruyssen, T.; Wilken, A. A highly versatile autonomous underwater vehicle with biomechanical propulsion. In Proceedings of the OCEANS 2009-EUROPE, Bremen, Germany, 11–14 May 2009; pp. 1–6. [Google Scholar]
- Chen, H.; Li, W.; Cui, W.; Yang, P.; Chen, L. Multi-objective multidisciplinary design optimization of a robotic fish system. J. Mar. Sci. Eng. 2021, 9, 478. [Google Scholar] [CrossRef]
- Yu, J.; Wang, M.; Dong, H.; Zhang, Y.; Wu, Z. Motion control and motion coordination of bionic robotic fish: A review. J. Bionic Eng. 2018, 15, 579–598. [Google Scholar] [CrossRef]
- Wang, R.; Wang, S.; Wang, Y.; Cheng, L.; Tan, M. Development and motion control of biomimetic underwater robots: A survey. IEEE Trans. Syst. Man Cybern. Syst. 2020, 52, 833–844. [Google Scholar] [CrossRef]
- Raj, A.; Thakur, A. Fish-inspired robots: Design, sensing, actuation, and autonomy—A review of research. Bioinspir. Biomim. 2016, 11, 31001. [Google Scholar] [CrossRef]
- Arslan, E.; Kadir, A. A design methodology for cuttlefish shaped amphibious robot. Avrupa Bilim Ve Teknol. Derg. 2019, 214–224. [Google Scholar] [CrossRef]
- Roper, D.; Sharma, S.; Sutton, R.; Culverhouse, P. A review of developments towards biologically inspired propulsion systems for autonomous underwater vehicles. Proc. Inst. Mech. Eng. Part M J. Eng. Marit. Environ. 2011, 225, 77–96. [Google Scholar] [CrossRef]
- Hu, T.; Shen, L.; Low, K. Bionic asymmetry: From amiiform fish to undulating robotic fins. Chin. Sci. Bull. 2009, 54, 562–568. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, H.; Wang, Z.; He, Q.; Chen, L.; Li, W.; Li, R.; Cui, W. A manta ray robot with soft material based flapping wing. J. Mar. Sci. Eng. 2022, 10, 962. [Google Scholar] [CrossRef]
- Henrion, S. Performance Analysis of Undulating Fin Propulsion. Master’s Thesis, Delft University of Technology, Delft, The Netherlands, 2010. [Google Scholar]
- Curet, O.M.; Patankar, N.A.; Lauder, G.V.; MacIver, M.A. Mechanical properties of a bio-inspired robotic knifefish with an undulatory propulsor. Bioinspir. Biomim. 2011, 6, 26004. [Google Scholar] [CrossRef] [PubMed]
- Vercruyssen, T.G.A. Phase Resolved piv Analysis of an Undulating Fin. Master’s Thesis, Delft University of Technology, Delft, The Netherlands, 2010. [Google Scholar]
- Neveln, I.D.; Bale, R.; Bhalla, A.P.S.; Curet, O.M.; Patankar, N.A.; MacIver, M.A. Undulating fins produce off-axis thrust and flow structures. J. Exp. Biol. 2014, 217, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Curet, O.M.; Patankar, N.A.; Lauder, G.V.; MacIver, M.A. Aquatic manoeuvering with counter-propagating waves: A novel locomotive strategy. J. R. Soc. Interface 2011, 8, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- English, I.; Liu, H.; Curet, O.M. Robotic device shows lack of momentum enhancement for gymnotiform swimmers. Bioinspir. Biomim. 2019, 14, 24001. [Google Scholar] [CrossRef]
- Lighthill, J.; Blake, R. Biofluiddynamics of balistiform and gymnotiform locomotion. Part 1. Biological background, and analysis by elongated-body theory. J. Fluid Mech. 1990, 212, 183–207. [Google Scholar] [CrossRef]
- Lighthill, J. Biofluiddynamics of balistiform and gymnotiform locomotion. Part 2. The pressure distribution arising in two-dimensional irrotational flow from a general symmetrical motion of a flexible flat plate normal to itself. J. Fluid Mech. 1990, 213, 1–10. [Google Scholar] [CrossRef]
- Lighthill, J. Biofluiddynamics of balistiform and gymnotiform locomotion. Part 3. Momentum enhancement in the presence of a body of elliptic cross-section. J. Fluid Mech. 1990, 213, 11–20. [Google Scholar] [CrossRef]
- Lighthill, J. Biofluiddynamics of balistiform and gymnotiform locomotion. Part 4. Short-wavelength limitations on momentum enhancement. J. Fluid Mech. 1990, 213, 21–28. [Google Scholar] [CrossRef]
- Liu, H.; Curet, O. Swimming performance of a bio-inspired robotic vessel with undulating fin propulsion. Bioinspi. Biomim. 2018, 13, 56006. [Google Scholar] [CrossRef]
- Maertens, A.; Triantafyllou, M.S.; Yue, D.K. Efficiency of fish propulsion. Bioinspir. Biomim. 2015, 10, 46013. [Google Scholar] [CrossRef] [PubMed]
- Lighthill, M.J. Large-amplitude elongated-body theory of fish locomotion. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1971, 179, 125–138. [Google Scholar]
- Eloy, C. Optimal Strouhal number for swimming animals. J. Fluids Struct. 2012, 30, 205–218. [Google Scholar] [CrossRef]
- Videler, J.J. Fish Swimming; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1993; Volume 10. [Google Scholar]
- Long, J.H.; Schumacher, J.; Livingston, N.; Kemp, M. Four flippers or two? Tetrapodal swimming with an aquatic robot. Bioinspir. Biomim. 2006, 1, 20. [Google Scholar] [CrossRef]
- Zhu, J.; White, C.; Wainwright, D.K.; Santo, V.D.; Lauder, G.V.; Bart-Smith, H. Tuna robotics: A high-frequency experimental platform exploring the performance space of swimming fishes. Sci. Robot. 2019, 4, eaax4615. [Google Scholar] [CrossRef] [PubMed]
- Eloy, C. On the best design for undulatory swimming. J. Fluid Mech. 2013, 717, 2. [Google Scholar] [CrossRef]
- Li, G.; Liu, H.; Müller, U.K.; Voesenek, C.J.; Van Leeuwen, J.L. Fishes regulate tail-beat kinematics to minimize speed-specific cost of transport. Proc. R. Soc. B 2021, 288, 20211601. [Google Scholar] [CrossRef] [PubMed]
- Schultz, W.W.; Webb, P.W. Power requirements of swimming: Do new methods resolve old questions? Integr. Comp. Biol. 2002, 42, 1018–1025. [Google Scholar] [CrossRef]
- Bale, R.; Hao, M.; Bhalla, A.P.S.; Patankar, N.A. Energy efficiency and allometry of movement of swimming and flying animals. Proc. Natl. Acad. Sci. USA 2014, 111, 7517–7521. [Google Scholar] [CrossRef] [PubMed]
- Karssen, D. Robotic Bipedal Running. Master’s Thesis, Delft University of Technology, Delft, The Netherlands, 2013. [Google Scholar]
- Rudinac, M. Exploration and Learning for Cognitive Robots. Master’s Thesis, Delft University of Technology, Delft, The Netherlands, 2013. [Google Scholar]
- Hu, T.; Shen, L.; Lin, L.; Xu, H. Biological inspirations, kinematics modeling, mechanism design and experiments on an undulating robotic fin inspired by Gymnarchus niloticus. Mech. Mach. Theory 2009, 44, 633–645. [Google Scholar] [CrossRef]
- Garrido-Jurado, S.; Muñoz-Salinas, R.; Madrid-Cuevas, F.J.; Marín-Jiménez, M.J. Automatic generation and detection of highly reliable fiducial markers under occlusion. Pattern Recognit. 2014, 47, 2280–2292. [Google Scholar] [CrossRef]
- Phillips, A.B.; Haroutunian, M.; Man, S.K.; Murphy, A.J.; Boyd, S.W.; Blake, J.I.R.; Griffiths, G. Nature in engineering for monitoring the oceans: Comparison of the energetic costs of marine animals and AUVs. In Further Advances in Unmanned Marine Vehicles; Roberts, G.N., Sutton, R., Eds.; Institution of Engineering and Technology (IET): Piscataway, NJ, USA, 2012; pp. 373–405. [Google Scholar]
- Blevins, E.L.; Lauder, G.V. Rajiform locomotion: Three-dimensional kinematics of the pectoral fin surface during swimming in the freshwater stingray Potamotrygon orbignyi. J. Exp. Biol. 2012, 215, 3231–3241. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, L.J. Pectoral fin locomotion in batoid fishes: Undulation versus oscillation. J. Exp. Biol. 2001, 204, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, L.J.; Westneat, M.W. Functional morphology of undulatory pectoral fin locomotion in the stingray Taeniura lymma (Chondrichthyes: Dasyatidae). J. Exp. Biol. 1999, 202, 3523–3539. [Google Scholar] [CrossRef]
- Youngerman, E.D.; Flammang, B.E.; Lauder, G.V. Locomotion of free-swimming ghost knifefish: Anal fin kinematics during four behaviors. Zoology 2014, 117, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Di Santo, V.; Blevins, E.L.; Lauder, G.V. Batoid locomotion: Effects of speed on pectoral fin deformation in the little skate, Leucoraja erinacea. J. Exp. Biol. 2017, 220, 705–712. [Google Scholar] [CrossRef]
- Ruiz-Torres, R.; Curet, O.M.; Lauder, G.V.; MacIver, M.A. Kinematics of the ribbon fin in hovering and swimming of the electric ghost knifefish. J. Exp. Biol. 2013, 216, 823–834. [Google Scholar] [CrossRef]
- White, C.H.; Lauder, G.V.; Bart-Smith, H. Tunabot Flex: A tuna-inspired robot with body flexibility improves high-performance swimming. Bioinspir. Biomim. 2021, 16, 26019. [Google Scholar] [CrossRef]
- Di Santo, V.; Kenaley, C.P.; Lauder, G.V. High postural costs and anaerobic metabolism during swimming support the hypothesis of a U-shaped metabolism–speed curve in fishes. Proc. Natl. Acad. Sci. USA 2017, 114, 13048–13053. [Google Scholar] [CrossRef]
- Bartol, I.K.; Mann, R.; Patterson, M.R. Aerobic respiratory costs of swimming in the negatively buoyant brief squid Lolliguncula brevis. J. Exp. Biol. 2001, 204, 3639–3653. [Google Scholar] [CrossRef]
- Tack, N.B.; Du Clos, K.T.; Gemmell, B.J. Anguilliform locomotion across a natural range of swimming speeds. Fluids 2021, 6, 127. [Google Scholar] [CrossRef]
- Santo, V.D.; Kenaley, C.P. Skating by: Low energetic costs of swimming in a batoid fish. J. Exp. Biol. 2016, 219, 1804–1807. [Google Scholar] [CrossRef] [PubMed]
- Sefati, S.; Neveln, I.; MacIver, M.A.; Fortune, E.S.; Cowan, N.J. Counter-propagating waves enhance maneuverability and stability: A bio-inspired strategy for robotic ribbon-fin propulsion. In Proceedings of the 2012 4th IEEE RAS & EMBS International Conference on Biomedical Robotics and Biomechatronics (BioRob), Rome, Italy, 24–27 June 2012; pp. 1620–1625. [Google Scholar]





| Swimmer | Frequency (Hz) | Amplitude | Wavenumber | vmax (L/s) 1 |
|---|---|---|---|---|
| robot | 0.55–3.19 | 30°–45° | 1–2 | 0.65 RL/s 1.3 FL/s |
| stingray | 1.5–3.0 2 0.78–4.2 3 | 0.21 (mid disk) DL 2 0.08–0.35 DL 3 | — 0.4–1.31 3 | — 0.9–3 DL/s 4 |
| knifefish | 6.74 5 5.1–9.3 6 | — 70° (max) 6 | 3.38 5 — | 0.51 BL/s 5 0.18–2.0 BL/s 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vercruyssen, T.G.A.; Henrion, S.; Müller, U.K.; van Leeuwen, J.L.; van der Helm, F.C.T. Cost of Transport of Undulating Fin Propulsion. Biomimetics 2023, 8, 214. https://doi.org/10.3390/biomimetics8020214
Vercruyssen TGA, Henrion S, Müller UK, van Leeuwen JL, van der Helm FCT. Cost of Transport of Undulating Fin Propulsion. Biomimetics. 2023; 8(2):214. https://doi.org/10.3390/biomimetics8020214
Chicago/Turabian StyleVercruyssen, Tim G. A., Sebastian Henrion, Ulrike K. Müller, Johan L. van Leeuwen, and Frans C. T. van der Helm. 2023. "Cost of Transport of Undulating Fin Propulsion" Biomimetics 8, no. 2: 214. https://doi.org/10.3390/biomimetics8020214
APA StyleVercruyssen, T. G. A., Henrion, S., Müller, U. K., van Leeuwen, J. L., & van der Helm, F. C. T. (2023). Cost of Transport of Undulating Fin Propulsion. Biomimetics, 8(2), 214. https://doi.org/10.3390/biomimetics8020214