
Biomimetic Cardiac Tissue Models for In Vitro Arrhythmia Studies
 , , ,
, , ,
Abstract
:1. Introduction
- Explore the mechanisms of arrhythmias in detail by isolating and examining individual cells or small cell clusters;
- Evaluate new drugs and treatments for arrhythmias by simulating the conditions in the heart and monitoring how they affect the heart’s electrical activity;
- Investigate the effects of different environmental factors on arrhythmias by exposing cells to various chemicals, toxins, and other environmental factors and measuring how they affect the heart’s electrical activity.
- The challenge of replicating the exact cellular composition and architecture of the human heart tissue;
- The variability and immaturity of iPSC-derived cardiac cells compared to native cardiac cells;
- The lack of standardized protocols and criteria for producing and assessing tissue models;
- The need for further validation and translation of tissue model findings to human clinical settings.
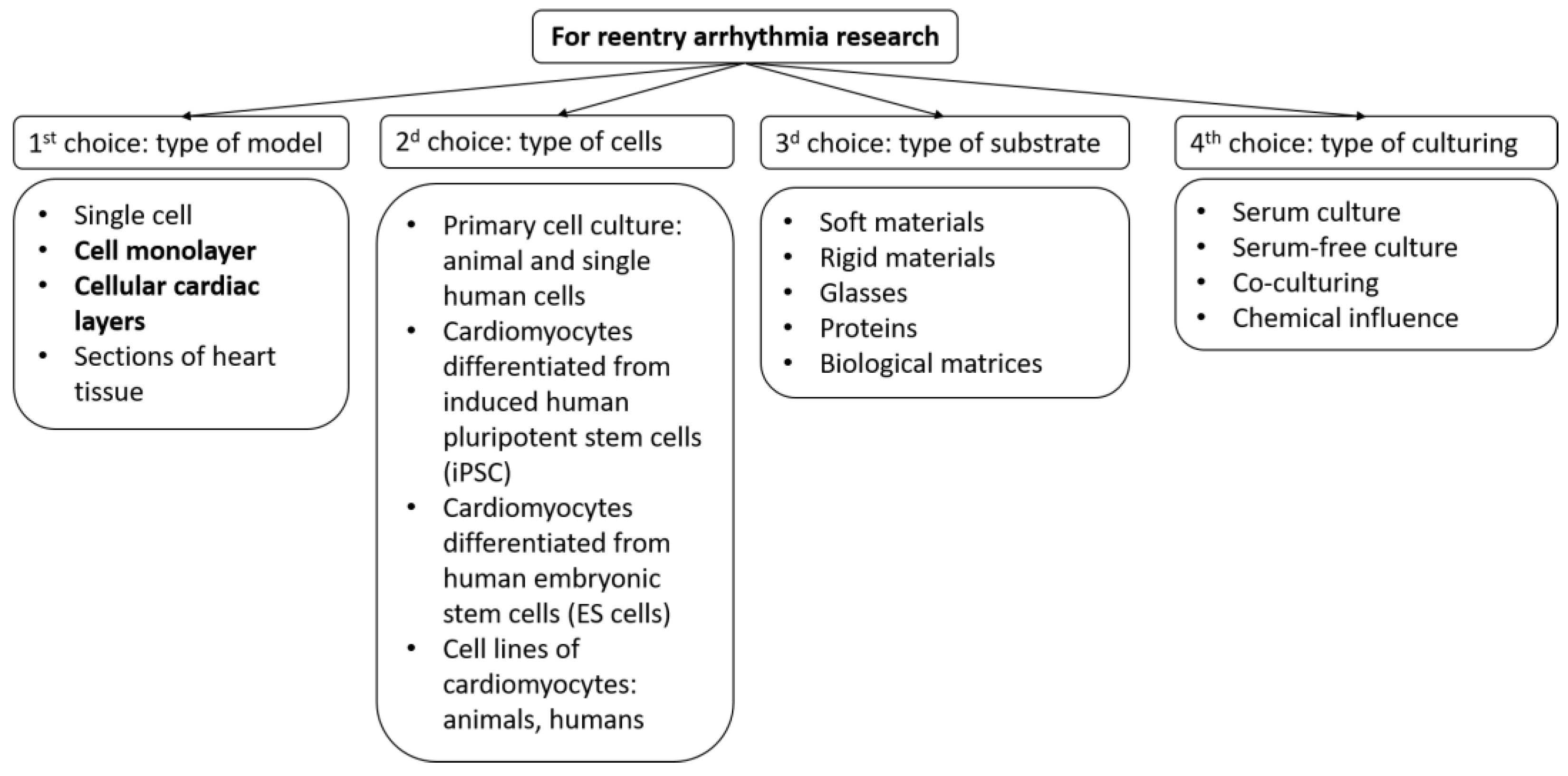
- Cell type;
- Type of substrates and coatings for cell seeding;
- Type of growth medium and external factors.
- Simulation of a conduction case in real heart tissue: fibrosis, anisotropy, domain location, and more;
- Congenital mutations leading to arrhythmias.
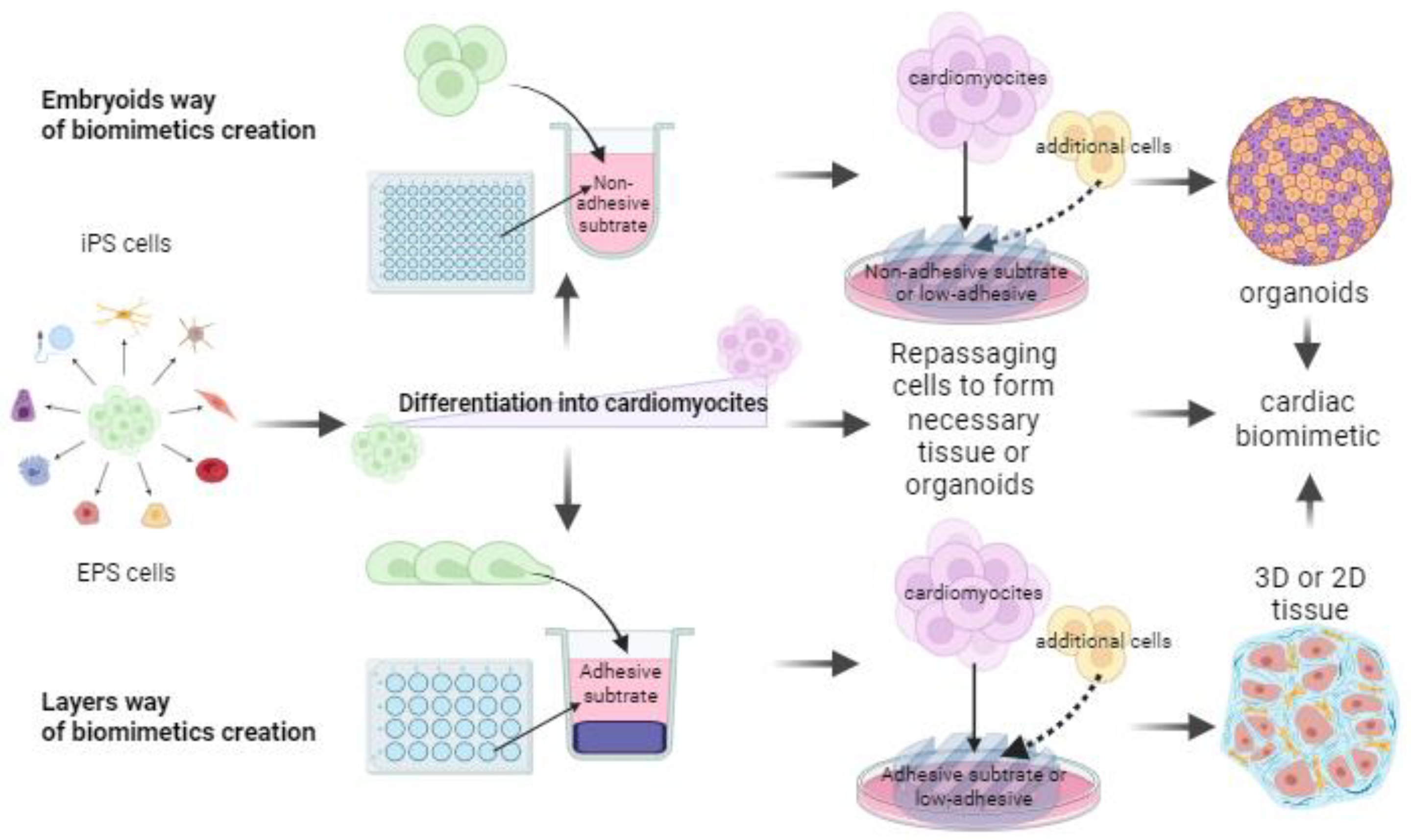
- Using different types of cells and rationale for their choice;
- By modeling the structural features of the tissue;
- Using various substrates and coatings;
- Using computer models based on experimental data.
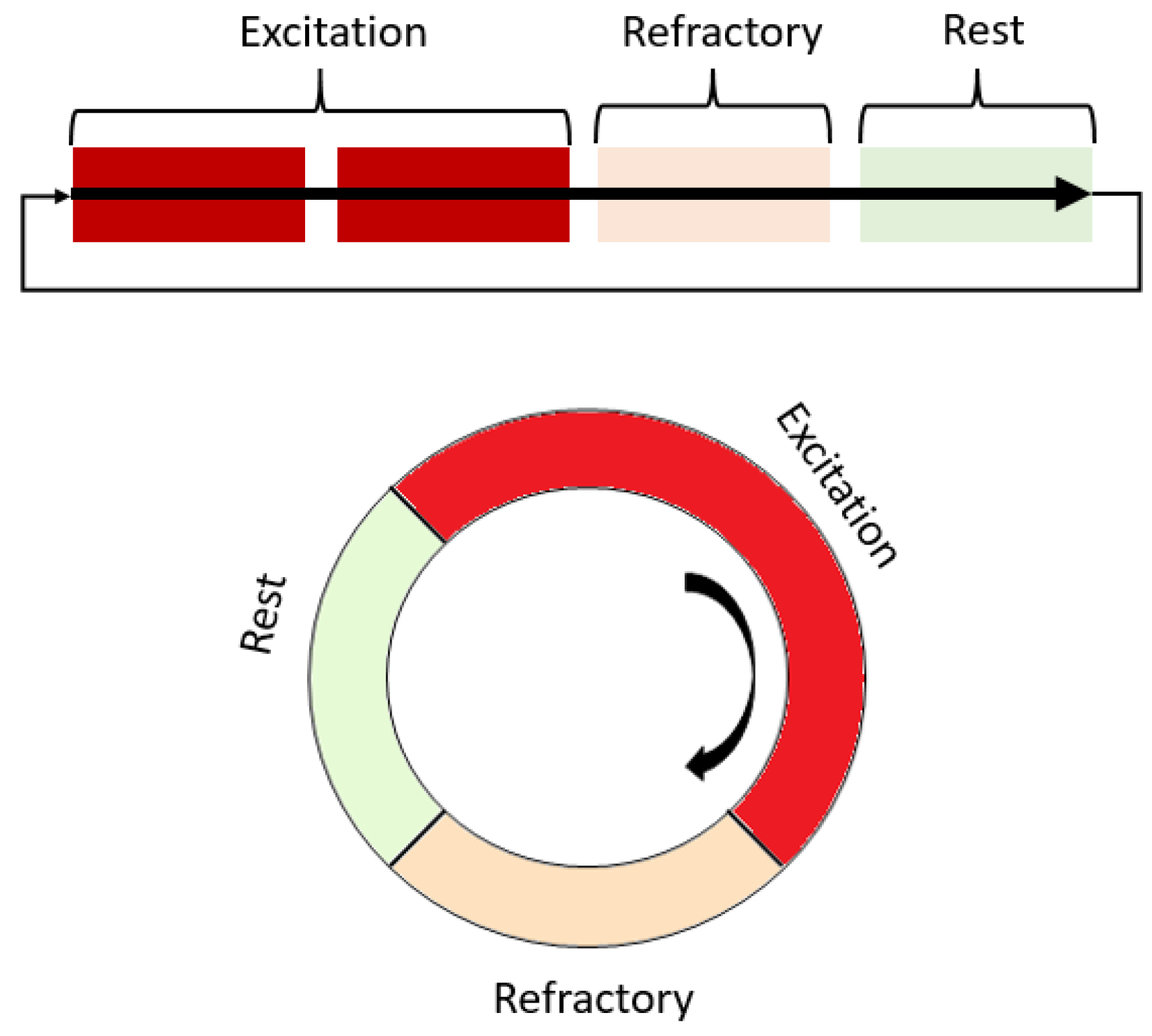
2. Reentry Arrhythmia in Cardiac Tissue Models
3. Modeling of Structural Features of Cardiac Tissue to Study the Occurrence of Reentry
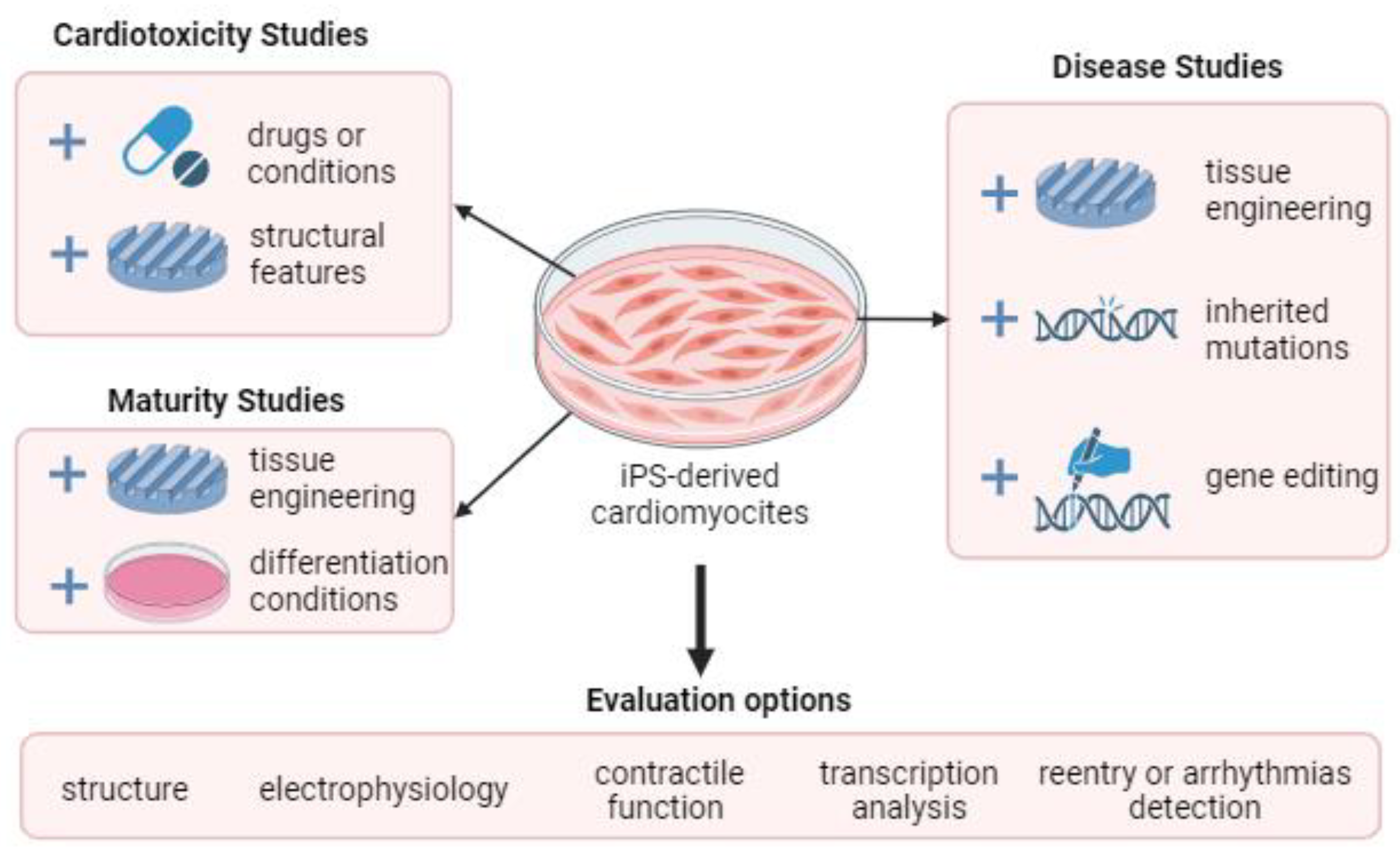
4. Application of Different Cell Types to Simulate Cardiac Tissue Models
5. Arrhythmogenicity Test as an Application of Cardiac Tissue Biomimetic
6. Variety of Substrates for Biomimetic Cardiac Tissue Models
7. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATP | Antitachycardia or overdrive pacing |
| CM | Cardiomyocytes |
| NRVM | Neonatal rat ventricular myocytes |
| iPSC | Induced pluripotent stem cells |
| hiPSC | Human induced pluripotent stem cells |
| iPS-CM | (iPSC)-derived cardiomyocytes |
| ESC | Embryonic stem cells |
| EHT | Engineered heart tissues |
| PMGI | Polymethylglutarimide |
| PCL | Poly(ϵ-caprolactone) |
| PLGA | Poly(lactide-co-glycolide) |
| PAA | Polyacrylamide |
| PDMS | Polydimethylsiloxane |
| PLLA | Poly-L-lactic acid |
| POMaC | Poly(octa methylene maleate-(anhydride) citrate) |
| ECM | Extracellular matrix |
| EPI cells | Epicardial cells |
| MEA | Multielectrode array |
| CPVT | Catecholaminergic polymorphic ventricular tachycardia |
References
- Nash, M.P.; Panfilov, A.V. Electromechanical model of excitable tissue to study reentrant cardiac arrhythmias. Prog. Biophys. Mol. Biol. 2004, 85, 501–522. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, T.A.; Medvescek, E.; Henriquez, C.S. Modeling dynamics in diseased cardiac tissue: Impact of model choice. Chaos 2017, 27, 093909. [Google Scholar] [CrossRef] [PubMed]
- Sumitomo, N.; Harada, K.; Nagashima, M.; Yasuda, T.; Nakamura, Y.; Aragaki, Y.; Saito, A.; Kurosaki, K.; Jouo, K.; Koujiro, M.; et al. Catecholaminergic polymorphic ventricular tachycardia: Electrocardiographic characteristics and optimal therapeutic strategies to prevent sudden death. Heart 2003, 89, 66–70. [Google Scholar] [CrossRef]
- Park, S.J.; Zhang, D.; Qi, Y.; Li, Y.; Lee, K.Y.; Bezzerides, V.J.; Yang, P.; Xia, S.; Kim, S.L.; Liu, X.; et al. Insights into the pathogenesis of catecholaminergic polymorphic ventricular tachycardia from engineered human heart tissue. Circulation 2019, 140, 390–404. [Google Scholar] [CrossRef] [PubMed]
- Bezzerides, V.J.; Caballero, A.; Wang, S.; Ai, Y.; Hylind, R.J.; Lu, F.; Heims-Waldron, D.A.; Chambers, K.D.; Zhang, D.; Abrams, D.J.; et al. Gene therapy for catecholaminergic polymorphic ventricular tachycardia by inhibition of Ca2+/calmodulin-dependent kinase II. Circulation 2019, 140, 405–419. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, T.; Virág, L.; Varró, A.; Rudy, Y. Simulation of the undiseased human cardiac ventricular action potential: Model formulation and experimental validation. PLoS Comput. Biol. 2011, 7, e1002061. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, S. Singularities: When time breaks down. Science 1987, 237, 1360. [Google Scholar] [CrossRef] [PubMed]
- Keener, J.P. A geometrical theory for spiral waves in excitable media. SIAM J. Appl. Math. 1986, 46, 1039–1056. [Google Scholar] [CrossRef]
- Markus, M.; Hess, B. Isotropic cellular automaton for modelling excitable media. Nature 1990, 347, 56–58. [Google Scholar] [CrossRef]
- Kadota, S.; Minami, I.; Morone, N.; Heuser, J.E.; Agladze, K.; Nakatsuji, N. Development of a reentrant arrhythmia model in human pluripotent stem cell-derived cardiac cell sheets. Eur. Heart J. 2013, 34, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Agladze, K.; Keener, J.P.; Müller, S.C.; Panfilov, A. Rotating spiral waves created by geometry. Science 1994, 264, 1746–1748. [Google Scholar] [CrossRef] [PubMed]
- Agladze, K.; Magome, N.; Aliev, R.; Yamaguchi, T.; Yoshikawa, K. Finding the optimal path with the aid of chemical wave. Phys. D 1997, 106, 247–254. [Google Scholar] [CrossRef]
- Yoshio, W.; Dreifus, L.S. Inhomogeneous conduction in the AV node: A model for re-entry. Am. Heart J. 1965, 70, 505–514. [Google Scholar]
- Pertsov, A.M.; Davidenko, J.M.; Salomonsz, R.; Baxter, W.T.; Jalife, J. Spiral waves of excitation underlie reentrant activity in isolated cardiac muscle. Circ. Res. 1993, 72, 631–650. [Google Scholar] [CrossRef]
- Agladze, K.; Kay, M.W.; Krinsky, V.; Sarvazyan, N. Interaction between spiral and paced waves in cardiac tissue. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H503–H513. [Google Scholar] [CrossRef]
- Charlton, N.P.; Lawrence, D.T.; Brady, W.J.; Kirk, M.A.; Holstege, C.P. Termination of drug-induced torsades de pointes with overdrive pacing. Am. J. Emerg. Med. 2010, 28, 95–102. [Google Scholar] [CrossRef]
- Volle, K.; Delmas, C.; Rollin, A.; Voglimacci-Stephanopoli, Q.; Mondoly, P.; Cariou, E.; Mandel, F.; Delasnerie, H.; Beneyto, M.; Galinier, M.; et al. Successful reversal of severe tachycardia-induced cardiomyopathy with cardiogenic shock by urgent rhythm or rate control: Only rhythm and rate matter. J. Clin. Med. 2021, 10, 4504. [Google Scholar] [CrossRef]
- Leventopoulos, G.; Koros, R.; Travlos, C.; Perperis, A.; Chronopoulos, P.; Tsoni, E.; Koufou, E.-E.; Papageorgiou, A.; Apostolos, A.; Kaouris, P.; et al. Mechanisms of Atrial Fibrillation: How Our Knowledge Affects Clinical Practice. Life 2023, 13, 1260. [Google Scholar] [CrossRef]
- Rosman, J.Z.; John, R.M.; Stevenson, W.G.; Epstein, L.M.; Tedrow, U.B.; Koplan, B.A.; Albert, C.M.; Michaud, G.F. Resetting criteria during ventricular overdrive pacing successfully differentiate orthodromic reentrant tachycardia from atrioventricular nodal reentrant tachycardia despite interobserver disagreement concerning QRS fusion. Heart Rhythm. 2011, 8, 2–7. [Google Scholar] [CrossRef]
- Bursac, N.; Tung, L. Acceleration of functional reentry by rapid pacing in anisotropic cardiac monolayers: Formation of multi-wave functional reentries. Cardiovasc. Res. 2006, 69, 381–390. [Google Scholar] [CrossRef]
- Ripplinger, C.M.; Krinsky, V.I.; Nikolski, V.P.; Efimov, I.R. Mechanisms of unpinning and termination of ventricular tachycardia. Am. J. Physiol.-Heart Circ. Physiol. 2006, 291, H184–H192. [Google Scholar] [CrossRef] [PubMed]
- Rappel, W.J.; Zaman, J.A.; Narayan, S.M. Mechanisms for the Termination of Atrial Fibrillation by Localized Ablation: Computational and Clinical Studies. Circ. Arrhythm. Electrophysiol. 2015, 8, 1325–1333. [Google Scholar] [CrossRef] [PubMed]
- Hussaini, S.; Venkatesan, V.; Biasci, V.; Romero Sepúlveda, J.M.; Quiñonez Uribe, R.A.; Sacconi, L.; Bub, G.; Richter, C.; Krinski, V.; Parlitz, U.; et al. Drift and termination of spiral waves in optogenetically modified cardiac tissue at sub-threshold illumination. Elife 2021, 10, e59954. [Google Scholar] [CrossRef] [PubMed]
- Ten Tusscher, K.H.; Panfilov, A.V. Reentry in heterogeneous cardiac tissue described by the Luo-Rudy ventricular action potential model. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H542–H548. [Google Scholar] [CrossRef]
- Fenton, F.H.; Cherry, E.M.; Hastings, H.M.; Evans, S.J. Multiple mechanisms of spiral wave breakup in a model of cardiac electrical activity. Chaos 2002, 12, 852–892. [Google Scholar] [CrossRef]
- Tung, L.; Zhang, Y. Optical imaging of arrhythmias in tissue culture. J. Electrocardiol. 2006, 39, S2–S6. [Google Scholar] [CrossRef]
- Cabo, C.; Pertsov, A.M.; Baxter, W.T.; Davidenko, J.M.; Gray, R.A.; Jalife, J. Wave-front curvature as a cause of slow conduction and block in isolated cardiac muscle. Circ. Res. 1994, 75, 1014–1028. [Google Scholar] [CrossRef]
- Fast, V.G.; Kleber, A.G. Role of wavefront curvature in propagation of cardiac impulse. Cardiovasc. Res. 1997, 33, 258–271. [Google Scholar] [CrossRef]
- Pravdin, S.F.; Nezlobinsky, T.V.; Panfilov, A.V.; Dierckx, H. High-frequency pacing of scroll waves in a three-dimensional slab model of cardiac tissue. Phys. Rev. E 2021, 103, 042420. [Google Scholar] [CrossRef]
- Isomura, A.; Hörning, M.; Agladze, K.; Yoshikawa, K. Eliminating spiral waves pinned to an anatomical obstacle in cardiac myocytes by high-frequency stimuli. Phys. Rev. E 2008, 78, 066216. [Google Scholar] [CrossRef]
- Riefolo, F.; Matera, C.; Garrido-Charles, A.; Gomila, A.M.J.; Sortino, R.; Agnetta, L.; Claro, E.; Masgrau, R.; Holzgrabe, U.; Batlle, M.; et al. Optical control of cardiac function with a photoswitchable muscarinic agonist. J. Am. Chem. Soc. 2019, 141, 7628–7636. [Google Scholar] [CrossRef] [PubMed]
- Nizamieva, A.A.; Kalita, I.Y.; Slotvitsky, M.M.; Berezhnoy, A.K.; Shubina, N.S.; Frolova, S.R.; Tsvelaya, V.A.; Agladze, K.I. Conduction of excitation waves and reentry drift on cardiac tissue with simulated photocontrol-varied excitability. Chaos 2023, 33, 023112. [Google Scholar] [CrossRef] [PubMed]
- Frolova, S.R.; Gaiko, O.; Tsvelaya, V.A.; Pimenov, O.Y.; Agladze, K.I. Photocontrol of voltage-gated ion channel activity by azobenzene trimethylammonium bromide in neonatal Rat cardiomyocytes. PLoS ONE 2016, 11, e0152018. [Google Scholar] [CrossRef]
- Bingen, B.O.; Engels, M.C.; Schalij, M.J.; Jangsangthong, W.; Neshati, Z.; Feola, I.; Ypey, D.L.; Askar, S.F.A.; Panfilov, A.V.; Pijnappels, D.A.; et al. Light-induced termination of spiral wave arrhythmias by optogenetic engineering of atrial cardiomyocytes. Cardiovasc. Res. 2014, 104, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Majumder, R.; Feola, I.; Teplenin, A.S.; de Vries, A.A.; Panfilov, A.V.; Pijnappels, D.A. Optogenetics enables real-time spatiotemporal control over spiral wave dynamics in an excitable cardiac system. Elife 2018, 7, e41076. [Google Scholar] [CrossRef]
- Kadota, S.; Kay, M.W.; Magome, N.; Agladze, K. Curvature-dependent excitation propagation in cultured cardiac tissue. JETP Lett. 2012, 94, 824–830. [Google Scholar] [CrossRef]
- Roth, B.J. Bidomain modeling of electrical and mechanical properties of cardiac tissue. Biophys. Rev. 2021, 2, 041301. [Google Scholar] [CrossRef]
- Gray, R.A.; Jalife, J.; Panfilov, A.; Baxter, W.T.; Cabo, C.; Davidenko, J.M.; Pertsov, A.M. Nonstationary vortexlike reentrant activity as a mechanism of polymorphic ventricular tachycardia in the isolated rabbit heart. Circulation 1995, 91, 2454–2469. [Google Scholar] [CrossRef]
- Ugarte, J.P.; Tobón, C.; Lopes, A.M.; Machado, J.A.T. Atrial rotor dynamics under complex fractional order diffusion. Front. Physiol. 2018, 9, 975. [Google Scholar] [CrossRef]
- Kachalov, V.N.; Tsvelaya, V.A.; Kudryashova, N.N.; Agladze, K.I. Success of spiral wave unpinning from heterogeneity in a cardiac tissue depends on its boundary conditions. JETP Lett. 2017, 106, 608–612. [Google Scholar] [CrossRef]
- Sládek, T.; Filkuka, J.; Dolezel, S.; Vasků, J.; Hartmannová, B.; Trávnícková, J. The border zone of the early myocardial infarction in dogs; its characteristics and viability. Basic Res. Cardiol. 1984, 79, 344–349. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, F.; Wang, C.; Ma, J. Stability of target waves inexcitable mediaunder electromagnetic induction and radiation. Phys. A Stat. Mech. Its Appl. 2019, 521, 519–530. [Google Scholar] [CrossRef]
- Zykov, V.; Krekhov, A.; Bodenschatz, E. Fast propagation regions cause self-sustained reentry in excitable media. Proc. Natl. Acad. Sci. USA 2017, 114, 1281–1286. [Google Scholar] [CrossRef]
- Orlova, Y.; Magome, N.; Liu, L.; Chen, Y.; Agladze, K. Electrospun nanofibers as a tool for architecture control in engineered cardiac tissue. Biomaterials 2011, 32, 5615–5624. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, L.; Guo, B.; Ma, P.X. Interwoven aligned conductive nanofiber yarn/hydrogel composite scaffolds for engineered 3D cardiac anisotropy. Acs Nano. 2017, 11, 5646–5659. [Google Scholar] [CrossRef]
- Kudryashova, N.N.; Teplenin, A.S.; Orlova, Y.V.; Selina, L.V.; Agladze, K. Arrhythmogenic role of the border between two areas of cardiac cell alignment. J. Mol. Cell. Cardiol. 2014, 76, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Kazbanov, I.V.; Ten Tusscher, K.H.W.J.; Panfilov, A.V. Effects of heterogenic diffuse fibrosis on ar-rhythmia dynamics and mechanism. Sci. Rep. 2016, 6, 20835. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.G.; Zhang, Y.; Chang, C.Y.; Xu, L.; Emokpae, R.; Tung, L.; Marbán, E.; Abraham, M.R. Spiral waves and reentry dynamics in an in vitro model of the healed infarct border zone. Circ. Res. 2009, 105, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Kawatou, M.; Masumoto, H.; Fukushima, H.; Morinaga, G.; Sakata, R.; Ashihara, T.; Yamashita, J.K. Modeling Torsade de Pointes arrhythmias in vitro in 3D human iPS cell-engineered heart tissue. Nat. Commun. 2017, 8, 1078. [Google Scholar] [CrossRef]
- Lin, B.; Li, Y.; Han, L.; Kaplan, A.D.; Ao, Y.; Kalra, S.; Bett, G.C.L.; Rasmusson, R.L.; Denning, C.; Yang, L. Modeling and study of the mechanism of dilated cardiomyopathy using induced pluripotent stem cells derived from individuals with Duchenne muscular dystrophy. Dis. Models Mech. 2015, 8, 457–466. [Google Scholar] [CrossRef]
- Li, W.; Stauske, M.; Luo, X.; Wagner, S.; Vollrath, M.; Mehnert, C.S.; Schubert, M.; Cyganek, L.; Chen, S.; Hasheminasab, S.-M.; et al. Disease Phenotypes and Mechanisms of iPSC-Derived Cardiomyocytes from Brugada Syndrome Patients with a Loss-of-Function SCN5A Mutation. Front. Cell Dev. Biol. 2020, 8, 592893. [Google Scholar] [CrossRef]
- Goldfracht, I.; Efraim, Y.; Shinnawi, R.; Kovalev, E.; Huber, I.; Gepstein, A.; Arbel, G.; Shaheen, N.; Tiburcy, M.; Zimmermann, W.H.; et al. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 2019, 92, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, A.; Shimizu, Y.; Okada, M.; Otani, K.; Yamawaki, H. Preventive Effect of Canstatin against Ventricular Arrhythmia Induced by Ischemia/Reperfusion Injury: A Pilot Study. Int. J. Mol. Sci. 2021, 22, 1004. [Google Scholar] [CrossRef] [PubMed]
- Heras-Bautista, C.O.; Carlos, O.; Mikhael, N.; Lam, J.; Shinde, V.; Katsen-Globa, A.; Dieluweit, S.; Molcanyi, M.; Uvarov, V.; Jütten, P.; et al. Cardiomyocytes facing fibrotic conditions re-express extracellular matrix transcripts. Acta Biomater. 2019, 89, 180–192. [Google Scholar] [CrossRef]
- Gunawan, M.G.; Sangha, S.S.; Shafaattalab, S.; Lin, E.; Heims-Waldron, D.A.; Bezzerides, V.J.; Laksman, Z.; Tibbits, G.F. Drug screening platform using human induced pluripotent stem cell-derived atrial cardiomyocytes and optical mapping. Stem Cells Transl. Med. 2021, 10, 68–82. [Google Scholar] [CrossRef] [PubMed]
- El-Battrawy, I.; Albers, S.; Cyganek, L.; Zhao, Z.; Lan, H.; Li, X.; Xu, Q.; Kleinsorge, M.; Huang, M.; Liao, Z.; et al. A cellular model of Brugada syndrome with SCN10A variants using human-induced pluripotent stem cell-derived cardiomyocytes. Europace 2019, 21, 1410–1421. [Google Scholar] [CrossRef] [PubMed]
- Shinnawi, R.; Shaheen, N.; Huber, I.; Shiti, A.; Arbel, G.; Gepstein, A.; Ballan, N.; Setter, N.; Tijsen, A.J.; Borggrefe, M.; et al. Modeling reentry in the short QT syndrome with human-induced pluripotent stem cell–derived cardiac cell sheets. J. Am. Coll. Cardiol. 2019, 73, 2310–2324. [Google Scholar] [CrossRef]
- Shiti, A.; Arbil, G.; Shaheen, N.; Huber, I.; Setter, N.; Gepstein, L. Utilizing human induced pluripotent stem cells to study atrial arrhythmias in the short QT syndrome. J. Mol. Cell Cardiol. 2023, 183, 42–53. [Google Scholar] [CrossRef] [PubMed]
- El-Battrawy, I.; Lan, H.; Cyganek, L.; Zhao, Z.; Li, X.; Buljubasic, F.; Lang, S.; Yücel, G.; Sattler, K.; Zimmermann, W.H.; et al. Modeling Short QT Syndrome Using Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes. J. Am. Heart Assoc. 2018, 7, e007394. [Google Scholar] [CrossRef]
- Giacomelli, E.; Sala, L.; Oostwaard, D.W.; Bellin, M. Cardiac microtissues from human pluripotent stem cells recapitulate the phenotype of long-QT syndrome. Biochem. Biophys. Res. Commun. 2021, 572, 118–124. [Google Scholar] [CrossRef]
- Bhagwan, J.R.; Mosqueira, D.; Chairez-Cantu, K.; Mannhardt, I.; Bodbin, S.E.; Bakar, M.; Smith, J.G.W.; Denning, C. Isogenic models of hypertrophic cardiomyopathy unveil differential phenotypes and mechanism-driven therapeutics. J. Mol. Cell Cardiol. 2020, 145, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Margara, F.; Psaras, Y.; Wang, Z.J.; Schmid, M.; Doste, R.; Garfinkel, A.C.; Repetti, G.G.; Seidman, J.G.; Seidman, C.E.; Rodriguez, B.; et al. Mechanism based therapies enable personalised treatment of hypertrophic cardiomyopathy. Sci Rep. 2022, 12, 22501. [Google Scholar] [CrossRef]
- Kohela, A.; van Kampen, S.J.; Moens, T.; Wehrens, M.; Molenaar, B.; Boogerd, C.J.; Monshouwer-Kloots, J.; Perini, I.; Goumans, M.J.; Smits, A.M.; et al. Epicardial Differentiation Drives Fibro-Fatty Remodeling in Arrhythmogenic Cardiomyopathy. Sci. Transl. Med. 2021, 13, eabf2750. [Google Scholar] [CrossRef] [PubMed]
- Bliley, J.M.; Vermeer, M.C.S.C.; Duffy, R.M.; Batalov, I.; Kramer, D.; Tashman, J.W.; Shiwarski, D.J.; Lee, A.; Teplenin, A.S.; Volkers, L.; et al. Dynamic loading of human engineered heart tissue enhances contractile function and drives a desmosome-linked disease phenotype. Sci. Transl. Med. 2021, 13, eabd1817. [Google Scholar] [CrossRef] [PubMed]
- Goldfracht, I.; Protze, S.; Shiti, A.; Setter, N.; Gruber, A.; Shaheen, N.; Nartiss, Y.; Keller, G.; Gepstein, L. Generating ring-shaped engineered heart tissues from ventricular and atrial human pluripotent stem cell-derived cardiomyocytes. Nat. Commun. 2020, 11, 75. [Google Scholar] [CrossRef]
- Richards, D.J.; Li, Y.; Kerr, C.M.; Yao, J.; Beeson, G.C.; Coyle, R.C.; Chen, X.; Jia, J.; Damon, B.; Wilson, R.; et al. Human cardiac organoids for the modelling of myocardial infarction and drug cardiotoxicity. Nat. Biomed. Eng. 2020, 4, 446–462. [Google Scholar] [CrossRef]
- MacQueen, L.A.; Sheehy, S.P.; Chantre, C.O.; Zimmerman, J.F.; Pasqualini, F.S.; Liu, X.; Goss, J.A.; Campbell, P.H.; Gonzalez, G.M.; Park, S.J.; et al. Addendum: A tissue-engineered scale model of the heart ventricle. Nat. Biomed. Eng. 2022, 6, 1318. [Google Scholar] [CrossRef]
- Ghosheh, M.; Ehrlich, A.; Ioannidis, K.; Ayyash, M.; Goldfracht, I.; Cohen, M.; Fischer, A.; Mintz, Y.; Gepstein, L.; Nahmias, Y. Electro-metabolic coupling in multi-chambered vascularized human cardiac organoids. Nat. Biomed. Eng. 2023, 1–21. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, Y.J.; Son, M.Y.; Oh, M.S.; Kim, J.; Ryu, B.; Kang, K.R.; Baek, J.; Chung, G.; Woo, D.H.; et al. Generation of human iPSCs derived heart organoids structurally and functionally similar to heart. Biomaterials 2022, 290, 121860. [Google Scholar] [CrossRef]
- Liang, P.Y.; Chang, Y.; Jin, G.; Lian, X.; Bao, X. Wnt signaling directs human pluripotent stem cells into vascularized cardiac organoids with chamber-like structures. Front. Bioeng. Biotechnol. 2022, 10, 1059243. [Google Scholar] [CrossRef]
- Hoang, P.; Wang, J.; Conklin, B.R.; E Healy, K.; Ma, Z. Generation of spatial-patterned early-developing cardiac organoids using human pluripotent stem cells. Nat. Protoc. 2018, 13, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, P.; Jahnel, S.M.; Papai, N.; Giesshammer, M.; Deyett, A.; Schmidt, C.; Penc, M.; Tavernini, K.; Grdseloff, N.; Meledeth, C.; et al. Cardioids reveal self-organizing principles of human cardiogenesis. Cell 2021, 184, 3299–3317. [Google Scholar] [CrossRef] [PubMed]
- Drakhlis, L.; Biswanath, S.; Farr, C.-M.; Lupanow, V.; Teske, J.; Ritzenhoff, K.; Franke, A.; Manstein, F.; Bolesani, E.; Kempf, H.; et al. Human heart-forming organoids recapitulate early heart and foregut development. Nat. Biotechnol. 2021, 39, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.; Henley, T.; Rossi, S.; Costello, M.J.; Polacheck, W.; Griffith, B.E.; Bressan, M. Adherens junction engagement regulates functional patterning of the cardiac pacemaker cell lineage. Dev. Cell. 2021, 56, 1498–1511. [Google Scholar] [CrossRef]
- Cimmino, C.; Netti, P.A.; Ventre, M. A switchable light-responsive azopolymer conjugating protein micropatterns with topography for mechanobiological studies. Front. Bioeng. Biotechnol. 2022, 10, 933410. [Google Scholar] [CrossRef]
- Caluori, G.; Pribyl, J.; Cmiel, V.; Pesl, M.; Potocnak, T.; Provaznik, I.; Skladal, P.; Rotrekl, V. Simultaneous study of mechanobiology and calcium dynamics on hESC-derived cardiomyocytes clusters. J. Mol. Recognit. 2019, 32, e2760. [Google Scholar] [CrossRef]
- Caluori, G.; Pribyl, J.; Pesl, M.; Jelinkova, S.; Rotrekl, V.; Skladal, P.; Raiteri, R. Non-invasive electromechanical cell-based biosensors for improved investigation of 3D cardiac models. Biosens. Bioelectron. 2019, 124, 129–135. [Google Scholar] [CrossRef]
- Lemme, M.; Ulmer, B.M.; Lemoine, M.D.; Zech, A.T.L.; Flenner, F.; Ravens, U.; Reichenspurner, H.; Rol-Garcia, M.; Smith, G.; Hansen, A.; et al. Atrial-like engineered heart tissue: An in vitro model of the human atrium. Stem. Cell Rep. 2018, 11, 1378–1390. [Google Scholar] [CrossRef]
- Shum, A.M.; Che, H.; Wong, A.O.; Zhang, C.; Wu, H.; Chan, C.W.; Costa, K.; Khine, M.; Kong, C.W.; Li, R.A. A micropatterned human pluripotent stem cell-based ventricular cardiac anisotropic sheet for visualizing drug-induced arrhythmogenicity. Adv. Mater. 2017, 29, 1602448. [Google Scholar] [CrossRef]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A platform for generation of chamber-specific cardiac tissues and disease modeling. Cell 2019, 176, 913–927.e18. [Google Scholar] [CrossRef]
- Slotvitsky, M.; Tsvelaya, V.; Frolova, S.; Dementyeva, E.; Agladze, K. Arrhythmogenicity test based on a human-induced pluripotent stem cell (iPSC)-derived cardiomyocyte layer. Toxicol. Sci. 2019, 168, 70–77. [Google Scholar] [CrossRef]
- Acun, A.; Nguyen, T.D.; Zorlutuna, P. In vitro aged, hiPSC-origin engineered heart tissue models with age-dependent functional deterioration to study myocardial infarction. Acta Biomater. 2019, 94, 372–391. [Google Scholar] [CrossRef] [PubMed]
- Jackman, C.P.; Ganapathi, A.M.; Asfour, H.; Qian, Y.; Allen, B.W.; Li, Y.; Bursac, N. Engineered cardiac tissue patch maintains structural and electrical properties after epicardial implantation. Biomaterials 2018, 159, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Bargehr, J.; Ong, L.P.; Colzani, M.; Davaapil, H.; Hofsteen, P.; Bhandari, S.; Gambardella, L.; Le Novère, N.; Iyer, D.; Sampaziotis, F.; et al. Epicardial cells derived from human embryonic stem cells augment cardiomyocyte-driven heart regeneration. Nature Biotechnol. 2019, 37, 895–906. [Google Scholar] [CrossRef]
- Błyszczuk, P.; Zuppinger, C.; Costa, A.; Nurzynska, D.; Di Meglio, F.D.; Stellato, M.; Agarkova, I.; Smith, G.L.; Distler, O.; Kania, G. Activated cardiac fibroblasts control contraction of human fibrotic cardiac microtissues by a β-adrenoreceptor-dependent mechanism. Cells 2020, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Miller, K.; Ma, X.; Dewan, S.; Lawrence, N.; Whang, G.; Chung, P.; McCulloch, A.D.; Chen, S. Direct 3D bioprinting of cardiac micro-tissues mimicking native myocardium. Biomaterials 2020, 256, 120204. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, E.; Meraviglia, V.; Campostrini, G.; Cochrane, A.; Cao, X.; Van Helden, R.W.; Garcia, A.K.; Mircea, M.; Kostidis, S.; Davis, R.P.; et al. Human-iPSC-derived cardiac stromal cells enhance maturation in 3D cardiac microtissues and reveal non-cardiomyocyte contributions to heart disease. Cell Stem Cell 2020, 26, 862–879. [Google Scholar] [CrossRef]
- Lee, M.O.; Jung, K.B.; Jo, S.J.; Hyun, S.A.; Moon, K.S.; Seo, J.W.; Kim, S.H.; Son, M.Y. Modelling cardiac fibrosis using three-dimensional cardiac microtissues derived from human embryonic stem cells. J. Biol. Eng. 2019, 13, 15. [Google Scholar] [CrossRef]
- Ergir, E.; Oliver-De La Cruz, J.; Fernandes, S.; Cassani, M.; Niro, F.; Pereira-Sousa, D.; Vrbský, J.; Vinarský, V.; Perestrelo, A.R.; Debellis, D.; et al. Generation and maturation of human iPSC-derived 3D organotypic cardiac microtissues in long-term culture. Sci Rep. 2022, 12, 17409. [Google Scholar] [CrossRef]
- Legant, W.R.; Chen, C.S.; Vogel, V. Force-induced fibronectin assembly and matrix remodeling in a 3D microtissue model of tissue morphogenesis. Integr. Biol. 2012, 4, 1164–1174. [Google Scholar] [CrossRef]
- Song, M.; Kim, J.; Shin, H.; Kim, Y.; Jang, H.; Park, Y.; Kim, S.-J. Development of Magnetic Torque Stimulation (MTS) Utilizing Rotating Uniform Magnetic Field for Mechanical Activation of Cardiac Cells. Nanomaterials 2020, 10, 1684. [Google Scholar] [CrossRef] [PubMed]
- Bracco Gartner, T.C.L.; Wang, Y.; Leiteris, L.; van Adrichem, I.; Marsman, J.; Goumans, M.J.; Bouten, C.V.C.; Sluijter, J.P.G.; den Toonder, J.M.J.; Suyker, W.J.L.; et al. Cyclic strain has antifibrotic effects on the human cardiac fibroblast transcriptome in a human cardiac fibrosis-on-a-chip platform. J. Mech. Behav. Biomed. Mater. 2023, 144, 105980. [Google Scholar] [CrossRef] [PubMed]
- Archer, C.R.; Sargeant, R.; Basak, J.; Pilling, J.; Barnes, J.R.; Pointon, A. Characterization and validation of a human 3D cardiac microtissue for the assessment of changes in car-diac pathology. Sci. Rep. 2018, 8, 10160. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.; Souza, T.; Verheijen, M.C.T.; Gmuender, H.; Selevsek, N.; Schlapbach, R.; Kleinjans, J.; Jennen, D. Translational proteomics analysis of anthracycline-induced cardiotoxicity from cardiac microtissues to human heart biopsies. Front. Genet. 2021, 12, 695625. [Google Scholar] [CrossRef] [PubMed]
- Iravanian, S.; Nabutovsky, Y.; Kong, C.R.; Saha, S.; Bursac, N.; Tung, L. Functional reentry in cultured monolayers of neonatal rat cardiac cells. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H449–H456. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.; Bittihn, P.; Luther, S. Complex restitution behavior and reentry in a cardiac tissue model for neonatal mice. Physiol. Rep. 2017, 5, e13449. [Google Scholar] [CrossRef]
- Kang, M.J.; Koh, G.Y. Differential and dramatic changes of cyclin-dependent kinase activities in cardiomyocytes during the neonatal period. J. Mol. Cell. Cardiol. 1997, 29, 1767–1777. [Google Scholar] [CrossRef]
- Claycomb, W.C.; Lanson, N.A., Jr.; Stallworth, B.S.; Egeland, D.B.; Delcarpio, J.B.; Bahinski, A.; Izzo, N.J., Jr. HL-1 cells: A cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad. Sci. USA 1998, 95, 2979–2984. [Google Scholar] [CrossRef]
- Harley, C.B. Telomere loss: Mitotic clock or genetic time bomb? Mutat. Res. 1991, 256, 271–282. [Google Scholar] [CrossRef]
- Liu, Y.; Mi, Y.; Mueller, T.; Kreibich, S.; Williams, E.G.; Van Drogen, A.; Borel, C.; Frank, M.; Germain, P.L.; Bludau, I.; et al. Multi-omic measurements of heterogeneity in HeLa cells across laboratories. Nat. Biotechnol. 2019, 37, 314–322. [Google Scholar] [CrossRef]
- Agladze, N.N.; Halaidych, O.V.; Tsvelaya, V.A.; Bruegmann, T.; Kilgus, C.; Sasse, P.; Agladze, K.I. Synchronization of excitable cardiac cultures of different origin. Biomater. Sci. 2017, 5, 1777–1785. [Google Scholar] [CrossRef] [PubMed]
- Kudryashova, N.; Nizamieva, A.; Tsvelaya, V.; Panfilov, A.V.; Agladze, K.I. Self-organization of conducting pathways explains electrical wave propagation in cardiac tissues with high fraction of non-conducting cells. PLoS Comput. Biol. 2019, 15, e1006597. [Google Scholar] [CrossRef] [PubMed]
- Kudryashova, N.; Tsvelaya, V.; Agladze, K.; Panfilov, A. Virtual cardiac monolayers for electrical wave propagation. Sci. Rep. 2017, 7, 7887. [Google Scholar] [CrossRef] [PubMed]
- Surani, M.A.; Hayashi, K.; Hajkova, P. Genetic and epigenetic regulators of pluripotency. Cell 2007, 128, 747–762. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef]
- Loh, Y.H.; Wu, Q.; Chew, J.L.; Vega, V.B.; Zhang, W.; Chen, X.; Bourque, G.; George, J.; Leong, B.; Liu, J.; et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat. Genet. 2006, 38, 431–440. [Google Scholar] [CrossRef]
- Tani, H.; Tohyama, S.; Kishino, Y.; Kanazawa, H.; Fukuda, K. Production of Functional Cardiomyocytes and Cardiac Tissue from Human Induced Pluripotent Stem Cells for Regenerative Therapy. J. Mol. Cell. Cardiol. 2022, 164, 83–91. [Google Scholar] [CrossRef]
- Mollova, M.; Bersell, K.; Walsh, S.; Savla, J.; Das, L.T.; Park, S.-Y.; Silberstein, L.E.; Dos Remedios, C.G.; Graham, D.; Colan, S.; et al. Cardiomyocyte Proliferation Contributes to Heart Growth in Young Humans. Proc. Natl. Acad. Sci. USA 2013, 110, 1446–1451. [Google Scholar] [CrossRef]
- Tani, H.; Tohyama, S. Human engineered heart tissue models for disease modeling and drug discovery. Front. Cell Dev. Biol. 2022, 10, 855763. [Google Scholar] [CrossRef]
- Hom, J.R.; Quintanilla, R.A.; Hoffman, D.L.; de Mesy Bentley, K.L.; Molkentin, J.D.; Sheu, S.S.; Porter, G.A. The Permeability Transition Pore Controls Cardiac Mitochondrial Maturation and Myocyte Differentiation. Dev. Cell 2011, 21, 469–478. [Google Scholar] [CrossRef]
- Mauritz, C.; Schwanke, K.; Reppel, M.; Neef, S.; Katsirntaki, K.; Maier, L.S.; Nguemo, F.; Menke, S.; Haustein, M.; Hescheler, J.; et al. Generation of functional murine cardiac myocytes from induced pluripotent stem cells. Circulation 2008, 118, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tohyama, S.; Murata, M.; Nomura, F.; Kaneko, T.; Chen, H.; Hattori, F.; Egashira, T.; Seki, T.; Ohno, Y.; et al. In vitro pharmacologic testing using human induced pluripotent stem cell-derived cardiomyocytes. Biochem. Biophys. Res. Commun. 2009, 385, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.J.; Martinez-Fernandez, A.; Yamada, S.; Perez-Terzic, C.; Ikeda, Y.; Terzic, A. Repair of acute myocardial infarction by human stemness factors induced pluripotent stem cells. Circulation 2009, 120, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.W.; Ripplinger, C.M.; Van Der Meer, P.; Sheehy, S.P.; Domian, I.; Chien, K.R.; Parker, K.K. Functional differences in engineered myocardium from embryonic stem cell-derived versus neonatal cardiomyocytes. Stem Cell Rep. 2013, 1, 387–396. [Google Scholar] [CrossRef]
- Smith, C. Differentiation of Human Embryonic Stem Cells to Cardiomyocytes: Role of Coculture with Visceral Endoderm-Like Cells. Circulation 2003, 107, 2733–2740. [Google Scholar]
- Mummery, C.L.; Zhang, J.; Ng, E.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of Human ES and iPS Cells to Cardiomyocytes: A Methods Overview. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef]
- Slotvitsky, M.M.; Tsvelaya, V.A.; Podgurskaya, A.D.; Agladze, K.I. Formation of an electrical coupling between differentiating cardiomyocytes. Sci. Rep. 2020, 10, 7774. [Google Scholar] [CrossRef]
- Chen, A.; Lee, E.; Tu, R.; Santiago, K.; Grosberg, A.; Fowlkes, C.; Khine, M. Integrated platform for functional monitoring of biomimetic heart sheets derived from human pluripotent stem cells. Biomaterials 2014, 35, 675–683. [Google Scholar] [CrossRef]
- Ribas, J.; Sadeghi, H.; Manbachi, A.; Leijten, J.; Brinegar, K.; Zhang, Y.S.; Ferreira, L.; Khademhosseini, A. Cardiovascular organ-on-a-chip platforms for drug discovery and development. Appl. Vitr. Toxicol. 2016, 2, 82–96. [Google Scholar] [CrossRef]
- Kurokawa, Y.K.; George, S.C. Tissue engineering the cardiac microenvironment: Multicellular microphysiological systems for drug screening. Adv. Drug Deliv. Rev. 2016, 96, 225–233. [Google Scholar] [CrossRef]
- Gintant, G. An Evaluation of hERG Current Assay Performance: Translating Preclinical Safety Studies to Clinical QT Prolongation. Pharmacol. Ther. 2011, 129, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Pourrier, M.; David, F. The emergence of human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) as a platform to model arrhythmogenic diseases. Int. J. Mol. Sci. 2020, 21, 657. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Papoian, T. Moving beyond the Comprehensive In Vitro Proarrhythmia Assay: Use of Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes to Assess Contractile Effects Associated with Drug-Induced Structural Cardiotoxicity. J. Appl. Toxicol. 2018, 38, 1166–1176. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, E.G.; Liang, P.; Lan, F.; Sanchez-Freire, V.; Simmons, C.; Gong, T.; Sharma, A.; Burridge, P.W.; Patlolla, B.; Lee, A.S.; et al. Screening drug-induced arrhythmia using human induced pluripotent stem cell–derived cardiomyocytes and low-impedance microelectrode arrays. Circulation 2013, 128 (Suppl. S1), S3–S13. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y.; Yamazaki, D.; Osada, T.; Yoshinaga, T.; Sawada, K. Development of Torsadogenic Risk Assessment Using Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes: Japan iPS Cardiac Safety Assessment (JiCSA) Update. J. Pharmacol. Sci. 2018, 138, 233–239. [Google Scholar] [CrossRef]
- Sakamoto, K.; Sakatoku, K.; Sugimoto, S.; Iwasaki, N.; Sano, Y.; Yamaguchi, M.; Kurokawa, J. Continued Exposure of Anti-cancer Drugs to Human iPS Cell-Derived Cardiomyocytes Can Unmask Their Cardiotoxic Effects. J. Pharmacol. Sci. 2019, 140, 345–349. [Google Scholar] [CrossRef]
- Podgurskaya, A.D.; Tsvelaya, V.A.; Slotvitsky, M.M.; Dementyeva, E.V.; Valetdinova, K.R.; Agladze, K.I. The use of iPSC-derived cardiomyocytes and optical mapping for erythromycin arrhythmogenicity testing. Cardiovasc. Toxicol. 2019, 19, 518–528. [Google Scholar] [CrossRef]
- Podgurskaya, A.D.; Slotvitsky, M.M.; Tsvelaya, V.A.; Frolova, S.R.; Romanova, S.G.; Balashov, V.A.; Agladze, K.I. Cyclophosphamide arrhythmogenicitytesting using human-induced pluripotent stem cell-derived cardiomyocytes. Sci. Rep. 2021, 11, 2336. [Google Scholar] [CrossRef]
- Jorba, I.; Mostert, D.; Hermans, L.H.L.; van der Pol, A.; Kurniawan, N.A.; Bouten, C.V.C. In vitro methods to model cardiac mechanobiology in health and disease. Tissue Eng. Part C Methods 2021, 27, 139–151. [Google Scholar] [CrossRef]
- Pasqualini, F.S.; Agarwal, A.; O’Connor, B.B.; Liu, Q.; Sheehy, S.P.; Parker, K.K. Traction force microscopy of engineered cardiac tissues. PLoS ONE 2018, 13, e0194706. [Google Scholar] [CrossRef]
- McCain, M.L.; Yuan, H.; Pasqualini, F.S.; Campbell, P.H.; Parker, K.K. Matrix elasticity regulates the optimal cardiac myocyte shape for contractility. Am. J. Physiol. Circ. Physiol. 2014, 306, H1525–H1539. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.J.S.; Ang, Y.-S.; Fu, J.-D.; Rivas, R.N.; Mohamed, T.M.; Higgs, G.C.; Srivastava, D.; Pruitt, B.L. Contractility of single cardiomyocytes differentiated from pluripotent stem cells depends on physiological shape and substrate stiffness. Proc. Natl. Acad. Sci. USA 2015, 112, 12705–12710. [Google Scholar] [CrossRef]
- Ribeiro, M.C.; Tertoolen, L.G.; Guadix, J.A.; Bellin, M.; Kosmidis, G.; D’Aniello, C.; Monshouwer-Kloots, J.; Goumans, M.J.; Wang, Y.L.; Feinberg, A.W.; et al. Functional maturation of human pluripotent stem cell derived cardiomyocytes in vitro—Correlation between contraction force and electrophysiology. Biomaterials 2015, 51, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Broughton, K.M.; Russell, B. Cardiomyocyte subdomain contractility arising from microenvironmental stiffness and topography. Biomech. Model. Mechanobiol. 2015, 14, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Lin, I.-K.; Ou, K.-S.; Liao, Y.-M.; Liu, Y.; Chen, K.-S.; Zhang, X. Viscoelastic characterization and modeling of polymer transducers for biological applications. J. Microelectromech. Syst. 2009, 18, 1087–1099. [Google Scholar]
- Corbin, E.A.; Vite, A.; Peyster, E.G.; Bhoopalam, M.; Brandimarto, J.; Wang, X.; Bennett, A.I.; Clark, A.T.; Cheng, X.; Turner, K.T.; et al. Tunable and reversible substrate stiffness reveals a dynamic mechanosensitivity of cardiomyocytes. ACS Appl. Mater. Interfaces 2019, 11, 20603–20614. [Google Scholar] [CrossRef]
- Engler, A.J.; Carag-Krieger, C.; Johnson, C.P.; Raab, M.; Tang, H.Y.; Speicher, D.W.; Sanger, J.W.; Sanger, J.M.; Discher, D.E. Embryonic cardiomyocytes beat best on a matrix with heart-like elasticity: Scar-like rigidity inhibits beating. J. Cell Sci. 2008, 121, 3794–3802. [Google Scholar] [CrossRef]
- Herron, T.J.; Da Rocha, A.M.; Campbell, K.F.; Ponce-Balbuena, D.; Willis, B.C.; Guerrero-Serna, G.; Liu, Q.; Klos, M.; Musa, H.; Zarzoso, M.; et al. Extracellular matrix-mediated maturation of human pluripotent stem cell-derived cardiac monolayer structure and electrophysiological function. Circ. Arrhythmia Electrophysiol. 2016, 9, e003638. [Google Scholar] [CrossRef]
- Ribeiro, M.C.; Slaats, R.H.; Schwach, V.; Rivera-Arbelaez, J.M.; Tertoolen, L.G.J.; van Meer, B.J.; Molenaar, R.; Mummery, C.L.; Claessens, M.M.A.E.; Passier, R. A cardiomyocyte show of force: A fluorescent alpha-actinin reporter line sheds light on human cardiomyocyte contractility versus substrate stiffness. J. Mol. Cell. Cardiol. 2020, 141, 54–65. [Google Scholar] [CrossRef]
- Sheehy, S.P.; Grosberg, A.; Qin, P.; Behm, D.J.; Ferrier, J.P.; Eagleson, M.A.; Nesmith, A.P.; Krull, D.; Falls, J.G.; Campbell, P.H.; et al. Toward improved myocardial maturity in an organ-on-chip platform with immature cardiac myocytes. Exp. Biol. Med. 2017, 242, 1643–1656. [Google Scholar] [CrossRef]
- Ariyasinghe, N.R.; Reck, C.H.; Viscio, A.A.; Petersen, A.P.; Lyra-Leite, D.M.; Cho, N.; McCain, M.L. Engineering micromyocardium to delineate cellular and extracellular regulation of myocardial tissue contractility. Integr. Biol. 2017, 9, 730–741. [Google Scholar] [CrossRef]
- Gopalan, S.M.; Flaim, C.; Bhatia, S.N.; Hoshijima, M.; Knoell, R.; Chien, K.R.; Omens, J.H.; McCulloch, A.D. Anisotropic stretch-induced hypertrophy in neonatal ventricular myocytes micropatterned on deformable elastomers. Biotechnol. Bioeng. 2003, 81, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Martewicz, S.; Luni, C.; Serena, E.; Pavan, P.; Chen, H.V.; Rampazzo, A.; Elvassore, N. Transcriptomic characterization of a human in vitro model of arrhythmogenic cardiomyopathy under topological and mechanical stimuli. Ann. Biomed. Eng. 2019, 47, 852–865. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, S.; Fujihara, K.; Teo, W.-E.; Lim, T.-C.; Ma, Z. An Introduction to Electrospinning and Nanofibers; World Scientific: Singapore, 2005; ISBN 978-981-256-415-3. [Google Scholar]
- Sell, S.A.; McClure, M.J.; Garg, K.; Wolfe, P.S.; Bowlin, G.L. Electrospinning of collagen/biopolymers for regenerative medicine and cardiovascular tissue engineering. Adv. Drug Deliver. Rev. 2009, 61, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Mondrinos, M.J.; Gandhi, M.R.; Ko, F.K.; Weiss, A.S.; Lelkes, P.I. Electrospun protein fibers as matrices for tissue engineering. Biomaterials 2005, 26, 5999–6008. [Google Scholar] [CrossRef]
- Zong, X.; Bien, H.; Chung, C.-Y.; Yin, L.; Fang, D.; Hsiao, B.S.; Chu, B.; Entcheva, E. Electrospun fine-textured scaffolds for heart tissue constructs. Biomaterials 2005, 26, 5330–5338. [Google Scholar] [CrossRef]
- Zhong, J.; Zhang, H.; Yan, J.; Gong, X. Effect of nanofiber orientation of electrospun nanofibrous scaffolds on cell growth and elastin expression of muscle cells. Colloids Surf. B Biointerfaces 2015, 136, 772–778. [Google Scholar] [CrossRef]
- Tomecka, E.; Wojasinski, M.; Jastrzebska, E.; Chudy, M.; Ciach, T.; Brzozka, Z. Poly(l-lactic acid) and polyurethane nanofibers fabricated by solution blow spinning as potential substrates for cardiac cell culture. Mater. Sci. Eng. C 2017, 75, 305–316. [Google Scholar] [CrossRef]
- Balashov, V.; Efimov, A.; Agapova, O.; Pogorelov, A.; Agapov, I.; Agladze, K. High resolution 3D microscopy study of cardiomyocytes on polymer scaffold nanofibers reveals formation of unusual sheathed structure. Acta Biomater. 2018, 68, 214–222. [Google Scholar] [CrossRef]
- Teplenin, A.; Krasheninnikova, A.; Agladze, N.; Sidoruk, K.; Agapova, O.; Agapov, I.; Bogush, V.; Agladze, K. Functional Analysis of the Engineered Cardiac Tissue Grown on Recombinant Spidroin Fiber Meshes. PLoS ONE 2015, 10, e0121155. [Google Scholar] [CrossRef]
- Sayed, M.M.; Mousa, H.M.; El-Aassar, M.R.; El-Deeb, N.M.; Ghazaly, N.M.; Dewidar, M.M.; Abdal-hay, A. Enhancing mechanical and biodegradation properties of polyvinyl alcohol/silk fibroin nanofibers composite patches for Cardiac Tissue Engineering. Mater. Lett. 2019, 255, 126510. [Google Scholar] [CrossRef]
- Safonova, L.; Bobrova, M.; Efimov, A.; Lyundup, A.; Agapova, O.; Agapov, I. A Comparative Analysis of the Structure and Biological Properties of Films and Microfibrous Scaffolds Based on Silk Fibroin. Pharmaceutics 2021, 13, 1561. [Google Scholar] [CrossRef] [PubMed]
- Safonova, L.; Bobrova, M.; Efimov, A.; Davydova, L.; Tenchurin, T.; Bogush, V.; Agapova, O.; Agapov, I. Silk Fibroin/Spidroin Electrospun Scaffolds for Full-Thickness Skin Wound Healing in Rats. Pharmaceutics 2021, 13, 1704. [Google Scholar] [CrossRef] [PubMed]
- Moisenovich, M.M.; Pustovalova, O.; Shackelford, J.; Vasiljeva, T.V.; Druzhinina, T.V.; Kamenchuk, Y.A.; Guzeev, V.V.; Sokolova, O.S.; Bogush, V.G.; Debabov, V.G.; et al. Tissue regeneration in vivo within recombinant spidroin 1 scaffolds. Biomaterials 2012, 33, 3887–3898. [Google Scholar] [CrossRef]
- Baklaushev, V.P.; Bogush, V.G.; Kalsin, V.A.; Sovetnikov, N.N.; Samoilova, E.M.; Revkova, V.A.; Sidoruk, K.V.; Konoplyannikov, M.A.; Timashev, P.S.; Kotova, S.L.; et al. Tissue Engineered Neural Constructs Composed of Neural Precursor Cells, Recombinant Spidroin and PRP for Neural Tissue Regeneration. Sci. Rep. 2019, 9, 3161. [Google Scholar] [CrossRef]
- Patra, C.; Talukdar, S.; Novoyatleva, T.; Velagala, S.R.; Muhlfeld, C.; Kundu, B.; Kundu, S.C.; Engel, F.B. Silk protein fibroin from Antheraea mylitta for cardiac tissue engineering. Biomaterials 2012, 33, 2673–2680. [Google Scholar] [CrossRef]
- Slotvitsky, M.; Berezhnoy, A.; Scherbina, S.; Rimskaya, B.; Tsvelaya, V.; Balashov, V.; Efimov, A.E.; Agapov, I.; Agladze, K. Polymer Kernels as Compact Carriers for Suspended Cardiomyocytes. Micromachines 2023, 14, 51. [Google Scholar] [CrossRef]
- Aitova, A.; Scherbina, S.; Berezhnoy, A.; Slotvitsky, M.; Tsvelaya, V.; Sergeeva, T.; Turchaninova, E.; Rybkina, E.; Bakumenko, S.; Sidorov, I.; et al. Novel Molecular Vehicle-Based Approach for Cardiac Cell Transplantation Leads to Rapid Electromechanical Graft–Host Coupling. Int. J. Mol. Sci. 2023, 24, 10406. [Google Scholar] [CrossRef]
- Stachewicz, U.; Qiao, T.; Rawlinson, S.C.F.; Almeida, F.V.; Li, W.Q.; Cattell, M.; Barber, A.H. 3D imaging of cell interactions with electrospun PLGA nanofiber membranes for bone regeneration. Acta Biomater. 2015, 27, 88–100. [Google Scholar] [CrossRef]
- Mochalov, K.E.; Chistyakov, A.A.; Solovyeva, D.O.; Mezin, A.V.; Oleinikov, V.A.; Vaskan, I.S.; Molinari, M.; Agapov, I.I.; Nabiev, I.; Efimov, A.E. An instrumental approach to combining confocal microspectroscopy and 3D scanning probe nanotomography. Ultramicroscopy 2017, 182, 118–123. [Google Scholar] [CrossRef]
- Efimov, A.E.; Agapov, I.I.; Agapova, O.I.; Oleinikov, V.A.; Mezin, A.V.; Molinari, M.; Nabiev, I.; Mochalov, K.E. A novel design of a scanning probe microscope integrated with an ultramicrotome for serial block-face nanotomography. Rev. Sci. Instrum. 2017, 88, 023701. [Google Scholar] [CrossRef] [PubMed]
- Efimov, A.E.; Agapova, O.I.; Safonova, L.A.; Bobrova, M.M.; Parfenov, V.A.; Koudan, E.V.; Pereira, F.D.A.S.; Bulanova, E.A.; Mironov, V.A.; Agapov, I.I. 3D scanning probe nanotomography of tissue spheroid fibroblasts interacting with electrospun polyurethane scaffold. Express Polym. Lett. 2019, 13, 632–641. [Google Scholar] [CrossRef]
- Wang, L.; Wu, Y.; Hu, T.; Guo, B.; Ma, P.X. Electrospun conductive nanofibrous scaffolds for engineering cardiac tissue and 3D bioactuators. Acta Biomater. 2017, 59, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.V.; Jalife, J. Rotors and the dynamics of cardiac fibrillation. Circ. Res. 2013, 112, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Starmer, C.F.; Biktashev, V.N.; Romashko, D.N.; Stepanov, M.R.; Makarova, O.N.; Krinsky, V.I. Vulnerability in an excitable medium: Analytical and numerical studies of initiating unidirectional propagation. Biophys. J. 1993, 65, 1775–1787. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Arrhythmias or Disease | Dimension | Basic Methods of Creating Heart Biomimetic | Cell Type and Gene Mutations | Substrate | Structural Features | Article |
|---|---|---|---|---|---|---|
| Ventricular tachycardias and Postmyocardial infarction | 2D | Coculturing, non-adhesive, and adhesive micropatterning | Neonatal rat ventricular myocytes | fibronectin | Model of the infarct border zone with human skeletal myotubes, nonuniform anisotropic architecture | Chang, Marvin G. et al. “Spiral waves and reentry dynamics in an in vitro model of the healed infarct border zone.” Circulation research 105.11 (2009): 1062–1071. [48] |
| 3D | Coculturing | hiPSCs (836B3 line18), mesenchymal cell | Gelatine; 48-multiwell UpCell | 3D cardiac tissue sheets with two cell types | Kawatou, Masahide et al. “Modelling Torsade de Pointes arrhythmias in vitro in 3D human iPS cell-engineered heart tissue.” Nature communications 8.1 (2017): 1078. [49] | |
| 3D | Molds with ECM pre-gel | hiPSC (from patient with LQTS 2 and CPVT2) | Matrigel | 3D cardiac rings on passive-stretcher devices | Goldfracht, Idit et al. “Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications.” Acta biomaterialia 92 (2019): 145–159. [52] | |
| 2D | Glucose-free and hypoxic conditions | Neonatal rat myocytes | Gelatine | Model of Ischemia/Reperfusion Injury, isotropic architecture | Sugiyama, A.; Shimizu, Y.; Okada, M.; Otani, K.; Yamawaki, H. Preventive Effect of Canstatin against Ventricular Arrhythmia Induced by Ischemia/Reperfusion Injury: A Pilot Study. Int. J. Mol. Sci. 2021, 22, 1004. https://doi.org/10.3390/ijms22031004 [53] | |
| 3D | Polyacrylamide hydrogels with variable stiffness | iPSC: line TaP4 | Gelatine | Engineered heart tissues (EHTs); different matrixes | Heras-Bautista, Carlos O. et al. “Cardiomyocytes facing fibrotic conditions re-express extracellular matrix transcripts.” Acta biomaterialia 89 (2019): 180–192. [54] | |
| 2D | Coculturing (hiPSC-aCMs and hiPSC-vCMs) | hiPSCs: WiCell and IMR90-1 | Matrigel | Non-structured monolayers | Gunawan, Marvin G. et al. “Drug screening platform using human induced pluripotent stem cell-derived atrial cardiomyocytes and optical mapping.” Stem Cells Translational Medicine 10.1 (2021): 68–82. [55] | |
| Brugada syndrome | 2D | Brugada syndrome associated line investigation | iPSCs lines (BrS1-iPSCs, Ctrl1-iPSCs, Ctrl2-iPSCs, and BrS2-iPSCs) | Geltrex | Non-structured monolayers | Li Wener, W., Stauske, M., Luo, X., Wagner, S., Vollrath, M., Mehnert, C.S. et al. (2020). Disease Phenotypes and Mechanisms of iPSC-Derived Cardiomyocytes from Brugada Syndrome Patients with a Loss-Of-Function SCN5A Mutation. Front. Cell Dev. Biol. 8, 592893. Epub 2020/11/17. doi:10.3389/fcell.2020.592893 [51] |
| Brugada syndrome associated line investigation | hiPSCs: he BrS cell lines isBrSd1.9 (GOEi098-A.9), isBrSd1.12 (GOEi098-A.12), and isBrSd1.23 (GOEi098-A.23) | Matrigel | Non-structured monolayers | El-Battrawy, Ibrahim et al. “A cellular model of Brugada syndrome with SCN10A variants using human-induced pluripotent stem cell-derived cardiomyocytes.” EP Europace 21.9 (2019): 1410–1421. [56] | ||
| Short QT interval syndrome | 2D | SQTS1 associated line investigation | hiPSCs: with N588K mutation in the KCNH2 gene and healthy hiPSC-CMs | Matrigel | Non-structured monolayers | Shinnawi, Rami et al. “Modeling reentry in the short QT syndrome with human-induced pluripotent stem cell–derived cardiac cell sheets.” Journal of the American College of Cardiology 73.18 (2019): 2310–2324. [57] |
| 2D | Embryoid body (EB)-based differentiating technology; SQTS1 associated line investigation | hiPSCs: SQTS (N588K KCNH2 mutation), isogenic control, and healthy control | Non-adhesive v-shaped plate; Matrigel | Atrial cell sheets (hiPSC-ACSs), non-structured monolayers | Shiti, Assad et al. “Utilizing human induced pluripotent stem cells to study atrial arrhythmias in the short QT syndrome.” Journal of Molecular and Cellular Cardiology 183 (2023): 42–53. [58] | |
| 2D | SQTS1 associated line investigation | hiPSCs: with mutation (N588K) in KCNH2 and two healthy controls | Matrigel | Cell sheets (hiPSC-ACSs), non-structured monolayers | El-Battrawy, I., Lan, H., Cyganek, L., Zhao, Z., Li, X., Buljubasic, F. et al. (2018). Modeling Short QT Syndrome Using Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes. J. Am. Heart Assoc. 7, 7. Epub 2018/03/27. doi:10.1161/JAHA.117.007394 [59] | |
| Long QT interval syndrome | 3D | Decellularization for matrix creation | hiPSCs: line with LQTS2 and line with polymorphic ventricular tachycardia type 2 (CPVT2) | ECM pre-gel solution | ECM-derived engineered heart tissues (ECM-EHTs); anisotropic architecture | Giacomelli E. et al. “Cardiac microtissues from human pluripotent stem cells recapitulate the phenotype of long-QT syndrome.” Biochemical and Biophysical Research Communications 572 (2021): 118–124. [60] |
| 3D | Coculturing (three cell types); embryoid body (EB)-based culturing | hiPSC: wild-type (WT) hiPSC line LUMC0020iCTRL-06 (female, LUMCi028-A), and long-QT syndrome type 1 (LQT1) hiPSC line LUMC0021iKCNQ-30 (female, LUMCi039-A) carrying the KCNQ1-R594Q | Non-adhesive v-shaped plates; Matrigel-coated multielectrode array (MEA) | Cardiac microtissues | Goldfracht, I., Efraim, Y., Shinnawi, R., Kovalev, E., Huber, I., Gepstein, A., ... & Gepstein, L. (2019). Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta biomaterialia, 92, 145–159. [52] | |
| Hypertrophic cardiomyopathy (HCM) | 2D | HCM-associated line investigation | hiPSC: wild-type (MYH7WT/WT), heterozygous (MYH7WT/MUT) and homozygous (MYH7MUT/MUT) | Matrigel | Monolayers | Bhagwan, Jamie R. et al. “Isogenic models of hypertrophic cardiomyopathy unveil differential phenotypes and mechanism-driven therapeutics.” Journal of molecular and cellular cardiology 145 (2020): 43–53. [61] |
| 2D | HCM-associated line investigation | hiPSC: TNNI3R21C/+, TNNT2R92Q/+ and MYH7R403Q/+ | Matrigel | Monolayers | Margara, Francesca et al. “Mechanism based therapies enable personalised treatment of hypertrophic cardiomyopathy.” Scientific Reports 12.1 (2022): 22501. [62] | |
| Duchenne muscular dystrophy (DMD) | 3D | Embryoid body-based differentiation | iPSC: with DMD and control line | Non-adhesive plate | Non-structured organoids | Lin, Bo et al. “Modeling and study of the mechanism of dilated cardiomyopathy using induced pluripotent stem cells derived from individuals with Duchenne muscular dystrophy.” Disease models & mechanisms 8.5 (2015): 457–466. [51] |
| Arrhythmogenic cardiomyopathy (ACM) | 2D | ACM-associated line investigation; coculturing | hiPSC: human PKP2 c.2013delC, PKP2 c.1849C>T, and control lines | Geltrex | Monolayers, spontaneous fibro-adipose cell differentiation | Kohela, A.v.K.S., Moens, T., Wehrens, M., Molenaar, B., Boogerd, C.J., Monshouwer-Kloots, J. et al. (2021). Epicardial Differentiation Drives Fibro-Fatty Remodeling in Arrhythmogenic Cardiomyopathy. Sci. Transl Med. 2021, 13. doi:10.1126/scitranslmed.abf2750 [63] |
| 3D | Generating EHT by PDMS; monolayer-based differentiation; and ACM-associated line investigation | ESC; hiPSC | PDMS strips; Pluronic F-127; collagen I; Matrigel | Micropatterning; engineered heart tissue (EHT) | Bliley, Jacqueline M. et al. “Dynamic loading of human engineered heart tissue enhances contractile function and drives a desmosome-linked disease phenotype.” Science translational medicine 13.603 (2021): eabd1817. [64] |
| Basic Heart Tissue Model | Purpose of Cardiac Tissue Modeling | Dimension | Cell Types or Cell Source | Substrate Types | Modeling of Structural Features | Article Examples |
|---|---|---|---|---|---|---|
| Organoids and organs-on-chip | Disease modeling and drug testing applications | 3D | hiPSC lines and LQTS2 and CPVT hiPSC lines for ventricular-like cardiomyocytes | Decellularized porcine cardiac tissues (ECM); solubilized porcine cardiac ECM (pcECM) hydrogel | Non-structured ECM hydrogel | Goldfracht, I., Efraim, Y., Shinnawi, R., Kovalev, E., Huber, I., Gepstein, A., ... & Gepstein, L. (2019). Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta biomaterialia, 92, 145–159. [52] |
| 3D | hESC line for ventricular and atrial cardiomyocytes | Decellularized porcine cardiac tissues (ECM); solubilized porcine cardiac ECM (pcECM) hydrogel | Non-structured ECM hydrogel | Goldfracht, Idit et al. “Generating ring-shaped engineered heart tissues from ventricular and atrial human pluripotent stem cell-derived cardiomyocytes.” Nature communications 11.1 (2020): 75. [65] | ||
| 3D | hiPSC-CMs (iCell Cardiomyocytes, Cellular Dynamics) | Non-adhesive agarose hydrogel molds | Non-structured organoids (50% hiPSC-CMs and 50% non-myocyte) | Richards D.J. et al. “Human cardiac organoids for the modelling of myocardial infarction and drug cardiotoxicity.” Nature Biomedical Engineering 4.4 (2020): 446–462. [66] | ||
| 3D | hiPSC-CMs and NRVMs | Polycaprolactone (PCL)/gelatin nanofibers | Structured ventricle chamber, anisotropic myocardial tissue | MacQueen, Luke A. et al. “Addendum: A tissue-engineered scale model of the heart ventricle.” Nature Biomedical Engineering 6.11 (2022): 1318–1318. [67] | ||
| The vascularization in vitro problem | 3D | hiPSC-derived cardiomyocytes; rat primary cardiac microvascular endothelial cells (CECs) | Organoid culturing in microwells mixed in Matrigel | Stress-structured vascularization | Ghosheh, Mohammad et al. “Electro-metabolic coupling in multi-chambered vascularized human cardiac organoids.” Nature biomedical engineering (2023): 1–21. [68] | |
| 3D | hiPSC: hiPSCs from human foreskin fibroblasts | Ultra-low attachment Petri dish; 10% Matrigel in medium | Non-structured heart organoids, vascularized in vivo (after injection) | Lee, Seul-Gi et al. “Generation of human iPSCs derived heart organoids structurally and functionally similar to heart.” Biomaterials 290 (2022): 121860. [69] | ||
| 3D | hiPSC: H9 and H13 cell lines | Matrigel; U-bottom culture plate | Non-structured embryoids with vascularisation | Liang, Po-Yu et al. “Wnt signaling directs human pluripotent stem cells into vascularized cardiac organoids with chamber-like structures.” Frontiers in Bioengineering and Biotechnology 10 (2022): 1059243. [70] | ||
| Investigation of cell maturation in vitro | 3D | hiPSC: WTC hiPSCs (from B.R. Conklin lab, Gladstone Institute of Cardiovascular Disease; also available from the Coriell Institute NIGMS Human Genetic Cell Repository, cat. no. GM25256) | Micropatterned PEG substrate: Photomasks; SU8 patterns on silicon wafers; PDMS; PEG nonfouling thin polymer film; Covering: hESC-qualified Matrigel | Well-structured micropatterns: small patterned organoids in different forms (circles, triangles, and squares) | Hoang, Plansky et al. “Generation of spatial-patterned early developing cardiac organoids using human pluripotent stem cells.” Nature protocols 13.4 (2018): 723–737. [71] | |
| 3D | Human cardiac microtissues (3D InSightTM Human Cardiac Microtissues from InSphero) and human biopsies | AggreWell400 plates, ultra-low attachment 96-well plates, hydrogel | Non-structured cocultured cardioids | Hofbauer, Pablo et al. “Cardioids reveal self-organizing principles of human cardiogenesis.” Cell 184.12 (2021): 3299–3317. [72] | ||
| 3D | hESC: HES3 NKX2.5–eGFP, HSC_ADCF_SeV-iPS, HES3 MIXL1–GFP, and HES3 NKX2.5–eGFP/eGFP (NKX2.5-KO) cell lines | U-shaped ultra-low attachment 96-well plate, Matrigel droplet per each well, and Geltrex (growth factor-reduced Matrigel preparation) or collagen I for embedding | High-structured heart-forming organoids (HFO) with three layers: IC (inner core with mesendoderm and cardiac-like cells), ML (NKX2.5–eGFP-positive myocardial layer), and OL (outer layer with mesendoderm) | Drakhlis, Lika et al. “Human heart-forming organoids recapitulate early heart and foregut development.” Nature Biotechnology 39.6 (2021): 737–746. [73] | ||
| 3D | ESC: atrial tissue (chicken embryos) | Polyacrylamide (PA) gels | Substrate micropatterning, Microstructured hydrogels | Thomas, Kandace et al. “Adherens junction engagement regulates functional patterning of the cardiac pacemaker cell lineage.” Developmental cell 56.10 (2021): 1498–1511. [74] | ||
| Investigation of mechanotransduction-related events in vitro | 3D | MSC: immortalized adipose-derived mesenchymal stem cells | Poly-L-lysine-g-poly and light-responsive substrates: Poly-Disperse Red 1-methacrylate (pDR1m) | Substrate micropatterning | Cimmino, C., Netti, P.A., & Ventre, M. (2022). A switchable light-responsive azopolymer conjugating protein micropatterns with topography for mechanobiological studies. Frontiers in Bioengineering and Biotechnology, 10, 933410. [75] | |
| 2D, 3D | hESC: line CCTL14 | Agarose gel | Non-structured embryoid bodies (“beating clusters”) | Caluori, Guido et al. “Simultaneous study of mechanobiology and calcium dynamics on hESC-derived cardiomyocytes clusters.” Journal of Molecular Recognition 32.2 (2019): e2760. [76] | ||
| 3D | hESC: line CCTL14 and hiPSC line reprogrammed from fibroblasts of a patient affected by Duchenne muscular dystrophy (DMD) | Laminin and fibronectin | Non-structured embryoid bodies (“beating clusters”) | Caluori, Guido et al. “Non-invasive electromechanical cell-based biosensors for improved investigation of 3D cardiac models.” Biosensors and Bioelectronics 124 (2019): 129–135. [77] | ||
| Engineered heart tissue (EHT) | Disease modeling and drug testing applications | 2D, 3D | hiPSC: Ctrl cell lines (C25, ERC18, and ERC1) | Geltrex for monolayer cultures, fibrinogen and thrombin for EHT | Non-structured isotropic EHT between elastic silicone posts for contractile detection | Lemme, Marta et al. “Atrial-like engineered heart tissue: an in vitro model of the human atrium.” Stem cell reports 11.6 (2018): 1378–1390. [78] |
| 2D | hESC: human embryonic cell line HES2 (ESI, NIH code ES02) | Polydimethylsiloxane (PDMS) mold, microgrooving by photolithography | Substrate micropatterning, anisotropic sheet | Shum, Angie MY et al. “A micropatterned human pluripotent stem cell-based ventricular cardiac anisotropic sheet for visualizing drug-induced arrhythmogenicity.” Advanced Materials 29.1 (2017): 1602448. [79] | ||
| 3D | hESC: lines HES2 (female) and HES3-NKX2-5eGFP/w (female); hiPSC: hiPSC line BJ1D (male), C2A hiPSC derived cardiomyocytes, and iCell and iCell2 cardiomyocytes; patient biopsies | Collagen hydrogel, negative polydimethylsiloxane (PDMS) master mold, and PDMS master for microwell creation | High-structured micropatterning cell sheet with two ends: atrial and ventricle with poly(octamethylene maleate-(anhydride) citrate) (POMaC) polymer wires on each end | Zhao, Yimu et al. “A platform for generation of chamber-specific cardiac tissues and disease modeling.” Cell 176.4 (2019): 913–927. [80] | ||
| 2D | iPSC: iSMA6L cell line | Geltrex | Isotropic cell sheets with standard sharp obstacle | Slotvitsky, M., Tsvelaya, V., Frolova, S., Dementyeva, E., & Agladze, K. (2019). Arrhythmogenicity test based on a hu-man-induced pluripotent stem cell (iPSC)-derived cardiomyocyte layer. Toxicological Sciences, 168(1), 70–77. [81] | ||
| Investigation of cell maturation in vitro | 3D | hiPSC: line DiPS 1016SevA (from skin fibroblasts) and line Huv-iPS4F1 (from human umbilical cord vein endothelial cells (HUVECs)) | Hydrogel GelMA and PEG-RGD | Non-structured monolayers | Acun, Aylin, Trung Dung Nguyen, and Pinar Zorlutuna. “In vitro aged, hiPSC-origin engineered heart tissue models with age-dependent functional deterioration to study myocardial infarction.” Acta biomaterialia 94 (2019): 372–391. [82] | |
| 2D | NRVM cells | Polydimethysiloxane (PDMS, Dow Corning) tissue molds; PTFE template; fibrinogen, Matrigel | Structured cell/hydrogel tissue patches | Jackman, Christopher P. et al. “Engineered cardiac tissue patch maintains structural and electrical properties after epicardial implantation.” Biomaterials 159 (2018): 48–58. [83] | ||
| 3D | hESC: H9, WiCell lines; hESC-EPIs and hESC-derived CMs | Polydimethylsiloxane (PDMS); Collagen gel with Collagen I Rat Protein, Geltrex | Non-structured cocultural ETH | Bargehr, Johannes et al. “Epicardial cells derived from human embryonic stem cells augment cardiomyocyte-driven heart regeneration.” Nature biotechnology 37.8 (2019): 895–906. [84] | ||
| Microtissues | Disease modeling and drug testing applications | 3D | iPSC-derived cardiomyocytes, Cellular Dynamics International, fetal human cardiac fibroblasts from Sigma (Cell Applications) | GravityTRAP plates | Microtissues with fibrosis modeling; cell cocultural aggregates (CM microtissues with fibroblasts) | Błyszczuk, Przemysław et al. “Activated cardiac fibroblasts control contraction of human fibrotic cardiac microtissues by a β-adrenoreceptor-dependent mechanism.” Cells 9.5 (2020): 1270. [85] |
| 3D | Neonatal mouse cardiomyocytes (NMVCMs) | PDMS spacers with methacrylated gelatin (GelMA); hyaluronic acid glycidyl methacrylate (HAGM) Photoinitiator lithium phenyl-2,4,6-trimethylbenzoylphosphinate (LAP) and PEGDA between | Substrate micropatterning: 3D methacrylated gelatin (GelMA) scaffolds patterned via Microscale Continuous Optical Printing (μCOP) | Liu, Justin et al. “Direct 3D bioprinting of cardiac micro-tissues mimicking native myocardium.” Biomaterials 256 (2020): 120204. [86] | ||
| 3D | hiPSC-CMs, hiPSC-cECs, and hiPSC-cFswild-type (WT) hiPSC line LUMC0020iCTRL-06, long-QT syndrome type 1 (LQT1) hiPSC line LUMC0021iKCNQ-30 | Non-adhesive v-shaped plates; Matrigel-coated multielectrode array (MEA) | Non-structured microtissues | Giacomelli, Elisa et al. “Cardiac microtissues from human pluripotent stem cells recapitulate the phenotype of long-QT syndrome.” Biochemical and Biophysical Research Communications. 572 (2021): 118–124. [60] | ||
| Investigation of cell maturation in vitro | 3D | hiPSC-CMs and hiPSC-cardiacECs, hiPSC-CFs (CMFs), and Dermal Fibroblasts | V-bottomed 96-well microplates (Greiner bio-one) | Non-structured spheroids, 5000 cells each (85% cardiomyocytes and 15%endothelial cells) | Giacomelli, Elisa et al. “Human-iPSC-derived cardiac stromal cells enhance maturation in 3D cardiac microtissues and reveal non-cardiomyocyte contributions to heart disease.” Cell stem cell 26.6 (2020): 862–879. [87] | |
| 3D | hESCs (WiCell line) | 6-well ultra-low attachment plates | Non-structured spheroids with fibrosis modeling; coculturing | Lee, Mi-Ok et al. “Modelling cardiac fibrosis using three-dimensional cardiac microtissues derived from human embryonic stem cells.” Journal of biological engineering 13.1 (2019): 1–17. [88] | ||
| 3D | iPSC: human iPSC cell line DF 19-9-7 T (iPS, karyotype: 46, XY) | 12-well Matrigel-coated plates | hiPSC-derived 3D organotypic cardiac microtissue (hOCMT); long-term culturing | Ergir, Ece et al. “Generation and maturation of human iPSC-derived 3D organotypic cardiac microtissues in long-term culture.” Scientific Reports 12.1 (2022): 17409. [89] | ||
| Investigation of mechanotransduction-related events in vitro | 3D | NIH 3T3 fibroblasts (American Type Culture Collection, ATCC) | PDMS with Pluronic F127 and collagen; fibronectin | Well-structured sheets; Substrate micropatterning | Legant, Wesley R., Christopher S. Chen, and Viola Vogel. “Force-induced fibronectin assembly and matrix remodeling in a 3D microtissue model of tissue morphogenesis.” Integrative Biology 4.10 (2012): 1164–1174. [90] | |
| 3D | hCM: AC16 (a human cardiomyocyte cell line; Merck Millipore, Burlington, MA, USA) and hCF (a human normal cardiac fibroblast-ventricular cell line; Lonza, Basel, Switzerland) cells | Cell culture dish | Structured sheets by magnetic torque stimulation (MTS) system: hCM was seeded with magnet nanoparticles conjugated with WGA | Song, M.; Kim, J.; Shin, H.; Kim, Y.; Jang, H.; Park, Y.; Kim, S.-J. Development of Magnetic Torque Stimulation (MTS) Utilizing Rotating Uniform Magnetic Field for Mechanical Activation of Cardiac Cells. Nanomaterials 2020, 10, 1684. https://doi.org/10.3390/nano10091684 [91] | ||
| 3D | Human fetal cardiac fibroblasts (hfCF) | Spin-coated with SU8-2050 silicon wafers, PDMS | Step-profile steel fibrosis-on-chip platform | Gartner, Tom CL Bracco et al. “Cyclic strain has antifibrotic effects on the human cardiac fibroblast transcriptome in a human cardiac fibrosis-on-a-chip platform.” Journal of the Mechanical Behavior of Biomedical Materials 144 (2023): 105980. [92] | ||
| Investigation of biomarkers | 2D, 3D | hiPS-CMs (iCell™ Cardiomyocytes), primary human cardiac microvascular endothelial cells (hCMECs), and primary human cardiac fibroblasts (hCFs) | Ultra-low attachment spheroid microplates (Corning, 3830); CellCarrier-384 Ultra microplates | Non-structured microtissues; coculturing; and modeling fibrosis | Archer, Caroline R. et al. “Characterization and validation of a human 3D cardiac microtissue for the assessment of changes in cardiac pathology.” Scientific reports 8.1 (2018): 10160. [93] | |
| 3D | iPS cells: human cardiac microtissues (3D InSightTM Human Cardiac Microtissues from InSphero) and human biopsies | Non-adhesive plates | Non-structured microtissues; coculturing; and modeling fibrosis | Nguyen, Nhan et al. “Translational proteomics analysis of anthracycline-induced cardiotoxicity from cardiac microtissues to human heart biopsies.” Frontiers in Genetics 12 (2021): 695625. [94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aitova, A.; Berezhnoy, A.; Tsvelaya, V.; Gusev, O.; Lyundup, A.; Efimov, A.E.; Agapov, I.; Agladze, K. Biomimetic Cardiac Tissue Models for In Vitro Arrhythmia Studies. Biomimetics 2023, 8, 487. https://doi.org/10.3390/biomimetics8060487
Aitova A, Berezhnoy A, Tsvelaya V, Gusev O, Lyundup A, Efimov AE, Agapov I, Agladze K. Biomimetic Cardiac Tissue Models for In Vitro Arrhythmia Studies. Biomimetics. 2023; 8(6):487. https://doi.org/10.3390/biomimetics8060487
Chicago/Turabian StyleAitova, Aleria, Andrey Berezhnoy, Valeriya Tsvelaya, Oleg Gusev, Alexey Lyundup, Anton E. Efimov, Igor Agapov, and Konstantin Agladze. 2023. "Biomimetic Cardiac Tissue Models for In Vitro Arrhythmia Studies" Biomimetics 8, no. 6: 487. https://doi.org/10.3390/biomimetics8060487
APA StyleAitova, A., Berezhnoy, A., Tsvelaya, V., Gusev, O., Lyundup, A., Efimov, A. E., Agapov, I., & Agladze, K. (2023). Biomimetic Cardiac Tissue Models for In Vitro Arrhythmia Studies. Biomimetics, 8(6), 487. https://doi.org/10.3390/biomimetics8060487