
Dual Magnetic Particle Imaging and Akaluc Bioluminescence Imaging for Tracking Cancer Cell Metastasis

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line Origins and Culture
2.2. Lentiviral Transduction
2.3. Cell Characterization
2.4. Cell Labeling
2.5. Cell Viability
2.6. In Vitro BLI
2.7. Characterization of Synomag-D
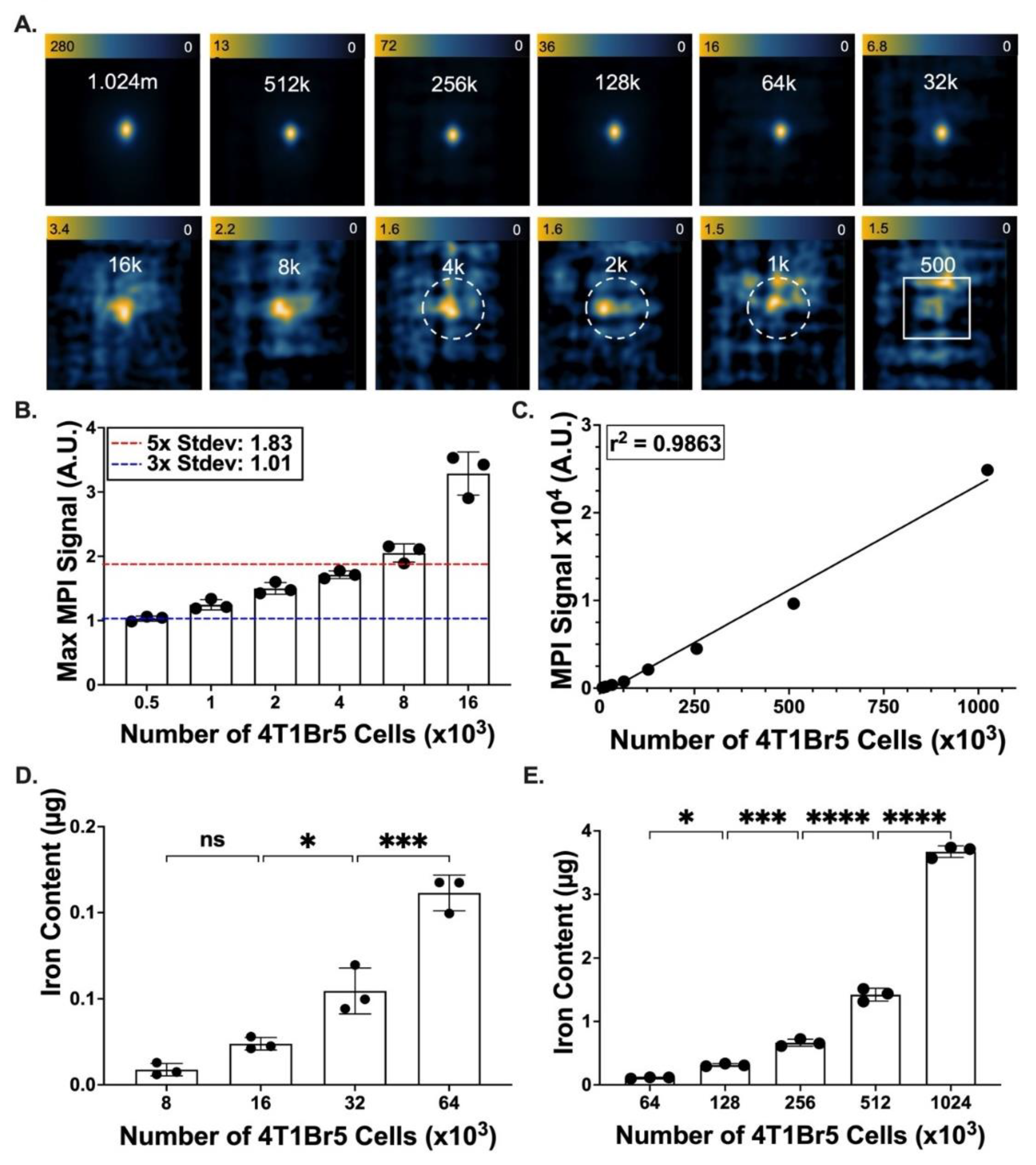
2.8. Determination of Detection Limits for Synomag-D Labelled 4T1Br5 Cells
2.9. Animal Model
2.10. In Vivo BLI and MPI
2.11. Image Analysis
2.12. Ex Vivo Analysis
2.13. Statistics
3. Results
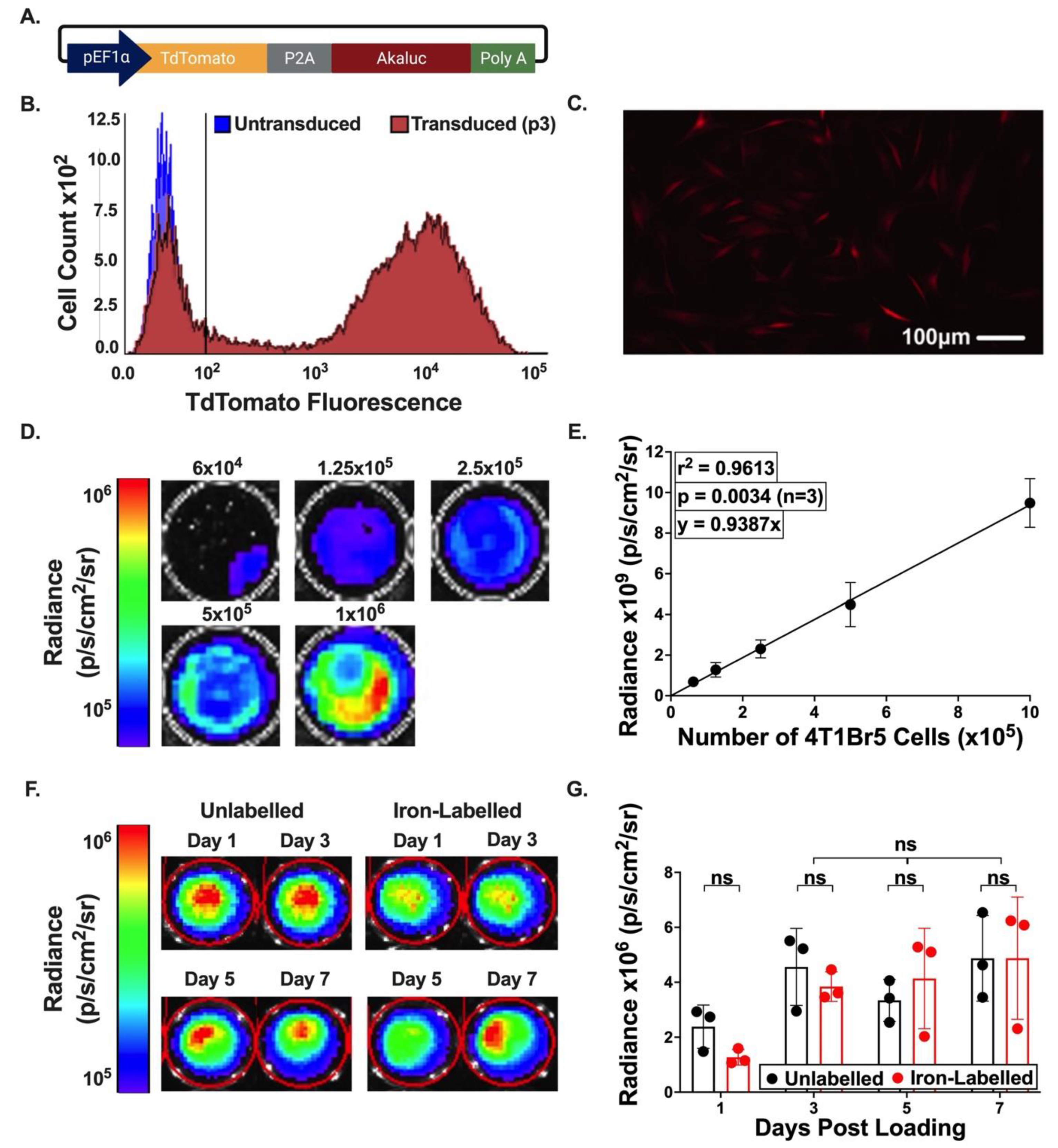
3.1. In Vitro Characterization of tdT-Akaluc Expressing Cells
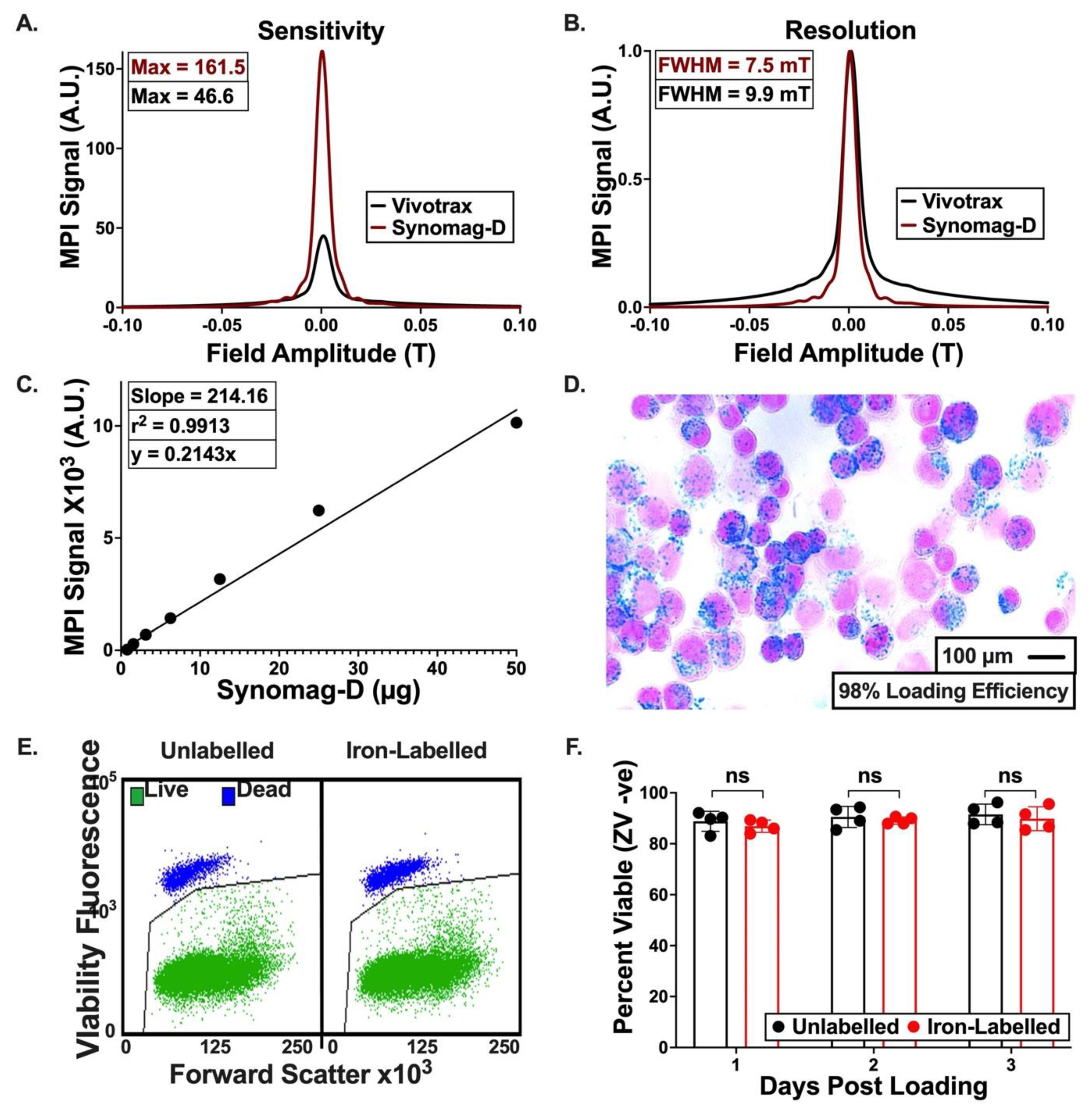
3.2. Characterization of Synomag-D
3.3. Iron Labeling of Cells
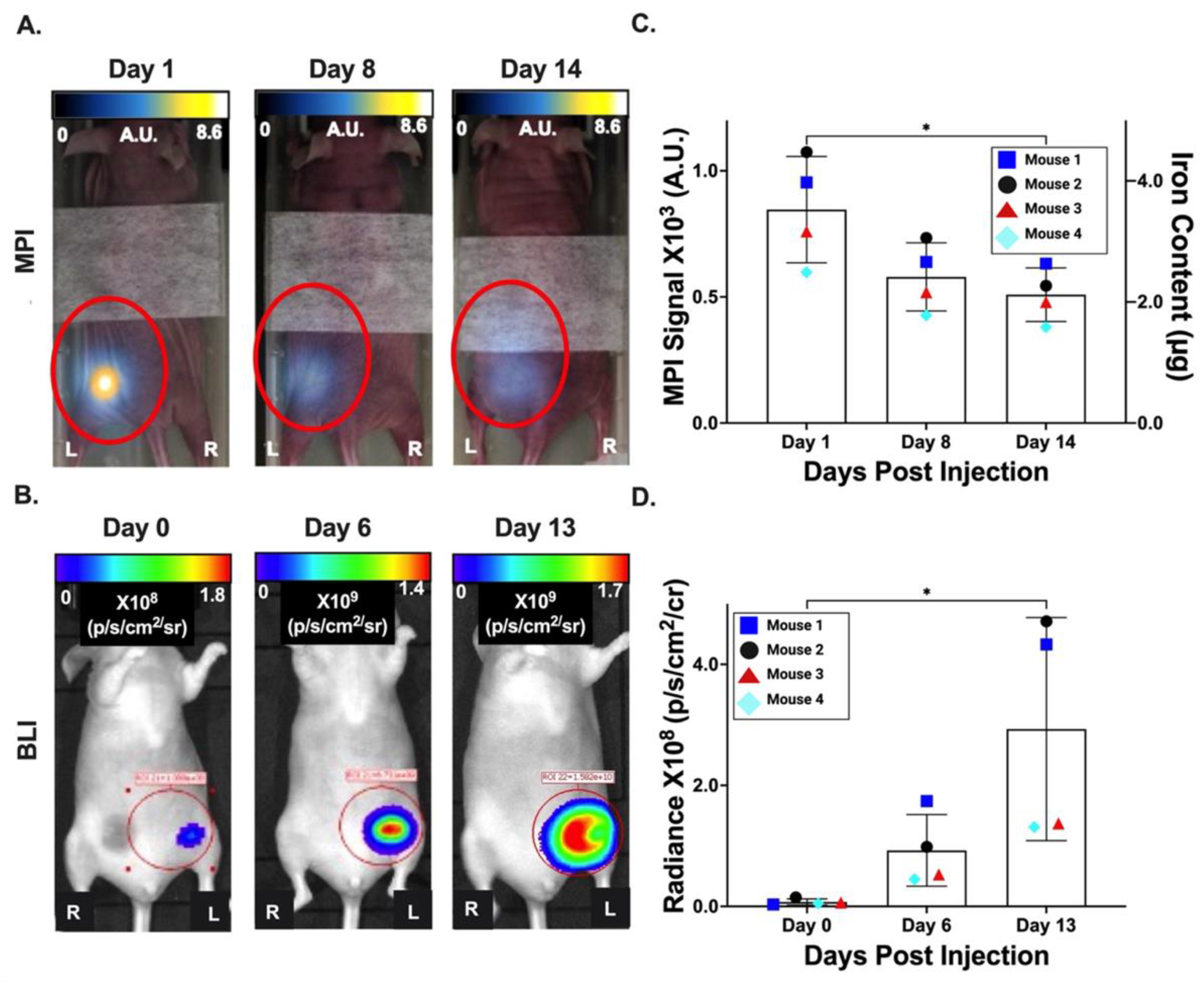
3.4. In Vivo Imaging of Primary Tumors
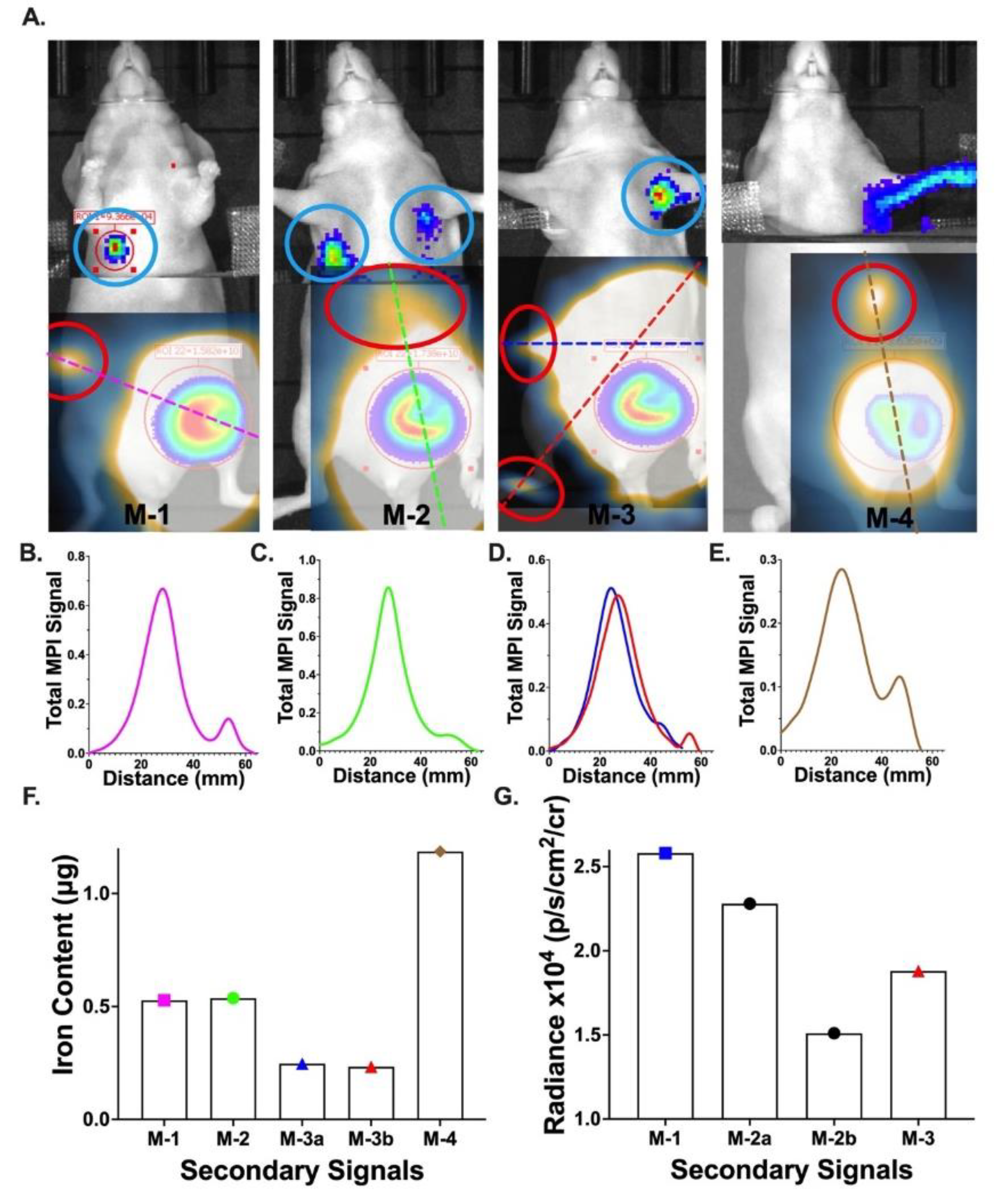
3.5. In Vivo Imaging of Spontaneous Metastases
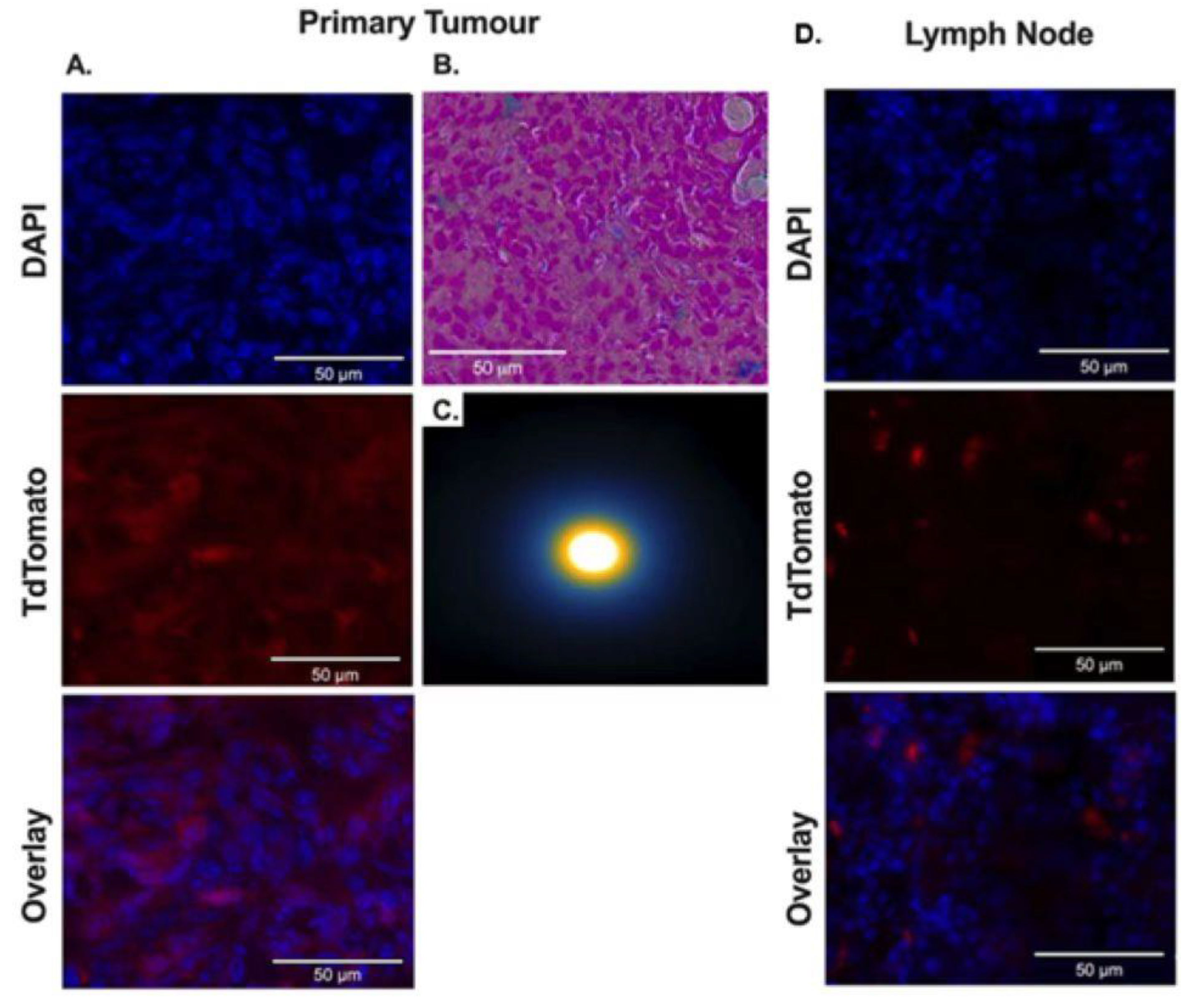
3.6. Ex Vivo Analysis of tdTomato Expression and Synomag D labeling in Tumors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med. 2019, 8, 5574–5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.T.; Sun, H.F.; Zhao, Y.; Fu, W.Y.; Yang, L.P.; Gao, S.P.; Li, L.D.; Jiang, H.L.; Jin, W. Comparison of patterns and prognosis among distant metastatic breast cancer patients by age groups: A SEER population-based analysis. Sci. Rep. 2017, 7, 9254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagan, R.; Meyer, P. Rethinking cancer: Current challenges and opportunities in cancer research. Dis. Model. Mech. 2017, 10, 349–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, Z.W.; Chandrasekharan, P.; Fellows, B.D.; Arrizabalaga, I.R.; Yu, E.; Olivo, M.; Conolly, S.M. Magnetic Particle Imaging: An emerging modality with prospects in diagnosis, targeting and therapy of cancer. Cancers 2021, 13, 5285. [Google Scholar] [CrossRef]
- Yang, X.; Shao, G.; Zhang, Y.; Wang, W.; Qi, Y.; Han, S.; Li, H. Applications of Magnetic Particle Imaging in biomedicine: Advancements and Prospects. Front. Physiol. 2022, 13, 898426. [Google Scholar] [CrossRef]
- Sehl, O.C.; Gevaert, J.J.; Melo, K.P.; Knier, N.N.; Foster, P.J. A Perspective on Cell Tracking with Magnetic Particle Imaging. Tomography 2020, 6, 315–324. [Google Scholar] [CrossRef]
- Wu, L.C.; Zhang, Y.; Steinberg, G.; Qu, H.; Huang, S.; Cheng, M.; Bliss, T.; Du, F.; Rao, J.; Song, G.; et al. A Review of Magnetic Particle Imaging and Perspectives on Neuroimaging. Am. J. Neuroradiol. 2019, 40, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Saritas, E.U.; Goodwill, P.W.; Croft, L.R.; Konkle, J.J.; Lu, K.; Zheng, B.; Connolly, S. Magnetic particle imaging (MPI) for NMR and MRI researchers. J. Magn. Reson. 2013, 229, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Yu, E.; Orendorff, R.; Lu, K.; Konkle, J.J.; Tay, Z.W.; Hensley, D.; Zhou, X.Y.; Chandrasekharan, P.; Saritas, E.U.; et al. Seeing SPIOs Directly In Vivo with Magnetic Particle Imaging. Mol. Imaging Biol. 2017, 19, 385–390. [Google Scholar] [CrossRef]
- Yu, E.Y.; Bishop, M.; Zheng, B.; Ferguson, R.M.; Khandhar, A.P.; Kemp, S.J.; Krishnan, K.M.; Goodwill, P.W.; Conolly, S.M. Magnetic Particle Imaging: A Novel in Vivo Imaging Platform for Cancer Detection. Nano Lett. 2017, 17, 1648–1654. [Google Scholar] [CrossRef]
- Talebloo, N.; Gudi, M.; Robertson, N.; Wang, P. Magnetic Particle Imaging: Current Applications in Biomedical Research. J. Magn. Reson. Imaging 2019, 51, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Vazin, T.; Goodwill, P.W.; Conway, A.; Verma, A.; Saritas, E.U.; Schaffer, D.; Conolly, S.M. Magnetic Particle Imaging tracks the long-term fate of in vivo neural cell implants with high image contrast. Sci. Rep. 2015, 5, 14055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grüttner, A.; Kowalski, F.; Fidler, M.; Steinke, F.; Westphal; Teller, H. Nanoflower particles: A new tracer for mpi, physical characterization and initial in vitro toxicity studies. Int. Workshop Magn. Part. Imaging 2018, 8, 17–18. [Google Scholar]
- Vogel, P.; Kampf, T.; Rückert, M.A.; Grüttner, C.; Kowalski, A.; Teller, H.; Behr, V. Synomag®: The new high-performance tracer for magnetic particle imaging. Int. J. Magn. Part Imaging 2021, 7, 2103003. [Google Scholar]
- Paysen, H.; Loewa, N.; Stach, A.; Wells, J.; Korsch, O.; Twamley, S.; Makowski, M.R.; Schaeffter, T.; Ludwig, A.; Wiekhorst, F. Cellular uptake of magnetic nanoparticles imaged and quantified by magnetic particle imaging. Sci. Rep. 2020, 10, 1922. [Google Scholar] [CrossRef] [Green Version]
- Cromer Berman, S.M.; Walczak, P.; Bulte, J.W.M. Tracking stem cells using magnetic nanoparticles. Wiley Interdiscip. Rev. NanoMed. Nanobiotechnol. 2011, 3, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Amsalem, Y.; Mardor, Y.; Feinberg, M.S.; Landa, N.; Miller, L.; Daniels, D.; Ocherashvilli, A.; Holbova, R.; Barbash, I.; Leor, J. Iron-oxide labeling and outcome of transplanted mesenchymal stem cells in the infarcted myocardium. Circulation 2007, 116, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Alsawaftah, N.; Farooq, A.; Dhou, S.; Majdalawieh, A.F. Bioluminescence Imaging Applications in Cancer: A Comprehensive Review. IEEE Rev. BioMed. Eng. 2021, 1, 307–326. [Google Scholar] [CrossRef]
- Bu, L.; Ma, X.; Tu, Y.; Shen, B.; Cheng, Z. Optical image-guided cancer therapy. Curr. Pharm. Biotechnol. 2013, 14, 723–732. [Google Scholar] [CrossRef]
- Söling, A.; Rainov, N.G. Bioluminescence imaging in vivo—Application to cancer research. Expert. Opin. Biol. Ther. 2003, 3, 1163–1172. [Google Scholar] [CrossRef]
- Minegishi, M.; Kuchimaru, T.; Nakagawa, K.; Isozaki, T.; Fujimori, S.; Kadonosono, T.; Kizaka-Kondoh, S. Multiplexed bioluminescence imaging of cancer cell response to hypoxia and inflammation in the caudal-artery injection model of bone metastasis during zoledronic acid treatment. J. Cancer Metastasis Treat. 2021, 7, 5. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, Y.; Zhang, F.; McGregor, H.; Yang, X. Radiofrequency hyperthermia enhances locally delivered oncolytic immuno-virotherapy for pancreatic adenocarcinoma. Cardiovasc. Interv. Radiol. 2022, 45, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Parkins, K.M.; Hamilton, A.M.; Dubois, V.P.; Wong, S.M.; Foster, P.J.; Ronald, J.A. Cellular MRI reveals altered brain arrest of genetically engineered metastatic breast cancer cells. Contrast Media Mol. Imaging 2019, 2019, 6501231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhang, H.C.; Gabrielson, K.L.; Laterra, J.; Fisher, P.B.; Pomper, M.G. Tumor-specific imaging through progression elevated gene-3 promoter-driven gene expression. Nat. Med. 2011, 17, 123–129. [Google Scholar] [CrossRef]
- Stollfuss, J.; Landvogt, N.; Abenstein, M.; Ziegler, S.; Scwaiger, M.; Senekowitsch-Schmidtke, R.; Wieder, H. Non-invasive imaging of implanted peritoneal carcinomatosis in mice using PET and bioluminescence imaging. EJNMMI Res. 2015, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Kircher, M.F.; Gambhir, S.S.; Grimm, J. Noninvasive cell-tracking methods. Nat. Rev. Clin. Oncol. 2011, 8, 677–688. [Google Scholar] [CrossRef]
- James, M.L.; Gambhir, S.S. A molecular imaging primer: Modalities, imaging agents, and applications. Physiol. Rev. 2012, 92, 897–965. [Google Scholar] [CrossRef] [Green Version]
- Modo, M. Noninvasive imaging of transplanted cells. Curr. Opin. Organ. Transplant 2008, 13, 654–658. [Google Scholar] [CrossRef]
- Galli, F.; Varani, M.; Lauri, C.; Silveri, G.G.; Onofrio, L.; Signore, A. Immune cell labelling and tracking: Implications for adoptive cell transfer therapies. EJNMMI Radiopharm. Chem. 2021, 6, 7. [Google Scholar] [CrossRef]
- Liu, S.; Su, Y.; Lin, M.Z.; Ronald, J.A. Brightening up Biology: Advances in Luciferase Systems for in Vivo Imaging. ACS Chem. Biol. 2021, 16, 2707–2718. [Google Scholar] [CrossRef]
- Iwano, S.; Obata, R.; Miura, C.; Kiyama, M.; Hama, K.; Nakamura, M.; Amano, Y.; Kojima, S.; Hirano, T.; Maki, S.; et al. Development of simple firefly luciferin analogs emitting blue, green, red, and near-infrared biological window light. Tetrahedron 2013, 69, 3847–3856. [Google Scholar] [CrossRef]
- Kuchimaru, T.; Iwano, S.; Kiyama, M.; Mitsumata, S.; Kadonosono, T.; Niwa, H.; Kizaka-Kondoh, S. A luciferin analogue generating near-infrared bioluminescence achieves highly sensitive deep-tissue imaging. Nat. Commun. 2016, 7, 11856. [Google Scholar] [CrossRef] [PubMed]
- Iwano, S.; Sugiyama, M.; Hama, H.; Watakabe, A.; Hasegawa, N.; Kuchimaru, T.; Tanaka, K.; Takahashi, M.; Ishida, Y.; Hata, J.; et al. Single-cell bioluminescence imaging of deep tissue in freely moving animals. Science 2018, 359, 935–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thu, M.S.; Bryant, L.H.; Coppola, T.; Jordan, E.K.; Budde, M.D.; Lewis, B.K.; Chaudhry, A.; Ren, J.; Varma, N.R.S.; Arbab, A.S.; et al. Self-assembling nanocomplexes by combining ferumoxytol, heparin and protamine for cell tracking by magnetic resonance imaging. Nat. Med. 2012, 18, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockman, P.R.; Mittapalli, R.K.; Taskar, K.S.; Rudraraju, V.; Gril, B.; Bohn, K.A.; Adkins, C.E.; Roberts, A.; Thorsheim, H.R.; Gaasch, J.A.; et al. Heterogeneous blood-tumor barrier permeability determines drug efficacy in experimental brain metastases of breast cancer. Clin. Cancer Res. 2010, 16, 5664–5678. [Google Scholar] [CrossRef] [Green Version]
- McFadden, C.; Mallett, C.L.; Foster, P.J. Labeling of multiple cell lines using a new iron oxide agent for cell tracking by MRI. Contrast Media Mol. Imaging 2011, 6, 514–522. [Google Scholar] [CrossRef]
- Sehl, O.C.; Tiret, B.; Berih, M.A.; Makela, A.V.; Goodwill, P.W.; Foster, P.J. MPI region of interest (ROI) analysis and quantification of iron in different volumes. IJMPI 2022, 8, 5–7. [Google Scholar]
- Abcam. Ab270789 Live and Dead Ccll Assay Kit (Calcein AM, 7-AAD). 2021. Available online: https://www.abcam.com/ps/products/270/ab270789/documents/Live-and-Dead-Cell-Assay-protocol-book-v2-ab270789%20(website).pdf (accessed on 13 December 2021).
- Heyn, C.; Ronald, J.A.; Mackenzie, L.T.; MacDonald, I.C.; Chambers, A.F.; Rutt, B.K.; Foster, P.J. In vivo magnetic resonance imaging of single cells in mouse brain with optical validation. Magn. Reson. Med. 2006, 55, 23–29. [Google Scholar] [CrossRef]
- Heyn, C.; Ronald, J.A.; Ramadan, S.S.; Snir, J.A.; Barry, A.M.; Mackenzie, L.T.; Mikulis, D.J.; Palmieri, D.; Bronder, J.L.; Steeg, P.S.; et al. In vivo MRI of cancer cell fate at the single-cell level in a mouse model of breast cancer metastasis to the brain. Magn. Reson. Med. 2006, 56, 1001–1010. [Google Scholar] [CrossRef]
- Foster, P.J.; Dunn, E.A.; Karl, K.E.; Snir, J.A.; Nycz, C.N.; Harvey, A.J.; Pettis, R.J. Cellular magnetic resonance imaging: In vivo imaging of melanoma cells in lymph nodes of mice. Neoplasia 2008, 10, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, E.M.; Sharer, K.; Skrtic, S.; Koretsky, A.P. In vivo detection of single cells by MRI. Magn. Reson. Med. 2006, 55, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Dahnke, H.; Jordan, E.K.; Schaeffter, T.; Frank, J.A. In vivo MRI using positive-contrast techniques in detection of cells labeled with superparamagnetic iron oxide nanoparticles. NMR BioMed. 2008, 21, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.O.; Jo, H.; Yu, J.H.; Gambhir, S.S.; Pratx, G. Development and MPI tracking of novel hypoxia-targeted theranostic exosomes. Biomaterials 2018, 177, 139–148. [Google Scholar] [CrossRef]
- Song, G.; Chen, M.; Zhang, Y.; Cui, L.; Qu, H.; Zheng, X.; Wintermark, M.; Liu, Z.; Rao, J. Janus Iron Oxides @ Semiconducting Polymer Nanoparticle Tracer for Cell Tracking by Magnetic Particle Imaging. Nano Lett. 2018, 18, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Parkins, K.M.; Melo, K.P.; Ronald, J.A.; Foster, P.J. Visualizing tumour self-homing with magnetic particle imaging. bioRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Melo, K.P.; Makela, A.V.; Hamilton, A.M.; Foster, P.J. Development of Magnetic Particle Imaging (MPI) for Cell Tracking and Detection. bioRxiv 2020. [Google Scholar]
- Knier, N.N.; Foster, P.J. Tracking cancer cells in the mouse brain with magnetic resonance imaging (MRI) and magnetic particle imaging (MPI). ISMRM 2021, 2021. [Google Scholar]
- Makela, A.V.; Schott, M.A.; Sehl, O.C.; Gevaert, J.J.; Foster, P.J.; Contag, C.H. Tracking the fates of iron-labeled tumor cells in vivo using Magnetic Particle Imaging. Nanoscale Adv. 2022, 4, 3617–3623. [Google Scholar] [CrossRef]
- Branchini, B.R.; Southworth, T.L.; DeAngelis, J.P.; Roda, A.; Michelini, E. Luciferase from the Italian firefly Luciola italica: Molecular cloning and expression. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 159–167. [Google Scholar] [CrossRef]
- Liu, S.; Nyström, N.N.; Kelly, J.J.; Hamilton, A.M.; Fu, Y.; Ronald, J.A. Molecular Imaging Reveals a High Degree of Cross-Seeding of Spontaneous Metastases in a Novel Mouse Model of Synchronous Bilateral Breast Cancer. Mol. Imaging Biol. 2022, 24, 104–114. [Google Scholar] [CrossRef]
- Ichise, H.; Tsukamoto, S.; Hirashima, T.; Konishi, Y.; Oki, C.; Tsukiji, S.; Iwano, S.; Miyawaki, A.; Sumiyama, K.; Terai, K.; et al. Metastatic Single Tumor Cells Evade NK Cell-mediated Killing by Thrombin-mediated Loss of the Activating Ligand CD155/PVR/Necl-5. bioRxiv 2021. [Google Scholar]
- Sehl, O.C.; Foster, P.J. The sensitivity of magnetic particle imaging and fluorine-19 magnetic resonance imaging for cell tracking. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Gevaert, J.J.; Fink, C.; Dikeakos, J.D.; Dekaban, G.A.; Foster, P.J. Magnetic Particle Imaging Is a Sensitive In Vivo Imaging Modality for the Detection of Dendritic Cell Migration. Mol. Imaging Biol. 2022, 24, 886–897. [Google Scholar] [CrossRef]
- Amintas, S.; Bedel, A.; Moreau-Gaudry, F.; Boutin, J.; Buscail, L.; Merlio, J.; Vendrely, V.; Dabernat, S.; Buscail, E. Circulating Tumor Cell Clusters: United We Stand Divided We Fall. Int. J. Mol. Sci. 2020, 21, 2653. [Google Scholar] [CrossRef] [Green Version]
- Economopoulos, V.; Chen, Y.; McFadden, C.; Foster, P.J. MRI detection of nonproliferative tumor cells in lymph node metastases using iron oxide particles in a mouse model of breast cancer. Transl. Oncol. 2013, 6, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makela, A.V.; Murrell, D.H.; Parkins, K.M.; Kara, J.; Gaudet, J.M.; Foster, P.J. Cellular imaging with MRI. Top Magn. Reson. Imaging 2016, 25, 177–186. [Google Scholar] [CrossRef]
- Su, Y.; Walker, J.; Park, Y.; Smith, T.P.; Liu, L.X.; Hall, M.P.; Labanieh, L.; Hurst, R.; Wang, D.; Encell, L.P.; et al. Novel NanoLuc substrates enable bright two-population bioluminescence imaging in animals. Nat. Methods 2020, 17, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Rodriguez, A.; Hoang-Minh, L.B.; Chiu-Lam, A.; Sarna, N.; Marrero-Morales, L.; Mitchell, D.A.; Rinaldi-Ramos, C.M. Tracking adoptive t cell immunotherapy using magnetic particle imaging. Nanotheranostics 2021, 5, 431–444. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, R.J.; Sehl, O.C.; Gevaert, J.J.; Liu, S.; Kelly, J.J.; Foster, P.J.; Ronald, J.A. Dual Magnetic Particle Imaging and Akaluc Bioluminescence Imaging for Tracking Cancer Cell Metastasis. Tomography 2023, 9, 178-194. https://doi.org/10.3390/tomography9010016
Williams RJ, Sehl OC, Gevaert JJ, Liu S, Kelly JJ, Foster PJ, Ronald JA. Dual Magnetic Particle Imaging and Akaluc Bioluminescence Imaging for Tracking Cancer Cell Metastasis. Tomography. 2023; 9(1):178-194. https://doi.org/10.3390/tomography9010016
Chicago/Turabian StyleWilliams, Ryan J., Olivia C. Sehl, Julia J. Gevaert, Shirley Liu, John J. Kelly, Paula J. Foster, and John A. Ronald. 2023. "Dual Magnetic Particle Imaging and Akaluc Bioluminescence Imaging for Tracking Cancer Cell Metastasis" Tomography 9, no. 1: 178-194. https://doi.org/10.3390/tomography9010016
APA StyleWilliams, R. J., Sehl, O. C., Gevaert, J. J., Liu, S., Kelly, J. J., Foster, P. J., & Ronald, J. A. (2023). Dual Magnetic Particle Imaging and Akaluc Bioluminescence Imaging for Tracking Cancer Cell Metastasis. Tomography, 9(1), 178-194. https://doi.org/10.3390/tomography9010016