
A Robust Protocol to Isolate Outer Membrane Vesicles from Nontypeable Haemophilus influenzae


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Design
2.1. Materials
- Nontypeable Haemophilus influenzae strains ATCC53600, 86-028NP, and NTHi 5
- Chocolate Agar Plates (Teknova, Hollister, CA, USA, cat no. C4900)
- Brain Heart Infusion broth (BHI, Teknova, Hollister, CA, USA, cat no. B9993)
- Fildes Enrichment (ThermoFisher Scientific, Waltham, MA, USA, cat no. R45037)
- Nicotinamide Adenine Dinucleotide (NAD, Millipore Sigma, St. Louis, MO, USA, cat no. 10127990001)
- Sterile 250 mL Polycarbonate Culture Flask (ThermoFisher Scientific, Waltham, MA, USA, cat no. PBV250)
- Two hundred fifty milliliter Nalgene Centrifuge Bottle (ThermoFisher Scientific, Waltham, MA, USA, cat no. 3141-0250PK)
- Sterile 150 mL Stericup Vacuum Filtration System, 0.22 μm, PVDF (Millipore Sigma, St. Louis, MO, USA, cat no. S2GVU01RE)
- Centricon Centrifugal Filter 100 kDa MWCO (Millipore Sigma, St. Louis, MO, USA, cat no. UFC710008)
- Three milliliter Thickwall Polycarbonate Tube, Beckman Coulter (ThermoFisher Scientific, Waltham, MA, USA, cat no. 362305)
- 1X Gibco Dulbecco’s Phosphate Buffered Saline (DPBS, ThermoFisher Scientific, Waltham, MA, USA, cat no. 14190144)
- Sterile 1 mL BD Insulin Syringes, 27 5/8 G (ThermoFisher Scientific, Waltham, MA, USA, cat no. 329412)
- Sterile BD 21G × 2 mm needle (ThermoFisher Scientific, Waltham, MA, USA, cat no. 305129)
- Sterile 3 mL Henke-Ject Luer Lock Syringe (ThermoFisher Scientific, Waltham, MA, USA, cat no. 4020-X00V0)
- Durapore 0.45 μm PVDF Syringe Filter (Millipore Sigma, St. Louis, MO, USA, cat no. SLHV013SL)
- Pierce Micro BCA Assay Kit (ThermoFisher Scientific, Waltham, MA, USA, cat no. 23235)
- Corning 96-well Microtiter Plate (ThermoFisher Scientific, Waltham, MA, USA, cat no. 07-200-89)
- Novex 4–20% Tris-Glycine WedgeWellTM Precast Gels (ThermoFisher Scientific, Waltham, MA, USA, cat no. XP04202BOX)
- Novex 10X Tris-Glycine SDS Running Buffer (ThermoFisher Scientific, Waltham, MA, USA, cat no. LC2675)
- Novex 2X Tris-Glycine SDS Sample Buffer (ThermoFisher Scientific, Waltham, MA, USA, cat no. LC2676)
- NuPAGE 10X Sample Reducing Agent (ThermoFisher Scientific, Waltham, MA, USA, cat no. NP0009)
- PageRuler Plus Prestained Protein Ladder (ThermoFisher Scientific, Waltham, MA, USA, cat no. 26619)
- InstantBlue Coomassie Stain (ThermoFisher Scientific, Waltham, MA, USA, cat no. ISB1L)
- Formvar/Carbon Supported 300 mesh Copper Grid (Millipore Sigma, St. Louis, MO, USA, cat no. TEM-FCF300CU50)
- Phosphotungstic Acid Hydrate (Millipore Sigma, St. Louis, MO, USA, cat no. 79690)
- Uranyl Acetate Dihydrate (ThermoFisher Scientific, Waltham, MA, USA, cat no. 18-607-644)
- Bovine Serum Albumin (BSA, Millipore Sigma, St. Louis, MO, USA, cat no. 10711454001)
2.2. Equipment
- 37 °C, 5% CO2 incubator with shaker
- SpectraMax M3 Plate Reader
- Beckman Coulter Avanti J-20 XP centrifuge with a JLA-16.250 rotor
- Sorvall Legend RT Tabletop Centrifuge
- Beckman Coulter Optima TLX Ultracentrifuge with a TLA100.4 rotor
- XCell SureLock Gel Electrophoresis Tank
- Electrophoresis Power Supply
- Platform Shaker
- BioRad ChemiDoc MP Gel Imager
- FEI Tecnai Spirit Twin Transmission Electron Microscope
3. Procedure
3.1. Isolation of Outer Membrane Vesicles (OMVs)
- Streak out NTHi strains onto separate chocolate agar plates and incubate overnight in a 37 °C, 5% CO2 incubator.
- Pre-warm 150 mL of NTHi growth medium (BHI broth + 2% Fildes enrichment + 20 μg/mL NAD) in 250 mL vented shake flask to 37 °C.
- Inoculate pre-warmed media with 3–5 isolated colonies of NTHi from step 1 and incubate overnight in a 37 °C, 5% CO2 incubator, shaking at 250 rpm.
- To obtain a measure of culture density, prepare a 1:10 dilution of the overnight culture by mixing 100 μL of the overnight culture and 900 μL fresh growth medium. Measure the OD600 of the resulting suspension. Multiply the OD600 by 10 to account for the 1:10 dilution.
- Transfer the remainder of the overnight culture to a 250 mL centrifuge bottle and pellet the cells at 10,000× g for 10 min at 4 °C. Carefully remove the supernatant and discard the bacterial pellet.
- Filter the supernatant using a 0.22 μm PVDF Stericup vacuum filter flask and collect the filtrate.
- To concentrate the filtered supernatant, use 2 Centricon centrifugal filters loading 60 mL of supernatant into each filter. Discard the remaining ~30 mL of supernatant (only 120 mL of the supernatant will be used for OMV isolation). The total volume of the supernatant can be scaled up by increasing the initial overnight culture as needed based on the yield obtained from initial trials.
- Centrifuge the filter assemblies at 3500× g for 20 min at 4 °C in a Sorvall Legend RT tabletop centrifuge using a swinging bucket rotor. Other NTHi strains or corresponding isogenic mutants may require longer run times for supernatant concentration. In each case, extend run time in 10 min increments.
- When the retentate in the Centricon filter reaches a volume of ~2 mL, elute the concentrated culture supernatant using the Centricon collection cup. Spin down the assembly at 1000× g for 1 min at 4 °C. Note that at this point, the concentrated supernatant will be slightly more viscous and visibly darker due to the concentration of the Fildes enrichment.
- Transfer the concentrated supernatant containing the OMVs into 3 mL thick-walled polycarbonate tubes and add cold 1X DPBS until the total volume is ~3 mL.
- To check the sterility of the OMV prep, spread a 50 μL aliquot onto a fresh chocolate agar plate. Incubate the plate overnight at 37 °C, 5% CO2. If colonies grow on the plate overnight, discard the entire OMV suspension as it contains contaminating viable bacterial cells.
- Pellet the OMVs by centrifuging tubes at 135,000× g (~50,000 rpm) for 18 h (overnight) at 4 °C in the Optima TLX ultracentrifuge using a TLA100.4 rotor. If time permits, this step can be reduced to a minimum of 5 h without compromising the yield. If there are NTHi colonies in the agar plates from Step 11, stop here and discard the OMV pellet, otherwise, proceed to Step 13.
- Carefully remove as much of the supernatant as possible using a 1 mL pipette. Once the OMV pellet is visible, photographs can be taken against a white background if desired.
- Fully resuspend the OMV pellet in 1 mL of ice-cold 1X DPBS by pipetting up and down several times until no visible clumps can be observed. Add 2 mL of fresh ice-cold 1X DPBS. Vortex the tubes at max speed for 30 s. This step washes away the soluble proteins that may be loosely adsorbed to the OMV surface. More importantly, this also washes away most of the remaining Fildes enrichment associated with the OMVs.
- Pellet the OMVs by centrifuging at 135,000× g for 5 h at 4 °C using the Optima TLX ultracentrifuge using the TLA100.4 rotor.
- Carefully remove as much of the supernatant as possible leaving only a wet OMV pellet.
- Resuspend the OMV pellet in 1 mL of ice-cold 1X DPBS until no visible clumps can be observed. This volume can be increased to 2–3 mL depending on how much starter culture was used. This volume is called Vres and will be used later to calculate OMV yield.
- To further resuspend the OMVs and make sure that no macroscopic OMV aggregates remain, pass the OMV suspension through a 1 cc Insulin syringe three times and vortex the suspension at max speed for 30 s.
- Pass the OMV suspension three times through a 21G × 2 mm needle attached to a 3 cc luer-lock syringe making sure to collect the OMV suspension inside the syringe after the third pass. This step further ensures that any small OMV aggregates are broken up.
- Aspirate ~1 mL of air to have an air pocket between the needle and the OMV suspension inside the syringe.
- While the OMV suspension is inside the syringe, carefully remove the needle and replace it with a 0.45 μm PVDF syringe filter.
- Pass the OMV suspension through the syringe filter collecting the final suspension in a sterile 1.5 mL tube.
- Divide the OMV suspension into 250 μL aliquots (or desired volume) and freeze at −80 °C.
3.2. Quantification of Protein Content and Calculation of OMV Yield
- Thaw an aliquot of the OMV suspension on ice for ~30 min. (Any future use of the OMVs should be preceded by the thawing of the frozen suspension on ice.)
- When measuring the protein concentration of the OMVs, prepare a dilution series of the OMVs to account for the possibility of the OMV protein concentration exceeding the upper limits of the calibration curve. Typically, a 1:4 starting dilution is necessary.
- Measure the protein concentration of the OMV prep using the Pierce MicroBCA Assay Kit according to the manufacturer’s protocol and using the accompanying bovine serum albumin (BSA) as a standard.
- Perform the MicroBCA assay in a 96-well polystyrene microtiter plate and read the absorbance at 562 nm using a standard plate reader.
- Calculate the protein concentration of the OMV suspension using the calibration curve obtained in the MicroBCA Assay and, if necessary, account for any dilutions done previously.
- The OMV yield (per 120 mL culture supernatant) is calculated based on the obtained protein concentration and the Vres normalized against the OD600 of the starting culture using the following equations:
3.3. SDS-PAGE Analysis of OMV Protein Profile
- Prepare 56 μL of each OMV suspension at 430 μg/mL in 1X DPBS.
- Add 70 μL of 2X Tris-Glycine SDS Sample Buffer and 14 μL of 10X NuPAGE Sample Reducing Agent to the OMV samples.
- Split the mixture into two 70 μL aliquots labeling one “heated” and the other “unheated”. Boil the “heated” fraction at 100 °C for 10 min and allow to cool to room temperature.
- Load 50 μL of each “heated” and “unheated” sample into separate wells of a Novex 4–20% Tris-Glycine WedgeWell™ gel. Run the gel at a constant 225 V for 45 min using 1X Tris-Glycine SDS Running Buffer.
- Remove the gel carefully from the cassette and stain using InstantBlue Coomassie Stain overnight in a low-speed shaker at room temperature.
- Image the gel using a BioRad ChemiDoc MP Gel Imager or any similar gel visualization device.
3.4. Transmission Electron Microscopy (TEM) of OMVs
- Secure 300 mesh copper grids with forceps and clasp tightly. Rinse the grids with 0.01% BSA and wick off any excess solution using a filter paper.
- Place a 2 μL aliquot of each OMV suspension (several dilutions may be required to ensure optimum vesicle population in every field of view) onto each copper grid and allow to air dry for 10 min. Wick off any residual material in the copper grid using a filter paper.
- Stain the samples with 20 μL of 2% Phosphotungstic acid or 2% Uranyl Acetate and incubate grids for 60 s at room temperature. Remove any excess solution using a clean filter paper.
- Mount the copper grids onto an FEI Tecnai Spirit Twin Transmission Electron Microscope and examine samples at high magnification at 80 kV.
- Take multiple images throughout the grid. Measure the diameter of several OMVs using the pixel-to-micron conversion factor specific to the microscope used.
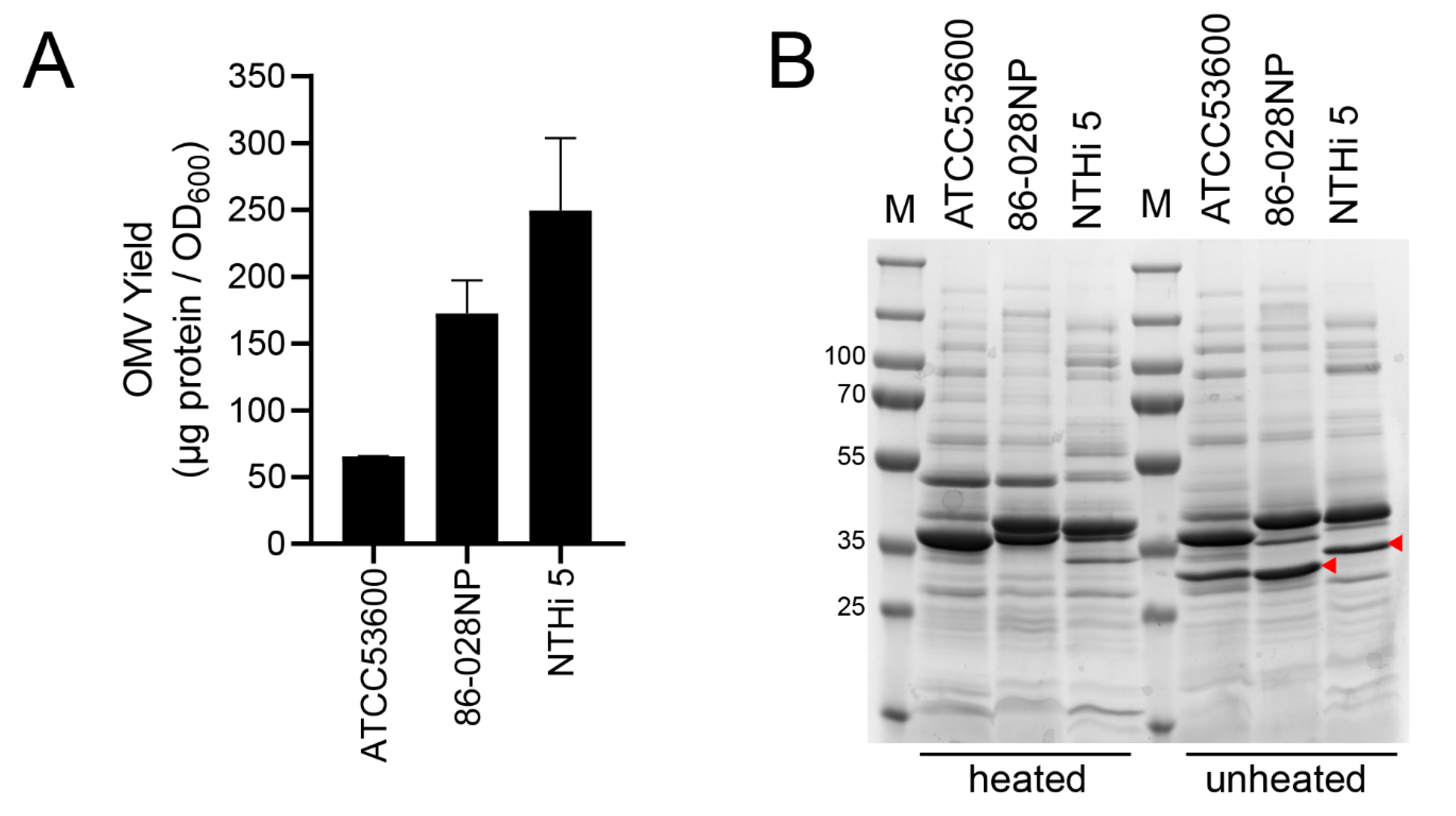
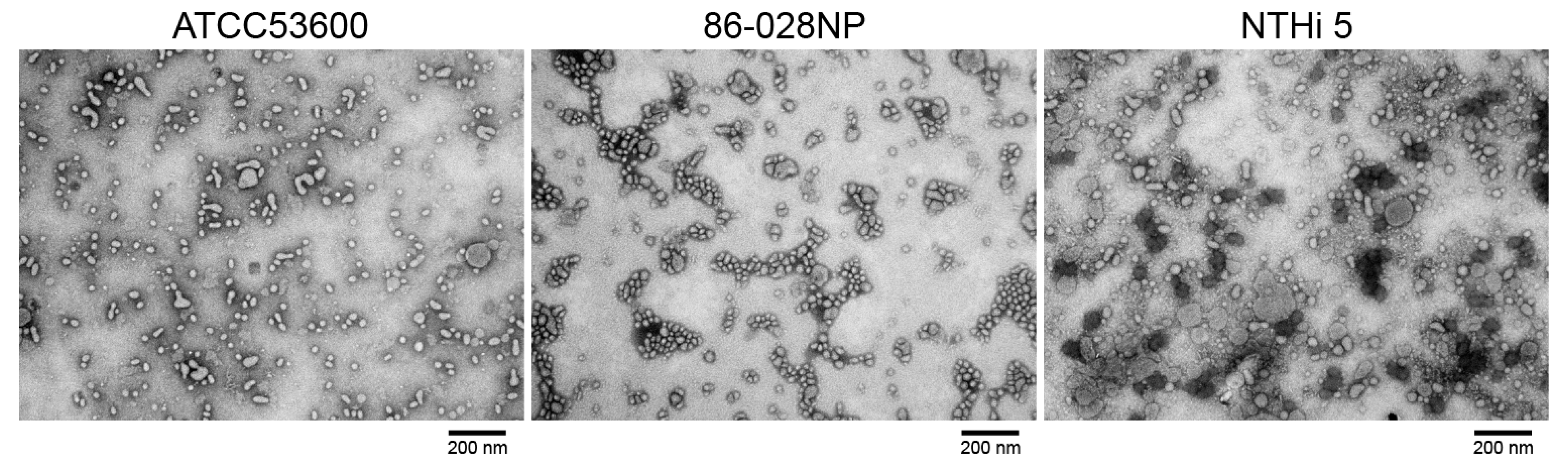
4. Expected Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [Green Version]
- Cecil, J.D.; Sirisaengtaksin, N.; O’Brien-Simpson, N.M.; Krachler, A.M. Outer Membrane Vesicle-Host Cell Interactions. Microbiol. Spectr. 2019, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberman, L.A. Outer membrane vesicles: A bacterial-derived vaccination system. Front. Microbiol. 2022, 13, 1029146. [Google Scholar] [CrossRef]
- Kashyap, D.; Panda, M.; Baral, B.; Varshney, N.; Sajitha, R.; Bhandari, V.; Parmar, H.S.; Prasad, A.; Jha, H.C. Outer Membrane Vesicles: An Emerging Vaccine Platform. Vaccines 2022, 10, 1578. [Google Scholar] [CrossRef]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The new multicomponent vaccine against meningococcal serogroup B, 4CMenB: Immunological, functional and structural characterization of the antigens. Vaccine 2012, 30 (Suppl. S2), B87–B97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.H.; Choi, H.I.; Hong, S.W.; Kim, K.S.; Gho, Y.S.; Jeon, S.G. Vaccination with Klebsiella pneumoniae-derived extracellular vesicles protects against bacteria-induced lethality via both humoral and cellular immunity. Exp. Mol. Med. 2015, 47, e183. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative immunogenicity and efficacy of equivalent outer membrane vesicle and glycoconjugate vaccines against nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef] [Green Version]
- Obiero, C.W.; Ndiaye, A.G.W.; Scire, A.S.; Kaunyangi, B.M.; Marchetti, E.; Gone, A.M.; Schutte, L.D.; Riccucci, D.; Auerbach, J.; Saul, A.; et al. A Phase 2a Randomized Study to Evaluate the Safety and Immunogenicity of the 1790GAHB Generalized Modules for Membrane Antigen Vaccine against Shigella sonnei Administered Intramuscularly to Adults from a Shigellosis-Endemic Country. Front. Immunol. 2017, 8, 1884. [Google Scholar] [CrossRef] [Green Version]
- Adriani, R.; Mousavi Gargari, S.L.; Nazarian, S.; Sarvary, S.; Noroozi, N. Immunogenicity of Vibrio cholerae outer membrane vesicles secreted at various environmental conditions. Vaccine 2018, 36, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Daleke-Schermerhorn, M.H.; Felix, T.; Soprova, Z.; Ten Hagen-Jongman, C.M.; Vikstrom, D.; Majlessi, L.; Beskers, J.; Follmann, F.; de Punder, K.; van der Wel, N.N.; et al. Decoration of outer membrane vesicles with multiple antigens by using an autotransporter approach. Appl. Environ. Microbiol. 2014, 80, 5854–5865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roier, S.; Blume, T.; Klug, L.; Wagner, G.E.; Elhenawy, W.; Zangger, K.; Prassl, R.; Reidl, J.; Daum, G.; Feldman, M.F.; et al. A basis for vaccine development: Comparative characterization of Haemophilus influenzae outer membrane vesicles. Int. J. Med. Microbiol. 2015, 305, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Roier, S.; Leitner, D.R.; Iwashkiw, J.; Schild-Prufert, K.; Feldman, M.F.; Krohne, G.; Reidl, J.; Schild, S. Intranasal immunization with nontypeable Haemophilus influenzae outer membrane vesicles induces cross-protective immunity in mice. PLoS ONE 2012, 7, e42664. [Google Scholar] [CrossRef]
- Winter, L.E.; Barenkamp, S.J. Immunogenicity of Nontypeable Haemophilus influenzae Outer Membrane Vesicles and Protective Ability in the Chinchilla Model of Otitis Media. Clin. Vaccine Immunol. 2017, 24, e00138-17. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, S.W.; Kuehn, M.J.; Mason, K.M. Elicitation of epithelial cell-derived immune effectors by outer membrane vesicles of nontypeable Haemophilus influenzae. Infect. Immun. 2011, 79, 4361–4369. [Google Scholar] [CrossRef] [Green Version]
- Behrouzi, A.; Mianroodi, R.A.; Afrough, P.; Ayadi, A.; Serajian, A. Evaluation of immunological responses against outer membrane vesicles (OMV) of nontypeable Haemophilus influenzae using MPLA-CpG adjuvant as a vaccine candidate. Iran J. Microbiol. 2020, 12, 417–423. [Google Scholar] [CrossRef]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Carcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [Green Version]
- Reimer, S.L.; Beniac, D.R.; Hiebert, S.L.; Booth, T.F.; Chong, P.M.; Westmacott, G.R.; Zhanel, G.G.; Bay, D.C. Comparative Analysis of Outer Membrane Vesicle Isolation Methods With an Escherichia coli tolA Mutant Reveals a Hypervesiculating Phenotype With Outer-Inner Membrane Vesicle Content. Front. Microbiol. 2021, 12, 628801. [Google Scholar] [CrossRef]
- De Chiara, M.; Hood, D.; Muzzi, A.; Pickard, D.J.; Perkins, T.; Pizza, M.; Dougan, G.; Rappuoli, R.; Moxon, E.R.; Soriani, M.; et al. Genome sequencing of disease and carriage isolates of nontypeable Haemophilus influenzae identifies discrete population structure. Proc. Natl. Acad. Sci. USA 2014, 111, 5439–5444. [Google Scholar] [CrossRef] [Green Version]
- Foxwell, A.R.; Kyd, J.M.; Cripps, A.W. Nontypeable Haemophilus influenzae: Pathogenesis and prevention. Microbiol. Mol. Biol. Rev. 1998, 62, 294–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakaletz, L.O.; Baker, B.D.; Jurcisek, J.A.; Harrison, A.; Novotny, L.A.; Bookwalter, J.E.; Mungur, R.; Munson, R.S., Jr. Demonstration of Type IV pilus expression and a twitching phenotype by Haemophilus influenzae. Infect. Immun. 2005, 73, 1635–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Libardo, M.D.J.; Durr, E.; Hernandez, L.D. A Robust Protocol to Isolate Outer Membrane Vesicles from Nontypeable Haemophilus influenzae. Methods Protoc. 2023, 6, 42. https://doi.org/10.3390/mps6020042
Libardo MDJ, Durr E, Hernandez LD. A Robust Protocol to Isolate Outer Membrane Vesicles from Nontypeable Haemophilus influenzae. Methods and Protocols. 2023; 6(2):42. https://doi.org/10.3390/mps6020042
Chicago/Turabian StyleLibardo, M. Daben J., Eberhard Durr, and Lorraine D. Hernandez. 2023. "A Robust Protocol to Isolate Outer Membrane Vesicles from Nontypeable Haemophilus influenzae" Methods and Protocols 6, no. 2: 42. https://doi.org/10.3390/mps6020042
APA StyleLibardo, M. D. J., Durr, E., & Hernandez, L. D. (2023). A Robust Protocol to Isolate Outer Membrane Vesicles from Nontypeable Haemophilus influenzae. Methods and Protocols, 6(2), 42. https://doi.org/10.3390/mps6020042