Evolutionary Philosophy of Science: A New Image of Science and Stance towards General Philosophy of Science


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Kuhn’s Evolutionary Philosophy of Science
3. Proposed Evolutionary Philosophy of Science
4. Historical Case Studies
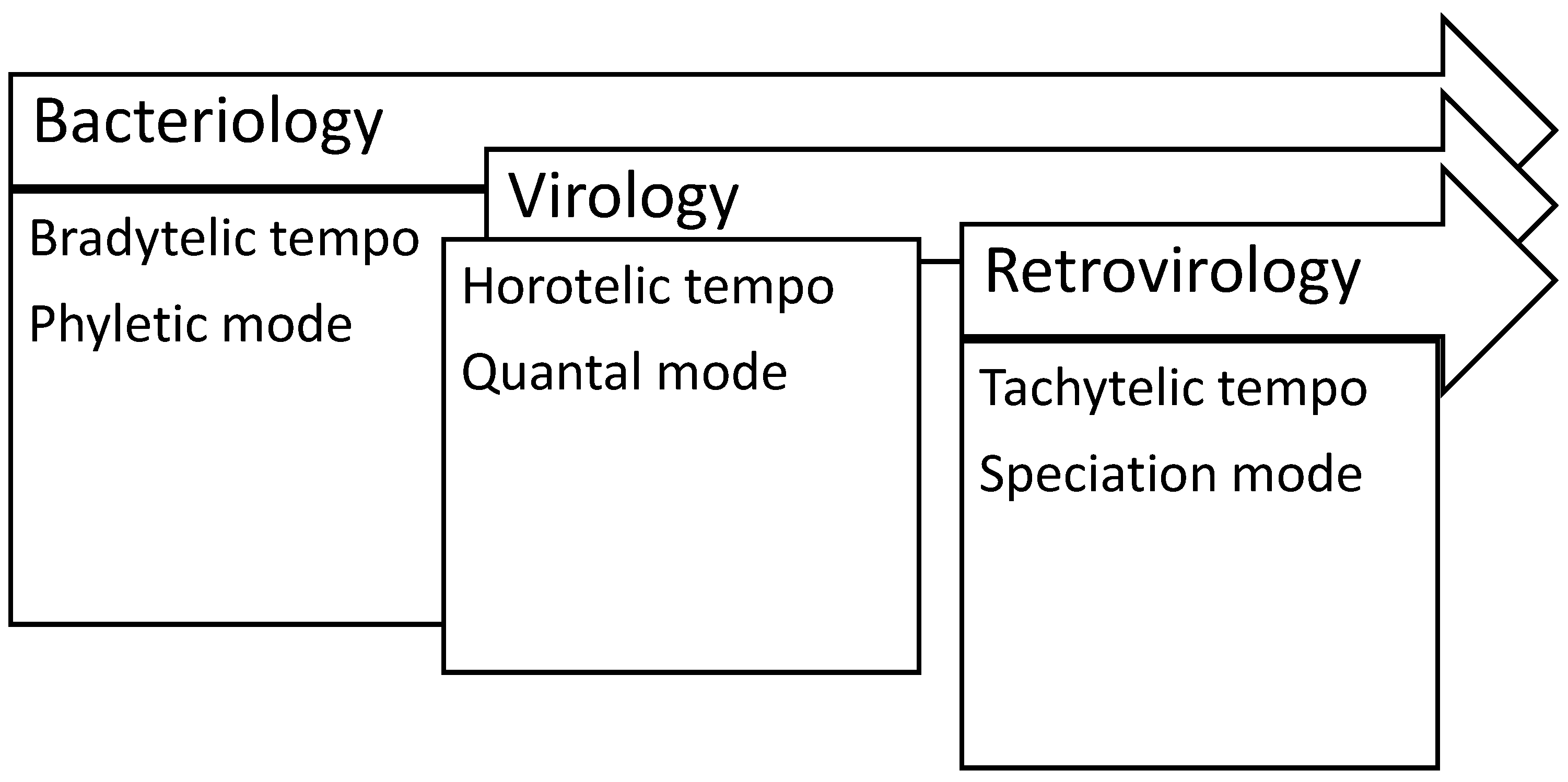
4.1. Microbiology Sciences
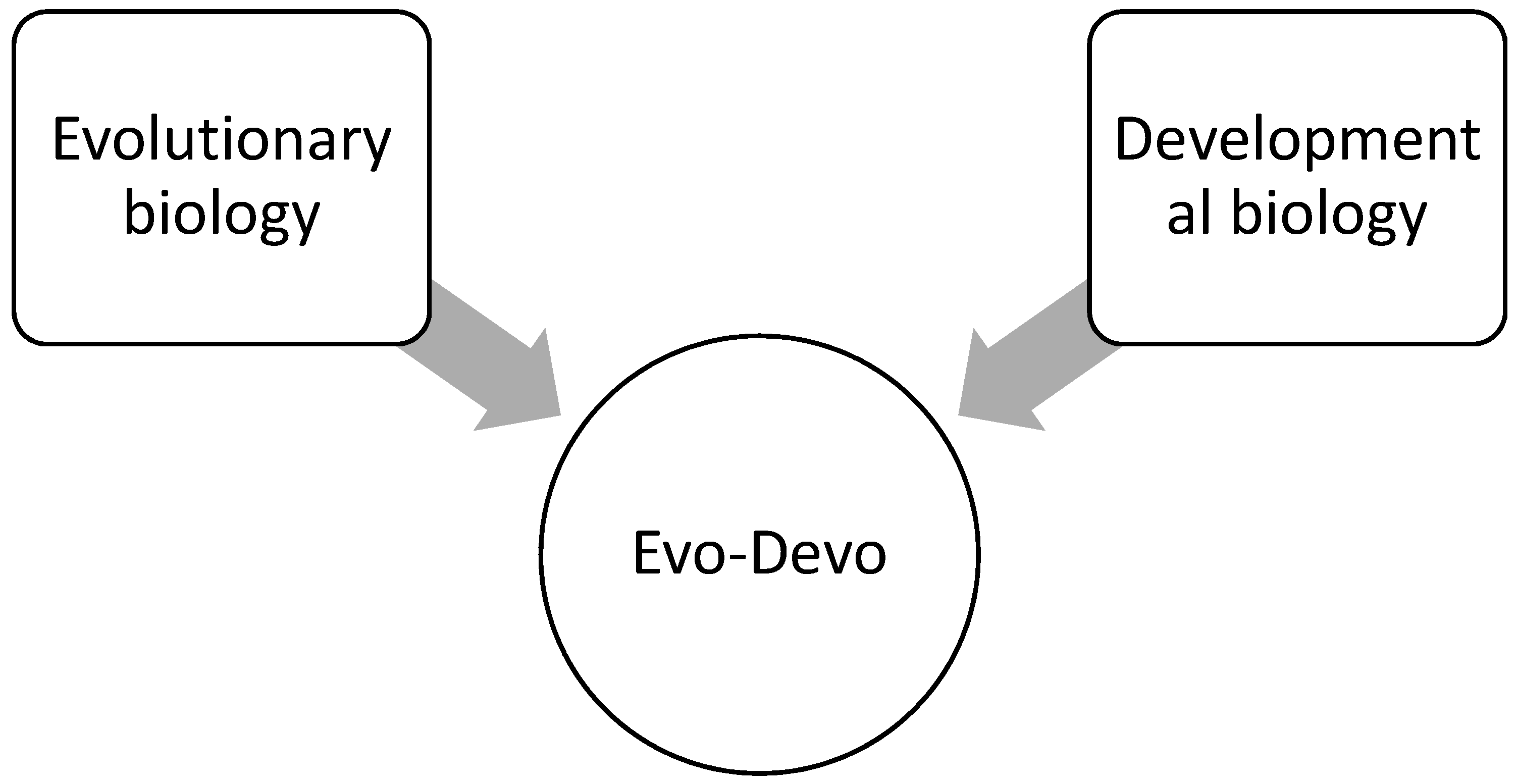
4.2. Evolutionary Developmental Biology
By all accounts ‘evo-devo’ has arrived. It is now solidly entrenched in the conceptual framework of modern biology and has all the markings of a new discipline, such as representation in professional societies, scientific journals devoted to the field, academic programs and job searches, panels at funding agencies, textbooks, etc.[49] (p. 1).
5. Conclusions
Conflicts of Interest
Abbreviations
| EPoS | evolutionary philosophy of science |
| GPoS | general philosophy of science |
| HPoS | historical philosophy of science |
| PPoS | pluralistic philosophy of science |
References and Notes
- Kellert, S.H.; Longino, H.E.; Waters, C.K. (Eds.) Scientific Pluralism; University of Minnesota Press: Minneapolis, MN, USA, 2006. [Google Scholar]
- Psillos, S. What is general philosophy of science? J. Gen. Philos. Sci. 2012, 43, 93–103. [Google Scholar] [CrossRef]
- Psillos, S. Having science in view: General philosophy of science and its significance. In The Oxford Handbook of Philosophy of Science; Humphrey, P., Ed.; Oxford University Press: New York, NY, USA, 2016; pp. 137–162. [Google Scholar]
- Kuhn, T.S. The Structure of Scientific Revolutions, 2nd ed.; University of Chicago Press: Chicago, IL, USA, 1970. [Google Scholar]
- Preston, J. Kuhn’s ‘The Structure of Scientific Revolutions’: A Reader’s Guide; Bloomsbury Press: London, UK, 2008. [Google Scholar]
- Marcum, J.A. Whither Kuhn’s historical philosophy of science? An evolutionary turn. In An Anthology of Philosophical Studies; Hanna, P., Ed.; Athens Institute for Education and Research: Athens, Greece, 2013; Volume 7, pp. 99–109. [Google Scholar]
- Kuhn, T.S. The Trouble with the Historical Philosophy of Science; Department of the History of Science, Harvard University: Cambridge, MA, USA, 1992. [Google Scholar]
- For Additional Discussion of Kuhn’s EPoS, Marcum, J.A. Thomas Kuhn’s Revolutions: An Historical and an Evolutionary Philosophy of Science? Bloomsbury Press: London, UK, 2015. [Google Scholar]
- Marcum, J.A. The evolving notion and role of Kuhn’s incommensurability thesis. In Kuhn’s ‘Structure of Scientific Revolutions’—50 Years On; Devlin, W.J., Bokulich, A., Eds.; Boston Studies in the Philosophy and History of Science; Springer: New York, NY, USA, 2015; Volume 311, pp. 115–134. [Google Scholar]
- Sankey, H. Taxonomic incommensurability. Int. Stud. Philos. Sci. 1998, 12, 7–16. [Google Scholar] [CrossRef]
- Damuth, J.D. Evolution: Tempo and mode. In Encyclopedia of Life Sciences; Wiley: New York, NY, USA, 2001; pp. 1–7. [Google Scholar] [CrossRef]
- Kutschera, U.; Niklas, K.J. The modern theory of biological evolution: An expanded synthesis. Naturwissenschaften 2004, 91, 255–276. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G. Tempo and Mode in Evolution; Columbia University Press: New York, NY, USA, 1944. [Google Scholar]
- Gould, S.J.; Eldredge, N. Punctuated equilibrium comes of age. Nature 1993, 366, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Gower, B. Scientific Method: An Historical and Philosophical Introduction; Routledge: New York, NY, USA, 1997. [Google Scholar]
- Boring, E.G. The nature and history of experimental control. Am. J. Psychol. 1954, 67, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Mayo, D.G. Error and the Growth of Experimental Knowledge; University of Chicago Press: Chicago, IL, USA, 1996. [Google Scholar]
- Bradbury, S. The Evolution of the Microscope; Pergamon Press: Oxford, UK, 1967. [Google Scholar]
- Croft, W.J. Under the Microscope: A Brief History of Microscopy; World Scientific Publishing: London, UK, 2006. [Google Scholar]
- Collard, P. The Development of Microbiology; Cambridge University Press: Cambridge, UK, 1976. [Google Scholar]
- Laubichler, M.D.; Maienschein, J. From Embryology to Evo-Devo: A History of Developmental Embryology; MIT Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Karamanou, M.; Panayiotakopoulos, G.; Tsoucalas, G.; Kousoulis, A.A.; Androutsos, G. From miasmas to germs: A historical approach to theories of infectious disease transmission. Infez Med. 2012, 20, 58–62. [Google Scholar] [PubMed]
- Pelling, M. The germ theory hypothesized a role for microorganisms in the etiology of infectious diseases and epidemics. Contagion/germ theory/specificity. Companion Encycl. Hist. Med. 1993, 1, 309–334. [Google Scholar]
- Gaynes, R.P. Germ Theory: Medical Pioneers in Infectious Diseases; ASM Press: Washington, DC, USA, 2011. [Google Scholar]
- Santer, M. Confronting Contagion: Our Evolving Understanding of Disease; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Calisher, C.H.; Horzinek, M.C. (Eds.) 100 Year of Virology: The Birth and Growth of a Discipline; Springer: New York, NY, USA, 1999. [Google Scholar]
- Waterson, A.P.; Wilkinson, L. An Introduction to the History of Virology; Cambridge University Press: Cambridge, UK, 1978. [Google Scholar]
- Grafe, A. A History of Experimental Virology; Springer: New York, NY, USA, 2012. [Google Scholar]
- D’hérelle, F. The nature of bacteriophage. Br. Med. J. 1922, 2, 289–297. [Google Scholar]
- Duckworth, D.H. Who discovered bacteriophage? Bacteriol. Rev. 1976, 40, 793–802. [Google Scholar] [PubMed]
- Taylor, M.W. Viruses and Man: A History of Interactions; Springer: New York, NY, USA, 2014. [Google Scholar]
- Hughes, S.S. The virus: A history of the concept. J. Hist. Biol. 1979, 12, 205–206. [Google Scholar]
- Van Helvoort, T. History of virus research in the twentieth century: The problem of conceptual continuity. Hist. Sci. 1994, 32, 185–235. [Google Scholar] [CrossRef] [PubMed]
- Strauss, E.G.; Strauss, J.H. Viral genomic material, in contrast to bacterial genomic material, could be composed of RNA. In Viruses and Human Diseases, 2nd ed.; Elsevier: New York, NY, USA, 2008. [Google Scholar]
- Rubin, H. The early history of tumor virology: Rous, RIF, and RAV. Proc. Natl. Acad. Sci. USA 2011, 108, 14389–14396. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.K. Historical introduction to the general properties of retroviruses. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Press: New York, NY, USA, 1997; pp. 1–25. [Google Scholar]
- Shors, T. Understanding Viruses, 2nd ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2013. [Google Scholar]
- Kurth, R.; Bannert, N. Retroviruses: Molecular Biology, Genomics, and Pathogenesis; Caister Academy Press: Norfolk, UK, 2010. [Google Scholar]
- Marcum, J.A. The transformation of oncology in the twentieth century: The molecularization of cancer. In Proceedings of the 37th International Congress on the History of Medicine; Burns, C.R., O’Neill, Y.V., Albou, P., Rigau-Pérez, J.G., Eds.; University of Texas Medical Branch: Galveston, TX, USA, 2001; pp. 41–49. [Google Scholar]
- Marcum, J.A. From heresy to dogma in accounts of opposition to Howard Temin’s DNA provirus hypothesis. Hist. Philos. Life Sci. 2002, 24, 165–192. [Google Scholar] [CrossRef] [PubMed]
- Mahy, B.M.J. The Dictionary of Virology, 4th ed.; Elsevier: New York, NY, USA, 2009. [Google Scholar]
- Raff, R.A. Evo-devo: The evolution of a new discipline. Nat. Rev. Genet. 2000, 1, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Ruse, M. The Darwinian Revolution: Science Red in Tooth and Claw; University of Chicago Press: Chicago, IL, USA, 1999. [Google Scholar]
- Mayr, E.; Provine, W.B. (Eds.) The Evolutionary Synthesis: Perspectives on the Unification of Biology; Harvard University Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Needham, J.; Hughes, A. A History of Embryology; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Gilbert, S.F. (Ed.) A Conceptual History of Modern Embryology: Volume 7: A Conceptual History of Modern Embryology; Springer: New York, NY, USA, 2013. [Google Scholar]
- Darwin, C. On the Origins of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Amundson, R. The Changing Role of the Embryo in Evolutionary Thought: Roots of Evo-Devo; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Laubichler, M.D.; Wagner, G.P. Introduction to the papers of the 2001 Kowalevsky Medal winner symposium. J. Exp. Zool. (Mol. Dev. Evol.) 2004, 302B, 1–4. [Google Scholar] [CrossRef]
- Moczek, A.P.; Sears, K.E.; Stollewerk, A.; Wittkopp, P.J.; Diggle, P.; Dworkin, I.; Ledon-Rettig, C.; Matus, D.Q.; Roth, S.; Abouheif, E.; et al. The significance and scope of evolutionary developmental biology: A vision for the 21st century. Evol. Dev. 2015, 17, 198–219. [Google Scholar] [CrossRef] [PubMed]
- Sachese, C. Reductionism in the Philosophy of Science; Ontos Verlag: Frankfurt, Germany, 2007. [Google Scholar]
- Kitcher, P. Toward a pragmatist philosophy of science. Theoria 2013, 77, 185–231. [Google Scholar] [CrossRef] [Green Version]
- Dupré, J. The Disorder of Things: Metaphysical Foundations of the Disunity of Science; Harvard University Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Galison, P.L.; Stump, D.J. (Eds.) The Disunity of Science: Boundaries, Contexts, and Power; Stanford University Press: Stanford, CA, USA, 1996. [Google Scholar]
- Ruphy, S. Scientific Pluralism Reconsidered: A New Approach to the (Dis)Unity of Science; University of Pittsburgh Press: Pittsburgh, PA, USA, 2013. [Google Scholar]
- Kuhn had asked a similar question when discussing the appropriateness of evolutionary theory for understanding science: “What must nature, including man, be like in order that science should be possible at all?” [4] (p. 173). Although he did not answer this question fully, even when articulating his EPoS, the question is central to establishing EPoS as a GPoS. In other words, the natural sciences are an adaptation of Homo sapiens sapiens to its environment and only the evolving of these sciences can determine whether they lead to the species’ survival or extinction.
- Metacognitive evolution refers to an ability to step back and evaluate how well cognitive, especially scientific, processes represent the world. In a very real sense, the proposed EPoS represents such a metacognitive process. In other words, if the proposed EPoS allows for a better evaluation of scientific cognitive processes, then it opens up the possibility of enhancing the overall representation of the nature in terms of scientific practice and knowledge.
- Gillett, C. Reduction and Emergence in Science and Philosophy; Cambridge University Press: New York, NY, USA, 2016. [Google Scholar]


© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcum, J.A. Evolutionary Philosophy of Science: A New Image of Science and Stance towards General Philosophy of Science. Philosophies 2017, 2, 25. https://doi.org/10.3390/philosophies2040025
Marcum JA. Evolutionary Philosophy of Science: A New Image of Science and Stance towards General Philosophy of Science. Philosophies. 2017; 2(4):25. https://doi.org/10.3390/philosophies2040025
Chicago/Turabian StyleMarcum, James A. 2017. "Evolutionary Philosophy of Science: A New Image of Science and Stance towards General Philosophy of Science" Philosophies 2, no. 4: 25. https://doi.org/10.3390/philosophies2040025
APA StyleMarcum, J. A. (2017). Evolutionary Philosophy of Science: A New Image of Science and Stance towards General Philosophy of Science. Philosophies, 2(4), 25. https://doi.org/10.3390/philosophies2040025