1. Introduction
Certain types of aquaculture disrupt various marine ecosystems, in no small part due to the use of wild pelagic fish, e.g., sardines and anchovies, to produce fish meal (FM), particularly when farming marine carnivorous fishes [
1]. Due to a rapid expansion of global aquaculture production, the development of alternative protein sources in feed for carnivorous fishes has become quite an urgent issue. Insects have attracted broad attention as a novel protein source for enhancing global food security since the Food and Agriculture Organization (FAO) published an assessment of edible insects as food and feed [
2]. Similarly, the replacement of FM with insects in the feed for ornamental aquaculture was reported [
3].
The housefly (
Musca domestica) can yield animal protein in the biodegradation of organic waste [
4], such as pig manure [
5,
6,
7,
8,
9,
10], cattle manure [
11], poultry litter [
12], and food waste [
13]. The larvae obtained from these systems are alternative feed ingredients that are rich in nutrients from within the human food chain without the need to harvest part of the wild marine ecosystem.
Although a number of studies have now reported on FM replacement in fish diets using housefly (HF) larvae, most of the studies were demonstrated with mainly freshwater-fish species, such as Nile tilapia (
Oreochromis niloticus) and catfish species (
Clarias gariepinus and
Heterobranchus longifil) [
14,
15]. Another study demonstrated that a 30% inclusion of HF larvae meal in a diet was possible in barramundi (
Lates calcarifer), but the authors concluded that up to 10% inclusion was recommended due to the physiological response in cultured fish caused by chitin, an imbalanced fatty acid profile (high
n−6 fatty acids), or another unknown factor contained in HF larvae [
16]. Substitution of FM with HF larvae seems to be limited, especially for marine carnivorous fish species, which have a high dietary protein requirement.
We previously reported that the low inclusion of HF (0.5–5%) in diets promoted growth and disease resistance for the marine carnivorous fish red seabream (
Pagrus major) [
17]; however, 25% or more inclusion of HF (50% or more replacement of FM) in diets resulted in growth reduction in red seabream (data not shown). While HF larvae appear to possess an adequate amino acid profile, similar to FM, the lack in high unsaturated omega−3 fatty acids was noted as a factor for the limit in their inclusion [
18,
19]. A negative factor in HF larvae for the growth of marine fish should be studied in detail for the establishment of sustainable aquaculture.
Insects are known to contain a variety of unique hydrophobic molecules [
20]. One of these hydrophobic molecules that are unique to insects, catechol and its derivatives, which are known to be abundant in the cuticle, play a central role in the sclerotization and melanization by covalently inducing protein–protein and protein–chitin crosslinks [
21,
22]. An isomer and derivatives of catechol have been reported to exhibit acute toxicity in fish [
23,
24]. Although the dietary inclusion of the black soldier fly (
Hermetia illucens) larvae was reported to affect enzymatic activities [
25] and intestinal microbiota [
26] in the laying hen, the dietary effect of these kinds of molecules from insects on cultured fish growth and their intestines remains unknown.
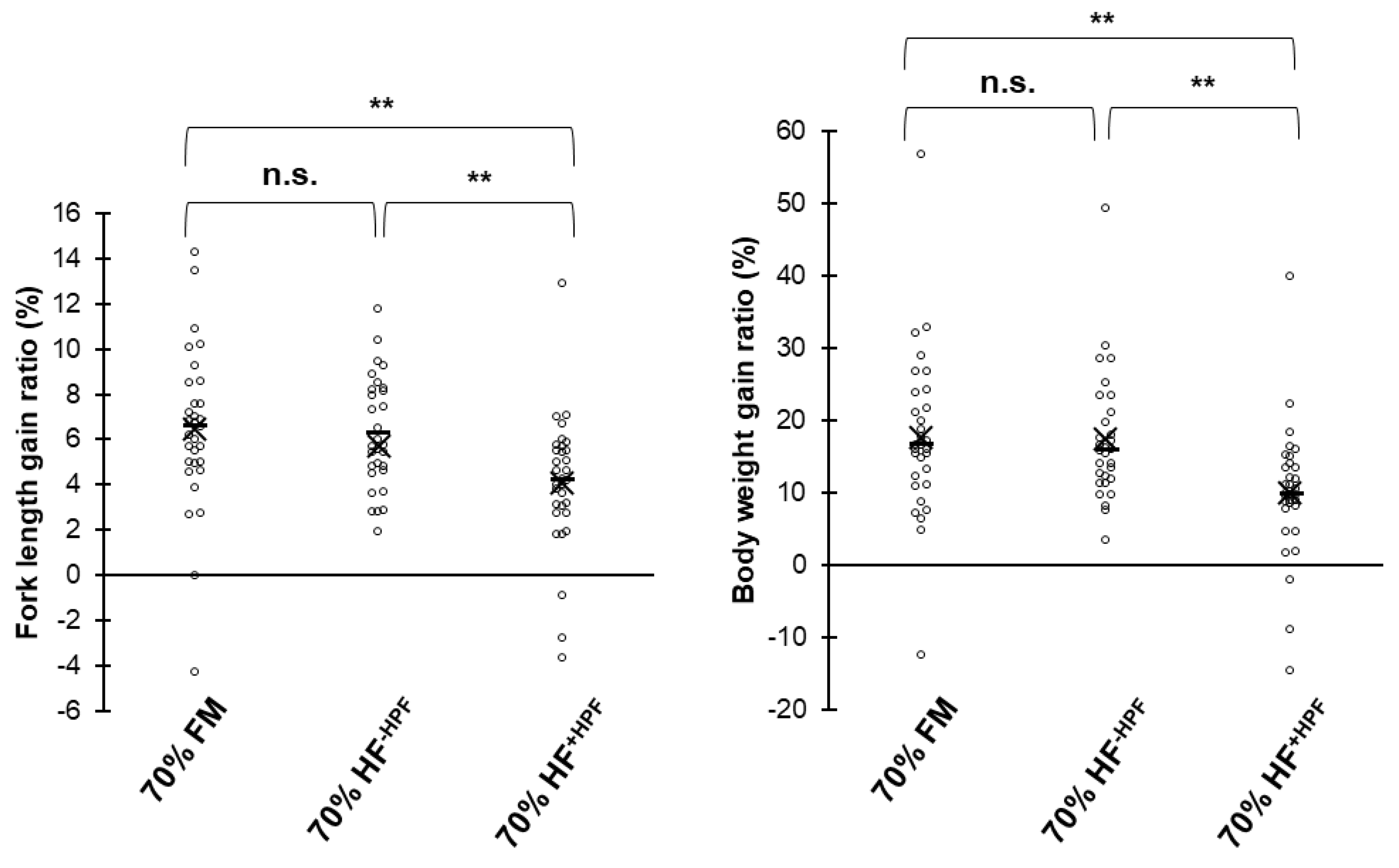
In the current study, we report that the removal of the hydrophobic fraction from HF larvae enables more significant replacement of FM in the diet of the red seabream. Moreover, the free catechol involved in insect cuticular was detected in the hydrophobic fraction from HF larvae. Although no significant difference in growth was observed with the low dietary level of catechol, morphology alterations were observed in the intestine of red seabream. This study indicates that catechol in HF larvae is thought to be one of the negative factors causing growth reduction in cultured marine fish.
3. Discussion
We found in our present study that FM can be successfully replaced with HF larvae in an aquaculture feed far beyond the 25% level if the hydrophobic fraction is removed with an organic-solvent elimination process. A large amount of FM in diets could be replaced with HF larvae preparation after the removal of hydrophobic fractions, and our findings show the potential of HF larvae as an alternative feed ingredient for sustainable aquaculture production. Although the cost of the hydrophobic fraction removal must be considered, the purchasing price of dried HF larvae produced from swine manure in a full-scale commercial system in China was reported to be
$1.25 to US
$1.43 kg
−1 [
8]; the annual average of the FM price in 2018 was US
$1.49 kg
−1 [
30]; therefore, HF larvae could economically substitute FM. The amino acid profile in HF larvae was comparable with that of FM except for taurine. Lack of dietary taurine resulted in green-liver syndrome and growth retardation [
31,
32,
33]. Hence, we determined that HF larvae are a potentially viable protein source for farmed fish in their amino acids, but that supplementation with taurine would be required for their use in aquaculture feeds.
With respect to fatty acids, n−3 high unsaturated fatty acids (HUFAs) are absent in HF larvae but could be added as a supplement to the diet. However, recovery of fish growth was partial, and improvements in BW gain and feed intake were insufficient. A substantial portion (12.4%) of total fatty acids were not identified in this study. Unidentified fatty acids in HF larvae or pupae were relatively high (7.2–8.5%) comparing with other insects [
19]. Since the composition of unidentified fatty acids in HF larvae reached 15% and depended on sources of poultry litter as rearing substrates for HF larvae [
12], these unidentified fatty acids seemed to be derived from pig manure used for rearing the larvae. Chitin, a linear homopolymer of β(1-4)-linked
N-acetylglucosamine units, a major constituent of insect cuticles, was thought to influence growth reduction in marine fish because of its difficulty in digestion [
34]. Although 3.0–6.8% nitrogen of the total nitrogen was thought to be chitin in insect larvae [
35], chitin in diets could be digested and utilized in red seabream, as the growth of red seabream was improved when diets were supplemented with 10% chitin [
36]. Chitinase activity in fish stomach is known to degrade chitin efficiently [
37]. Therefore, HF larvae in diets may contain other factors to prevent growth in the red seabream because chitin from HF larvae could be utilized by the red seabream.
Insects are known to contain a variety of unique hydrophobic molecules [
20]. These molecules in the hydrophobic fractions of HF larvae, other than the nutritional deficiency caused by the absence of n-3 HUFAs, could also participate in growth reduction. Since the dietary intake of a hydrophobic fraction from yellow-mealworm (
Tenebrio molitor) larvae resulted in significant growth reduction in red seabream [
38], this property could be observed not only in HF larvae but in a wide range of insect species. One explanation for the reduced growth is the involvement of insect cuticular molecules, such as straight or methyl-branched n-alkanes, n-alkenes, fatty acids, alcohols, wax esters [
39,
40], and catechol and its derivatives [
22,
23] in the hydrophobic fractions. In particular, hydroquinone (1, 3-benzendiol), a structural isomer of catechol, was reported to exhibit acute toxicity in fathead minnow (
Pimephales promelas) and rainbow trout (
Oncorhynchus mykiss) [
23]; toxicity of catechol derivatives, such as dichlorocatechol, was also known in brown trout (
Salmo trutta) [
24] in exposure assessments. The mechanisms of their toxicity were thought to be the generation of reactive oxygen species (ROS), DNA and protein damage, or interference with electron transport in energy-transducing membranes [
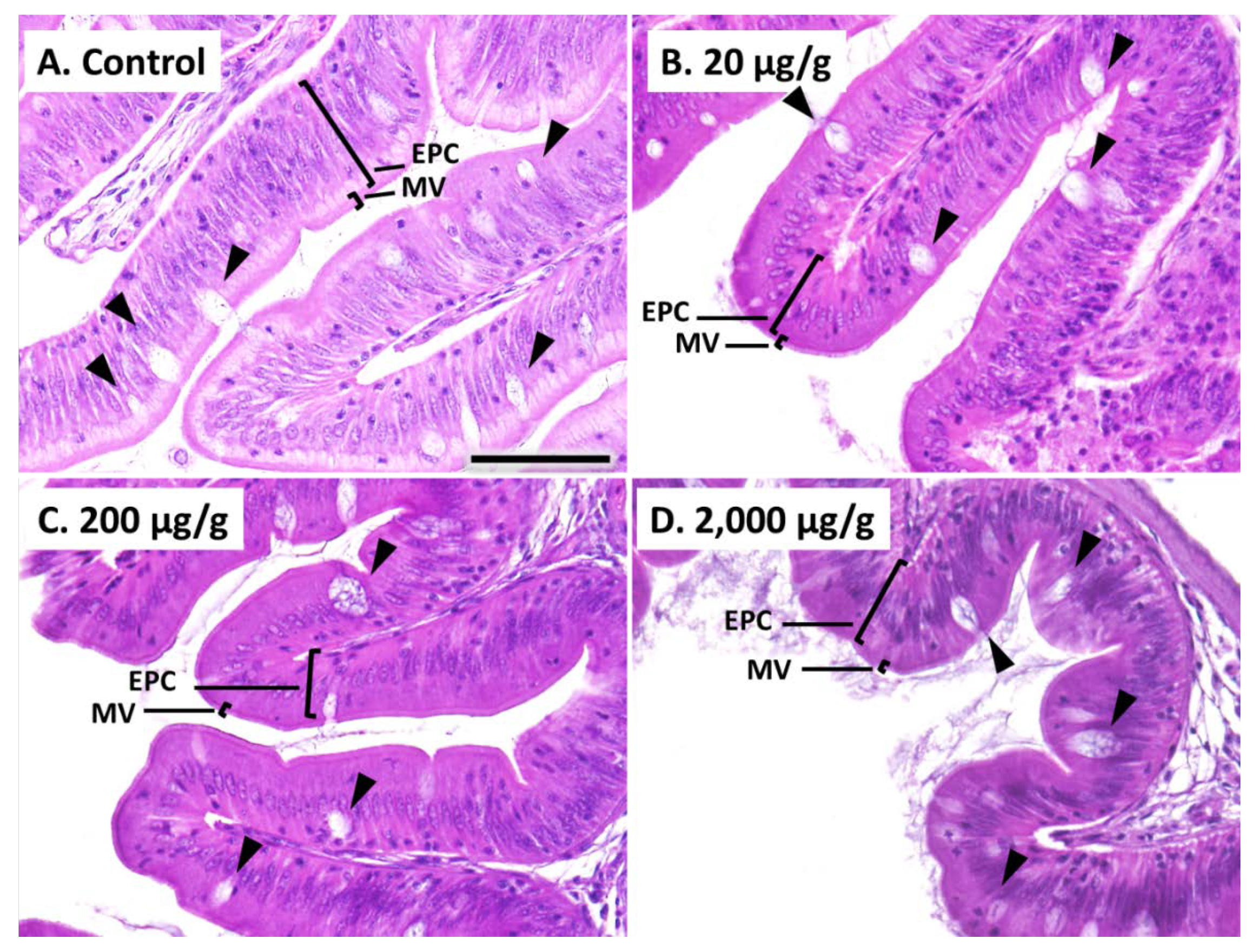
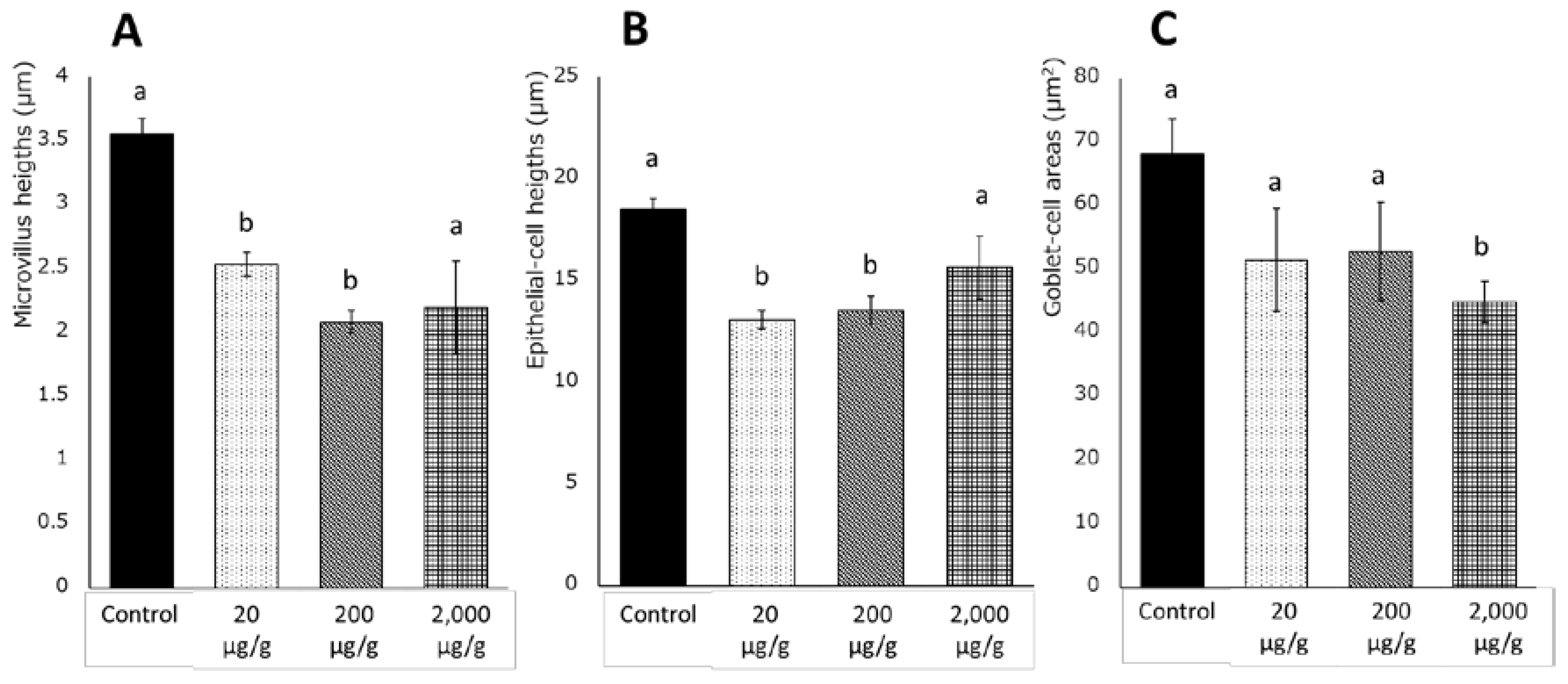
41]. In the current study, red seabreams with a dietary intake of catechol showed intestinal morphological alterations at lower doses and growth reductions at higher doses. These findings suggest that catechol in the hydrophobic fraction of HF larvae causes incomplete growth recovery despite supplementation with DHA and EPA in diets. In HF larvae, 27.1 μg/g catechol was detected before the removal of hydrophobic fractions, and approximately 20 μg/g catechol was estimated to be contained in the 70% HF
+HPF diet. Since no significant difference in growth after four weeks of feeding was observed with the addition of 20 μg/g and 200 μg/g catechol, dietary intake of other hydrophobic molecules than catechol in the fractions of HF larvae seems to affect fish growth. Nevertheless, regression of microvillus, intestinal epithelium cells, and goblet cells, which have important roles in nutrition absorption, were observed in the intestine of red seabream with the dietary intake of 20 μg/g catechol; thus, these results imply that strain on the intestine caused by dietary catechol even at low levels resulted in partial growth reduction. As we showed in the previous study, the diet with a low inclusion of HF gave disease resistance in red seabream [
17]. In contrast, since intestinal epithelial cells in fish are related to innate immunity, and the recognition of pathogens [
42], morphological alterations in the intestine by the dietary intake of catechol might provoke infectious diseases. Although further observation of the intestine with another method, such as Fourier Transform Infrared (FTIR) spectroscopy [
43] is needed, removal of the hydrophobic fraction including catechol was thought to be an essential process for the high inclusion of HF larvae in diets.
Importantly, without defatting, it is not easy to use HF larvae in a marine-fish diet and obtain appropriate crude protein and lipid composition, since HF larvae have an excess of crude lipids (20–35%, dry matter) and markedly different fatty acid profiles than feeds made from wild fish. The fatty acid profile in black-soldier fly larvae (
Hermetia illucens) was reported to affect lipid composition and accumulation in zebrafish (
Danio rerio) [
44,
45]. Eliminating the hydrophobic faction is, therefore, an important first step in the development of HF larva-based feeds for marine-fish aquaculture.
Long-term feeding trials of different HF larva formulations are still needed to further evaluate the growth performance and safety of the hydrophobic fraction of HF larvae in farmed fish. Other bioactive molecules than catechol in the hydrophobic fraction should be studied, and the upper limits of the HF larvae of fish feed after the removal of the hydrophobic fraction should also be elucidated in other marine-fish species. Furthermore, the development of defatting methods that do not require organic solvents is desirable for practical reasons. However, our present study provides some key new findings; the replacement of FM with HF larvae in diets for marine carnivorous fish like red seabream is a potentially viable option that could lead to a more environmentally sustainable aquaculture industry in the future.
4. Materials and Methods
4.1. Feed Ingredients
HF larvae were reared with pig manure by E’s Inc. (Tokyo, Japan), boiled for 15 min and frozen at –30 °C until use. HF larvae were ground in a mill mixer after air-drying at 80 °C for 5 h or more. To defat the HF larvae, the ground HF larvae were treated twice with a four-fold volume of normal hexane + ethanol (9:1) for 2 h at room temperature. This defatting process was repeated several times until the crude lipids reached below 10% (
Table 7). The supernatant in the defatting process was removed after centrifugation, and the sediment was air-dried at 60 °C for one day or more until completely air-dried. The HF hydrophobic fractions (HPF) were prepared from the aforementioned supernatant containing hexane + ethanol (9:1) by evaporation at room temperature until the volume was decreased by about 10-fold; then the temperature was gradually increased from 40 to 80 °C until the residual organic solvents had evaporated. FM from Peruvian anchoveta (
Engraulis ringens) comprising 65% crude protein and cod fish oil were obtained from the Shintoa Corporation (Tokyo, Japan). DHA and EPA were kindly provided by Bizen Chemical Co., Ltd. (Akaiwa, Okayama, Japan).
4.2. Experimental Diets
We tried the complete replacement of 70% FM and 40% FM with HF larvae. In the first trial, a control diet included 70% FM, and FM in the diet was replaced with 70% defatted HF larvae (100% replacement of FM). Fish oil was added at 7% to the control diet and HF diet (HF
−HPF), and the hydrophobic fraction from HF was added to another HF diet (HF
+HPF) as an alternative to fish oil (
Table 8). In the second study, the control diet included 40% FM and 11% fish oil, and the FM in the diet was replaced with 40% defatted HF larvae and fish oil (HF
−HPF). We produced two more diets where 40% FM in the control diet was replaced with undefatted HF larvae, and we added DHA and EPA to the one diet to be equivalent to n-3 HUFA content levels among the control, HF
−HPF, and undefatted HF
+DHA+EPA (
Table 9). Crude protein in all these experimental diets was targeted to be around 50% to obtain the satisfactory growth of red seabream [
46]. Supplementation with plant-derived protein sources (soybean meal and corn gluten meal) was done to achieve this protein level. To prepare dried pellets for use in the feeding trials, the powdered forms of all components were thoroughly mixed and supplemented with fish oil. These mixtures were then granulated after adding water and air-dried at 60 °C for more than one day. The resulting dry pellets were stored at 4 °C until use. In the feeding trial for the evaluation of catechol, catechol (Nakalai Tesque, Kyoto, Japan; ≥99.0% GC assay) was added to commercially available extruded pellet diets for juvenile red seabream Otohime
TM (Marubeni Nisshin Feed Co., Ltd., Tokyo, Japan). Catechol was added to the diets at 20, 200, and 2000 μg/g, and the surface of the diets was coated with plant-derived oil.
4.3. Feeding Trials
Animal experiments were carried out in accordance with the guidelines at Ehime University. The study protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of Ehime University (Permit Number: 3908). All body measurements and surgical manipulations were performed under anesthesia with 0.5–1 mL/L 2-phenoxyethanol, and all efforts were made to minimize suffering. Juvenile red seabreams were kindly provided by Yamasaki Giken Co., Ltd. (Susaki, Kochi, Japan), and had been maintained in 1000 L tanks containing natural sea water on a flow-through system with sand filtration. Fish under one year of age with no traumatic injuries or malformations were included in the trials. Unique identification tags were intraperitoneally injected into the fish bodies, and the animals were allocated into different feeding study groups of 22–40 fish. Two tanks per study group containing half of the fish were cultivated in 100 or 200 L round tanks, depending on availability at the time of the trial. In the trials, fish were fed once per day except for Sunday with the test diets by satiation. The water temperature was 14–18 °C in the 70% FM/HF trial, 15–20 °C in 40% FM/HF Trial 1, and 27–29 °C in 40% FM/HF Trial 2. The main trial with the 70% FM/HF diets was initiated after preliminary feeding with subject diets for two weeks. In the feeding trial for the evaluation of catechol, juvenile red seabreams provided by Bio Ehime Co. Ltd. (Imabari, Ehime, Japan) were used. Thirty fish were cultured in a 200 L round tank, and the water temperature was 17–20 °C. All trials were conducted over a one month period. FL and BW were measured every two weeks.
BW gain, BW gain rate, total feed intake per fish, and feed conversion ratio were calculated as follows:
FL gain (mm) = FL at trial end—initial FL,
BW gain (g) = BW at trial end—initial BW,
FL gain percentage (%) = FL gain/initial FL × 100,
BW gain percentage (%) = BW gain/initial BW × 100,
Feed intake per fish (g) = total feed intake per group (g)/number of fish,
Feed conversion ratio = total feed intake per group (g)/BW gain per group.
Measurements in individual fish were used to obtain the FL, BW, FL gain, BW gain, FL gain rate, and BW gain rate in the feeding test groups, and values in the duplicate tanks were used to obtain the total feed intake per fish and feed conversion ratio in each study group.
4.4. Histological Observations
At the end of the feeding trial for the evaluation of catechol, intestines excised from red seabream were fixed in Davidson’s solution and embedded in paraffin wax. Sections were cut at 5 μm thickness and stained with hematoxylin and eosin. Morphological differences were evaluated with measurement of microvillus heights, epithelial-cell heights, and goblet-cell areas using a phase-contrast microscope (BX51N-33-PH, Olympus, Tokyo, Japan), and ImageJ (National Institutes of Health, Bethesda, MD, USA). Five heights or areas in each intestine from five fish per group were measured (n = 25/group).
4.5. Analysis of Free Catechol in Housefly Larvae
To analyze the free catechol in HF larvae, 0.5 g of samples was mixed well in 1 mL water. Subsequently, 5 mL of acetone was added, and samples were incubated for 30 min at room temperature. The collected supernatants after centrifugation were analyzed by GC/MS analysis using the Agilent 7890 gas chromatography system (Agilent Technologies, Santa Clara, CA, USA) equipped with a CP-Sil8-CB capillary column (30 m × 0.25 mm × 0.25 μm; Agilent Technologies, Santa Clara, CA, USA) connected to a JMS-Q1500 mass spectrometer (JEOL, Tokyo, Japan). Helium was used as the carrier gas at a constant flow of 1.2 mL/min. Column temperature conditions were as follows: 100 °C (1 min) −10 °C/min − 200 °C (10 min) – 40 °C/min −300 °C (10 min). MS parameters for m/z 63, 64, 81, 92, and 110 were obtained using SIM mode and the electron-ionization method. Pyrocatechol (Sigma-Aldrich, St. Louis, MO, USA) was used as the standard.
4.6. Proximate-Composition, Amino Acid, and Fatty Acid Analysis
The proximate composition, amino acids, and fatty acids were analyzed using the Association of Analytical Communities (AOAC) methods [
47]. The content of crude protein was analyzed using the Kjeldahl method. “Kjeltab” (containing K
2SO
4) was added to the ground samples, and samples were digested in a block heater (Tecator TM Digestion Systems 2520, FOSS, Hillerød, Denmark). Nitrogen content was automatically analyzed using an autoanalyzer (Kjeltec
TM 8400, FOSS, Hillerød, Denmark). The nitrogen–protein conversion factor was 6.25 for the calculation of crude protein from the nitrogen content. Crude fat was analyzed using the Soxhlet extraction method. Petroleum ether was used as the solvent for extraction from the ground samples in an automated extractor (Soxtec
TM 8000, FOSS, Hillerød, Denmark). Ash was analyzed using an electric furnace (MMF-1, AS ONE, Osaka, Japan).
Proteinogenic amino acids in the meal samples were analyzed using an automated amino acid analyzer (Shimadzu, Kyoto, Japan) after hydrochloric hydrolysis with sodium chloride. For methionine and cystine, samples were oxidized with performic acid prior to hydrochloric hydrolysis. For tryptophan, samples were prepared with barium hydroxide octahydrate and thiodietylene glycol before hydrolysis with sodium chloride and analyzed with high-performance liquid chromatography. Taurine was measured using high-performance liquid chromatography. Fatty acids in samples were prepared with hydrolysis and extracted with diethyl ether-petroleum ether (1:1). Gas chromatograph GC-2010 (Shimadzu Corporation, Kyoto, Japan) and Agilent J&W column DB-23 (Agilent Technologies Inc., Santa Clara, CA, USA) were used for analysis with a method described in food-labelling standards by the Consumer Affairs Agency of Japan (CAA, 2015). Fatty acids were identified with Supelco 37 Component FAME Mix (Sigma-Aldrich Co. LLC, St. Louis, MO, USA). Analysis of proteinogenic amino acids, taurine, and fatty acids was conducted by the Japan Food Research Laboratories (Osaka, Japan).
4.7. Statistical Analysis
The statistical methods and
p-values in the present study are shown in the footnotes of the figures and tables. All tests were conducted with the “R” software (
https://www.r-project.org).

 ,
, 




{kind=link}
{kind=link}
{kind=link}