
Slight Increases in Salinity Improve Muscle Quality of Grass Carp (Ctenopharyngodon idellus)


 , and
, and 
Abstract
:1. Introduction
2. Results
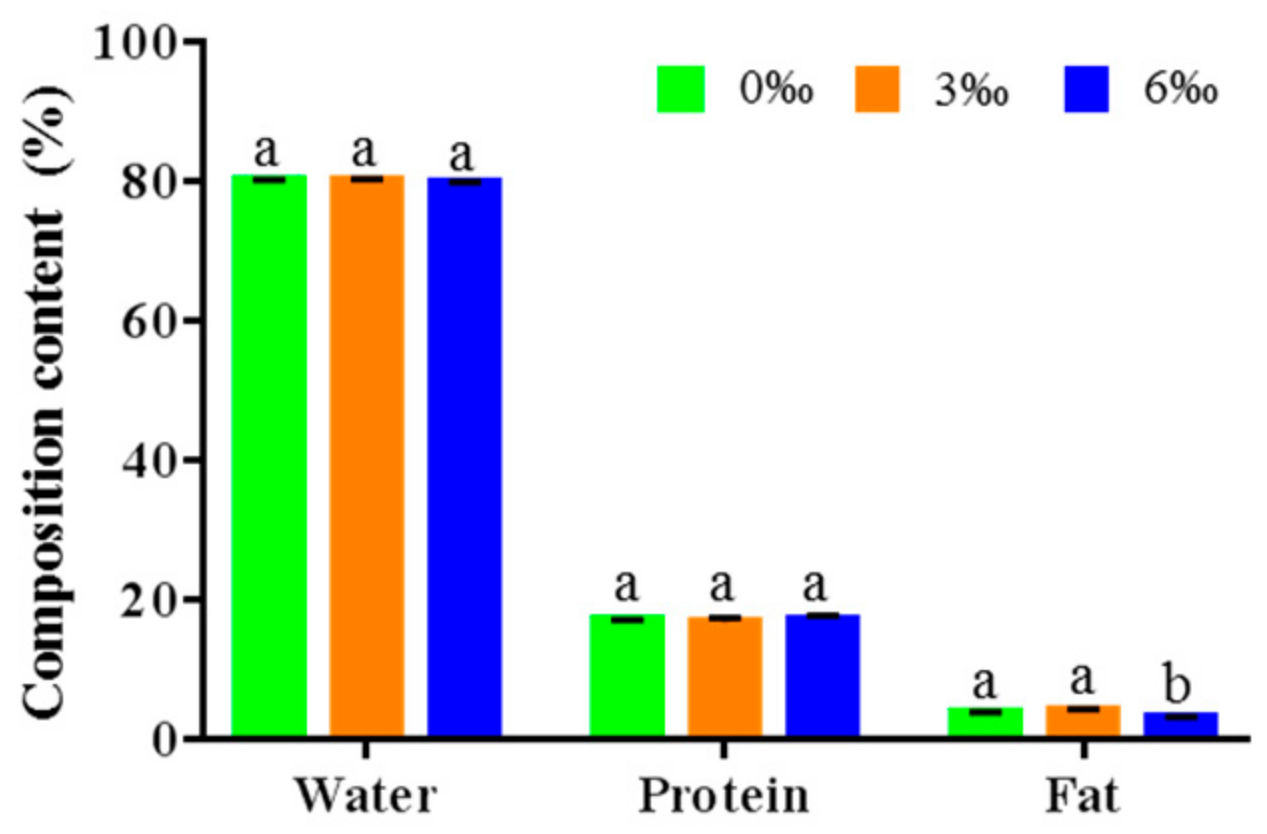
2.1. Chemical Composition
2.2. Amino-Acid Analysis
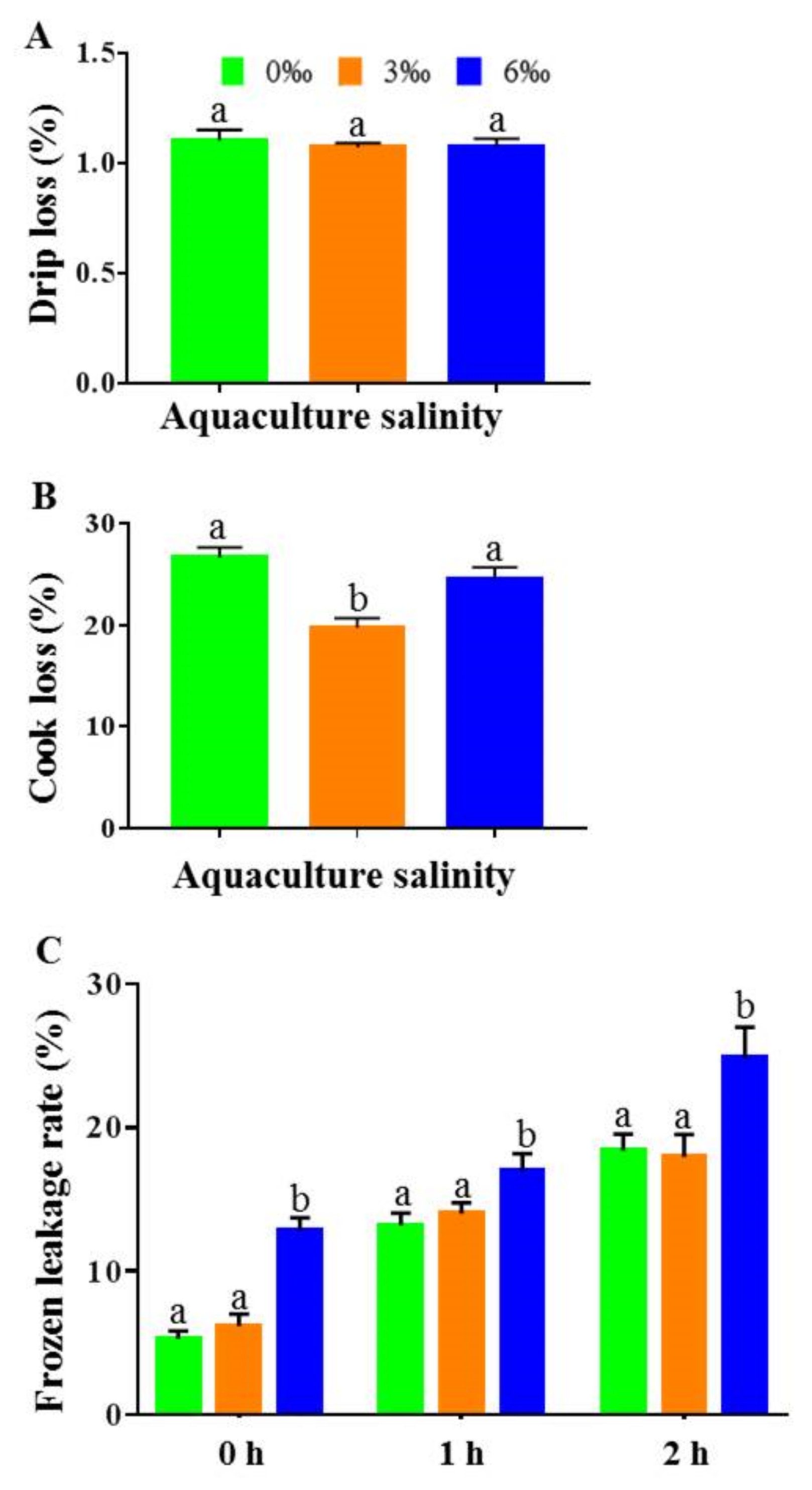
2.3. WHC
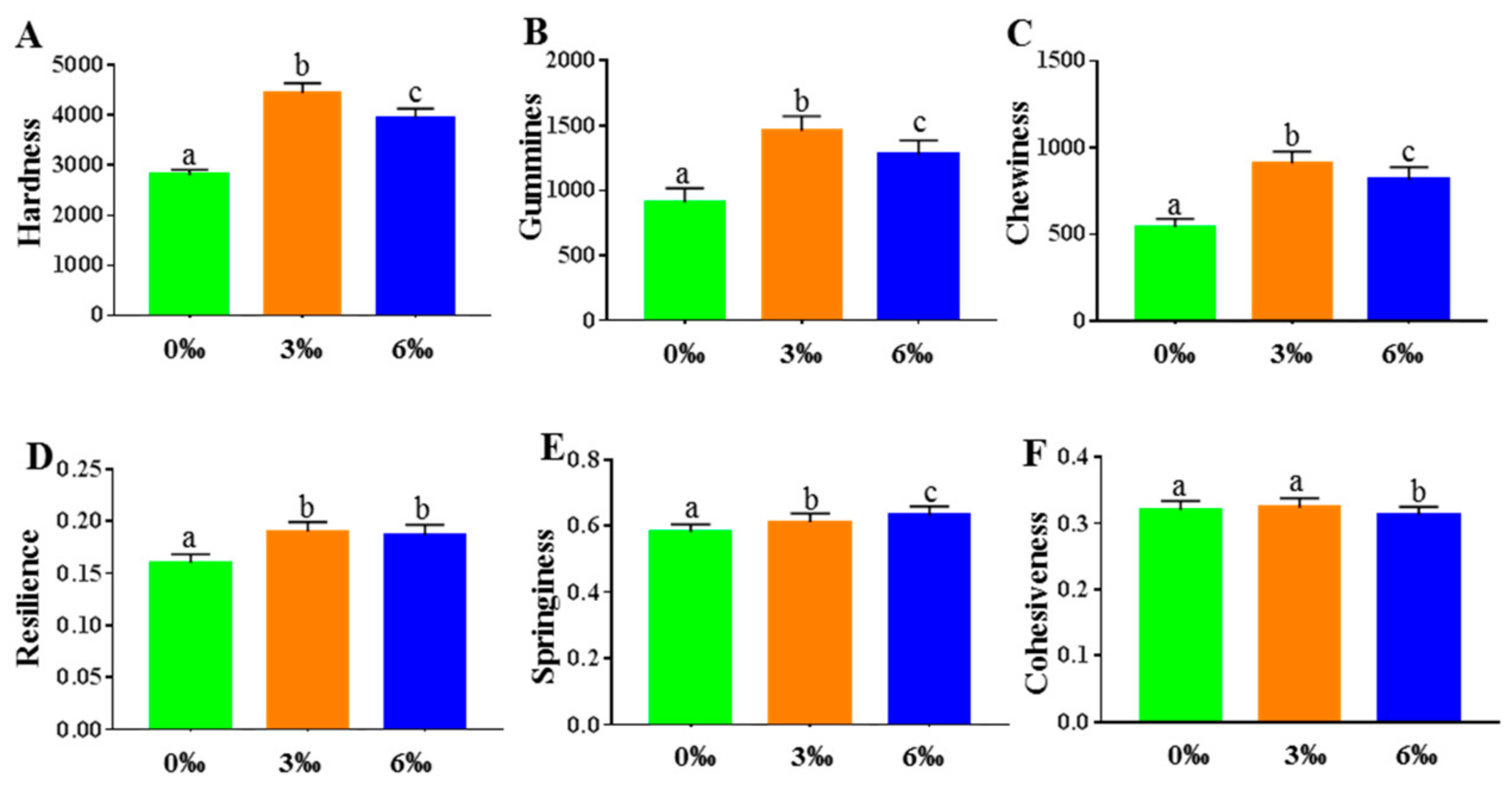
2.4. Analysis of Muscle-Texture Profiles
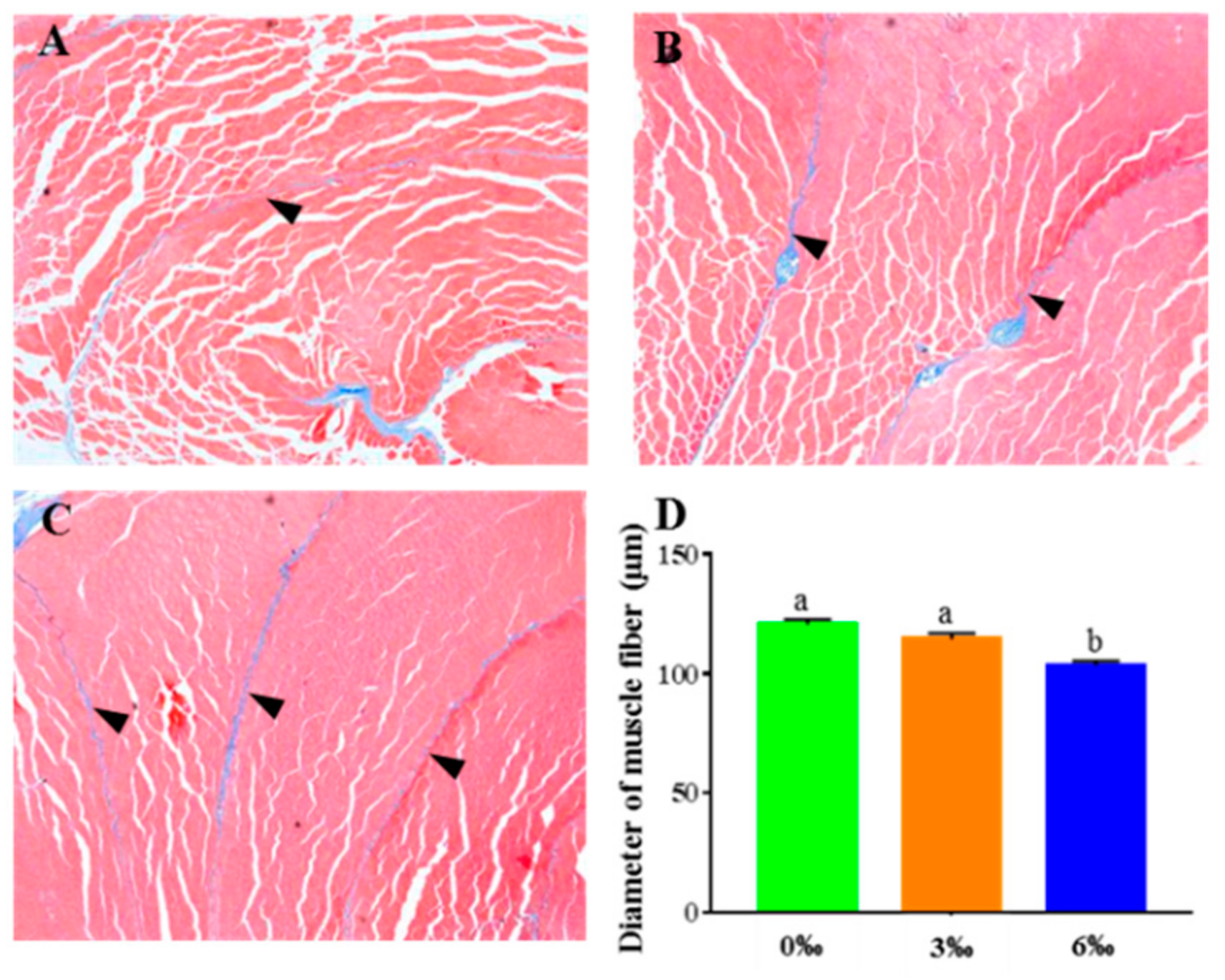
2.5. Histological Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Protocol and Sampling
4.2. Chemical Composition Measurements
4.3. Amino Acid Composition Measurements
4.4. WHC Measurements
4.5. Texture Measurements
4.6. Histological Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Chen, L.; Liu, J.; Kaneko, G.; Xie, J.; Wang, G.; Yu, D.; Li, Z.; Ma, L.; Qi, D.; Tian, J.; et al. Quantitative phosphoproteomic analysis of soft and firm grass carp muscle. Food Chem. 2020, 303, 125367. [Google Scholar] [CrossRef]
- Xie, C.; Li, J.; Li, D.; Shen, Y.; Gao, Y.; Zhang, Z. Grass Carp: The Fish that Feeds Half of China. Aquac. China 2018, 93–115. [Google Scholar] [CrossRef]
- Yu, E.; Fu, B.; Wang, G.; Li, Z.; Ye, D.; Jiang, Y.; Ji, H.; Wang, X.; Yu, D.; Ehsan, H.; et al. Proteomic and metabolomic basis for improved textural quality in crisp grass carp (Ctenopharyngodon idellus C.et V) fed with a natural dietary pro-oxidant. Food Chem. 2020, 325, 126906. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Tang, R.; He, X.; Li, L.; Takagi, Y.; Li, D. Improvement of Muscle Quality of Grass Carp (Ctenopharyngodon idellus) With a Bio-Floating Bed in Culture Ponds. Front. Physiol. 2019, 10, 683. [Google Scholar] [CrossRef]
- Zhao, H.; Xia, J.; Zhang, X.; He, X.; Li, L.; Tang, R.; Chi, W.; Li, D. Diet Affects Muscle Quality and Growth Traits of Grass Carp (Ctenopharyngodon idellus): A Comparison between Grass and Artificial Feed. Front. Physiol. 2018, 9, 283. [Google Scholar] [CrossRef]
- De Paula, T.G.; De Almeida, F.L.A.; Carani, F.R.; Vechetti-Júnior, I.J.; Padovani, C.R.; Salomão, R.A.S.; Mareco, E.A.; Dos Santos, V.B.; Dal-Pai-Silva, M. Rearing temperature induces changes in muscle growth and gene expression in juvenile pacu (Piaractus mesopotamicus). Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2014, 169, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Johnston, I.A. Genetic and Environmental Determinants of Muscle Growth Patterns. Fish Physiology 2001, 18, 141–186. [Google Scholar] [CrossRef]
- Ma, X.-Z.; Feng, L.; Wu, P.; Liu, Y.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q.; Jiang, W.-D. Enhancement of flavor and healthcare substances, mouthfeel parameters and collagen synthesis in the muscle of on-growing grass carp (Ctenopharyngodon idella) fed with graded levels of glutamine. Aquaculture 2020, 528, 735486. [Google Scholar] [CrossRef]
- Chand, B.; Trivedi, R.; Dubey, S.; Rout, S.; Beg, M.; Das, U. Effect of salinity on survival and growth of giant freshwater prawn Macrobrachium rosenbergii (de Man). Aquac. Rep. 2015, 2, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.K.; Trivedi, R.K.; Chand, B.K.; Mandal, B.; Rout, S.K. Farmers’ perceptions of climate change, impacts on freshwater aquaculture and adaptation strategies in climatic change hotspots: A case of the Indian Sundarban delta. Environ. Dev. 2017, 21, 38–51. [Google Scholar] [CrossRef]
- Kilambi, R.V. Food consumption, growth and survival of grass carp Ctenopharyngodon idella Val at four salinities. J. Fish Biol. 1980, 17, 613–618. [Google Scholar] [CrossRef]
- Martin, R. Contaminants in relation to the quality of seafoods. Food Technol. 1998, 42, 104. [Google Scholar]
- Alasalvar, C.; Taylor, K.; Zubcov, E.; Shahidi, F.; Alexis, M. Differentiation of cultured and wild sea bass (Dicentrarchus labrax): Total lipid content, fatty acid and trace mineral composition. Food Chem. 2002, 79, 145–150. [Google Scholar] [CrossRef]
- Cheng, J.-H.; Sun, D.-W.; Han, Z.; Zeng, X.-A. Texture and Structure Measurements and Analyses for Evaluation of Fish and Fillet Freshness Quality: A Review. Compr. Rev. Food Sci. Food Saf. 2013, 13, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.; Fernández-Segovia, I.; Serra, J.A.; Barat, J.M. Comparison of wild and cultured sea bass (Dicentrarchus labrax) quality. Food Chem. 2010, 119, 1514–1518. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.; Shahidi, F.; Whittick, A.; Hooper, R. Effect of season and artificial diet on amino acids and nucleic acids in gonads of green sea urchin Strongylocentrotus droebachiensis. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2002, 133, 389–398. [Google Scholar] [CrossRef]
- Hyldig, G.; Nielsen, D. A Review of Sensory and Instrumental Methods Used to Evaluate the Texture of Fish Muscle. J. Texture Stud. 2001, 32, 219–242. [Google Scholar] [CrossRef]
- Palmeri, G.; Turchini, G.M.; Caprino, F.; Keast, R.; Moretti, V.M.; De Silva, S.S. Biometric, nutritional and sensory changes in intensively farmed Murray cod (Maccullochella peelii peelii, Mitchell) following different purging times. Food Chem. 2008, 107, 1605–1615. [Google Scholar] [CrossRef]
- Du, H.; Lv, H.; Xu, Z.; Zhao, S.; Huang, T.; Manyande, A.; Xiong, S. The mechanism for improving the flesh quality of grass carp (Ctenopharyngodon idella) following the micro-flowing water treatment using a UPLC-QTOF/MS based metabolomics method. Food Chem. 2020, 327, 126777. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Hu, W.; Xiong, S.; You, J.; Fan, Q. Depuration and starvation improves flesh quality of grass carp (Ctenopharyngodon idella). Aquac. Res. 2018, 49, 3196–3206. [Google Scholar] [CrossRef]
- Menezes, C.; Ruiz-Jarabo, I.; Martos-Sitcha, J.A.; Toni, C.; Salbego, J.; Becker, A.; Loro, V.L.; Martínez-Rodríguez, G.; Mancera, J.M.; Baldisserotto, B. The influence of stocking density and food deprivation in silver catfish (Rhamdia quelen): A metabolic and endocrine approach. Aquaculture 2015, 435, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Montero, D.; Izquierdo, M.; Tort, L.; Robaina, L.; Vergara, J. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Andersen, U.B.; Thomassen, M.S.; Rørå, A.M.B. Texture Properties of Farmed Rainbow Trout (Oncorhynchus mykiss): Effects of Diet, Muscle Fat Content and Time of Storage on Ice. J. Sci. Food Agric. 1997, 74, 347–353. [Google Scholar] [CrossRef]
- Otto, G.; Roehe, R.; Looft, H.; Thoelking, L.; Henning, M.; Plastow, G.; Kalm, E. Drip loss of case-ready meat and of premium cuts and their associations with earlier measured sample drip loss, meat quality and carcass traits in pigs. Meat Sci. 2006, 72, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Huff-Lonergan, E.; Lonergan, S.M. Mechanisms of water-holding capacity of meat: The role of postmortem biochemical and structural changes. Meat Sci. 2005, 71, 194–204. [Google Scholar] [CrossRef]
- Bee, G.; Anderson, A.L.; Lonergan, S.M.; Huff-Lonergan, E. Rate and extent of pH decline affect proteolysis of cytoskeletal proteins and water-holding capacity in pork. Meat Sci. 2007, 76, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Nathanailides, C.; Lopez-Albors, O.; Abellán, E.; Vazquez, J.M.; Tyler, D.D.; Rowlerson, A.; Stickland, N.C. Muscle cellularity in relation to somatic growth in the European sea bass Dicentrarchus labrax (L.). Aquac. Res. 1996, 27, 885–889. [Google Scholar] [CrossRef]
- Periago, M.; Ayala, M.; López-Albors, O.; Abdel, I.; Martínez, C.; García-Alcázar, A.; Ros, G.; Gil, F. Muscle cellularity and flesh quality of wild and farmed sea bass, Dicentrarchus labrax L. Aquaculture 2005, 249, 175–188. [Google Scholar] [CrossRef]
- Lin, W.L.; Zeng, Q.X.; Zhu, Z.W. Different changes in mastication between crisp grass carp (Ctenopharyngodon idellus C. et V) and grass carp (Ctenopharyngodon idellus) after heating: The relationship between texture and ultrastructure in muscle tissue. Food Res. Int. 2009, 42, 271–278. [Google Scholar] [CrossRef]
- Pearce, K.L.; Rosenvold, K.; Andersen, H.J.; Hopkins, D.L. Water distribution and mobility in meat during the conversion of muscle to meat and ageing and the impacts on fresh meat quality attributes—A review. Meat Sci. 2011, 89, 111–124. [Google Scholar] [CrossRef]
- Aussanasuwannakul, A.; Kenney, P.B.; Brannan, R.G.; Slider, S.D.; Salem, M.; Yao, J. Relating Instrumental Texture, Determined by Variable-Blade and Allo-Kramer Shear Attachments, to Sensory Analysis of Rainbow Trout, Oncorhynchus mykiss, Fillets. J. Food Sci. 2010, 75, S365–S374. [Google Scholar] [CrossRef]
- Messia, M.C.; Di Falco, T.; Panfili, G.; Marconi, E. Rapid determination of collagen in meat-based foods by microwave hydrolysis of proteins and HPAEC–PAD analysis of 4-hydroxyproline. Meat Sci. 2008, 80, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Zarkadas, C.G.; Yu, Z.; Zarkadas, G.C.; Minero-Amador, A. Assessment of the Protein Quality of Beefstock Bone Isolates for Use as an Ingredient in Meat and Poultry Products. J. Agric. Food Chem. 1995, 43, 77–83. [Google Scholar] [CrossRef]
- Suárez, M.; Abad, M.; Ruiz-Cara, T.; Estrada, J.; García-Gallego, M. Changes in muscle collagen content during post mortem storage of farmed sea bream (Sparus aurata): Influence on textural properties. Aquac. Int. 2005, 13, 315–325. [Google Scholar] [CrossRef]
- Zhang, X.; Ookawa, M.; Tan, Y.; Ura, K.; Adachi, S.; Takagi, Y. Biochemical characterisation and assessment of fibril-forming ability of collagens extracted from Bester sturgeon Huso huso × Acipenser ruthenus. Food Chem. 2014, 160, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T. The role of intramuscular connective tissue in meat texture. Anim. Sci. J. 2010, 81, 21–27. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | 0‰ | 3‰ | 6‰ |
|---|---|---|---|
| Asp | 89.90 ± 0.77 a | 82.36 ± 0.02 b | 85.66 ± 2.52 ab |
| Glu | 145.14 ± 0.83 a | 134.28 ± 0.15 b | 137.97 ± 3.94 ab |
| Ser | 31.45 ± 0.71 a | 32.74 ± 0.03 a | 32.35 ± 0.29 a |
| His | 16.16 ± 0.46 a | 17.47 ± 0.15 a | 16.56 ± 0.18 a |
| Gly | 43.37 ± 1.61 a | 45.82 ± 0.44 a | 46.89 ± 1.43 a |
| Thr | 36.12 ± 0.32 a | 36.40 ± 0.00 a | 35.72 ± 0.33 a |
| Arg | 51.13 ± 0.28 a | 51.16 ± 0.12 a | 50.84 ± 0.16 a |
| Ala | 48.38 ± 0.38 a | 48.78 ± 0.13 a | 48.78 ± 0.33 a |
| Tyr | 23.50 ± 0.42 a | 26.98 ± 0.20 b | 24.15 ± 0.98 a |
| Cys-s | 2.43 ± 0.55 a | 2.90 ± 0.14 a | 2.37 ± 0.53 a |
| Val | 44.44 ± 1.00 a | 43.20 ± 0.08 a | 42.92 ± 0.23 a |
| Met | 21.75 ± 0.68 a | 24.76 ± 0.06 a | 22.62 ± 0.96 a |
| Phe | 35.62 ± 0.73 a | 34.57 ± 0.10 a | 34.44 ± 0.13 a |
| Ile | 44.09 ± 1.78 a | 40.33 ± 0.01 a | 40.18 ± 0.18 a |
| Leu | 70.83 ± 1.94 a | 67.56 ± 0.14 a | 66.56 ± 0.39 a |
| Lys | 86.92 ± 2.83 a | 88.27 ± 0.27 a | 84.21 ± 1.82 a |
| Pro | 22.66 ± 3.06 a | 37.32 ± 1.50 b | 35.02 ± 3.50 b |
| Hyp | 26.87 ± 0.55 a | 33.05 ± 0.95 b | 32.96 ± 2.22 b |
| TAA | 840.76 ± 4.45 a | 847.97 ± 1.17 a | 840.20 ± 4.67 a |
| EAA | 355.93 ± 8.12 a | 352.56 ± 0.65 a | 343.21 ± 3.17 a |
| NEAA | 458.90 ± 5.39 a | 432.47 ± 1.80 a | 437.51 ± 3.99 a |
| EAA/TAA | 0.42 ± 0.01 a | 0.42 ± 0.00 a | 0.41 ± 0.00 a |
| EAA/NEAA | 0.78 ± 0.02 a | 0.76 ± 0.00 a | 0.73 ± 0.01 a |
| Amino Acid | 0‰ | 3‰ | 6‰ |
|---|---|---|---|
| Asp | 1.04 ± 0.19 a | 4.49 ± 0.80 b | 3.69 ± 0.34 b |
| Glu | 3.86 ± 0.51 a | 6.29 ± 1.22 a | 5.70 ± 0.42 a |
| Ser | 5.32 ± 0.33 a | 3.01 ± 0.01 b | 3.99 ± 0.06 c |
| His | 24.01 ± 4.14 a | 23.83 ± 5.64 a | 22.74 ± 0.44 a |
| Gly | 62.97 ± 8.42 a | 76.52 ± 8.48 a | 78.61 ± 5.55 a |
| Thr | 13.90 ± 0.14 a | 17.01 ± 0.14 a | 23.63 ± 1.28 b |
| Arg | 1.45 ± 0.01 a | 3.01 ± 0.25 b | 6.63 ± 0.10 c |
| Ala | 10.99 ± 0.16 a | 13.88 ± 1.21 a | 11.48 ± 0.46 a |
| Tyr | 1.35 ± 0.06 a | 2.25 ± 0.12 b | 2.67 ± 0.01 c |
| Cys-s | 0.40 ± 0.06 a | 1.13 ± 0.23 a | 0.65 ± 0.11 a |
| Val | 4.41 ± 0.34 a | 7.03 ± 1.03 b | 9.01 ± 0.03 b |
| Met | 1.66 ± 0.05 a | 2.35 ± 0.11 b | 2.77 ± 0.04 c |
| Phe | 1.53 ± 0.00 a | 2.30 ± 0.23 b | 2.94 ± 0.06 c |
| Ile | 1.68 ± 0.05 a | 1.94 ± 0.15 a | 2.76 ± 0.05 b |
| Leu | 2.90 ± 0.05 a | 1.68 ± 0.28 b | 7.72 ± 0.24 c |
| Lys | 4.43 ± 0.13 a | 5.48 ± 0.32 a | 20.43 ± 0.73 b |
| Pro | 4.28 ± 0.28 a | 7.16 ± 0.65 a | 12.01 ± 0.98 b |
| TFAA | 146.21 ± 12.75 a | 179.70 ± 12.37 ab | 217.44 ± 15.62 b |
| UMAA | 4.90 ± 0.32 a | 11.12 ± 1.73 b | 9.38 ± 1.32 b |
| SWAA | 101.90 ± 9.47 a | 123.06 ± 10.17 a | 150.16 ± 14.11 a |
| BIAA | 38.98 ± 3.57 a | 44.40 ± 3.69 a | 57.25 ± 0.39 b |
| SOAA | 28.91 ± 3.82 a | 34.95 ± 3.91 a | 32.13 ± 2.08 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Shen, Z.; Qi, T.; Xi, R.; Liang, X.; Li, L.; Tang, R.; Li, D. Slight Increases in Salinity Improve Muscle Quality of Grass Carp (Ctenopharyngodon idellus). Fishes 2021, 6, 7. https://doi.org/10.3390/fishes6010007
Zhang X, Shen Z, Qi T, Xi R, Liang X, Li L, Tang R, Li D. Slight Increases in Salinity Improve Muscle Quality of Grass Carp (Ctenopharyngodon idellus). Fishes. 2021; 6(1):7. https://doi.org/10.3390/fishes6010007
Chicago/Turabian StyleZhang, Xi, Zhiyuan Shen, Tianpeng Qi, Rujuan Xi, Xiao Liang, Li Li, Rong Tang, and Dapeng Li. 2021. "Slight Increases in Salinity Improve Muscle Quality of Grass Carp (Ctenopharyngodon idellus)" Fishes 6, no. 1: 7. https://doi.org/10.3390/fishes6010007
APA StyleZhang, X., Shen, Z., Qi, T., Xi, R., Liang, X., Li, L., Tang, R., & Li, D. (2021). Slight Increases in Salinity Improve Muscle Quality of Grass Carp (Ctenopharyngodon idellus). Fishes, 6(1), 7. https://doi.org/10.3390/fishes6010007