
Do Two Different Approaches to the Season in Modeling Affect the Predicted Distribution of Fish? A Case Study for Decapterus maruadsi in the Offshore Waters of Southern Zhejiang, China

Abstract
:1. Introduction
2. Materials and Methods
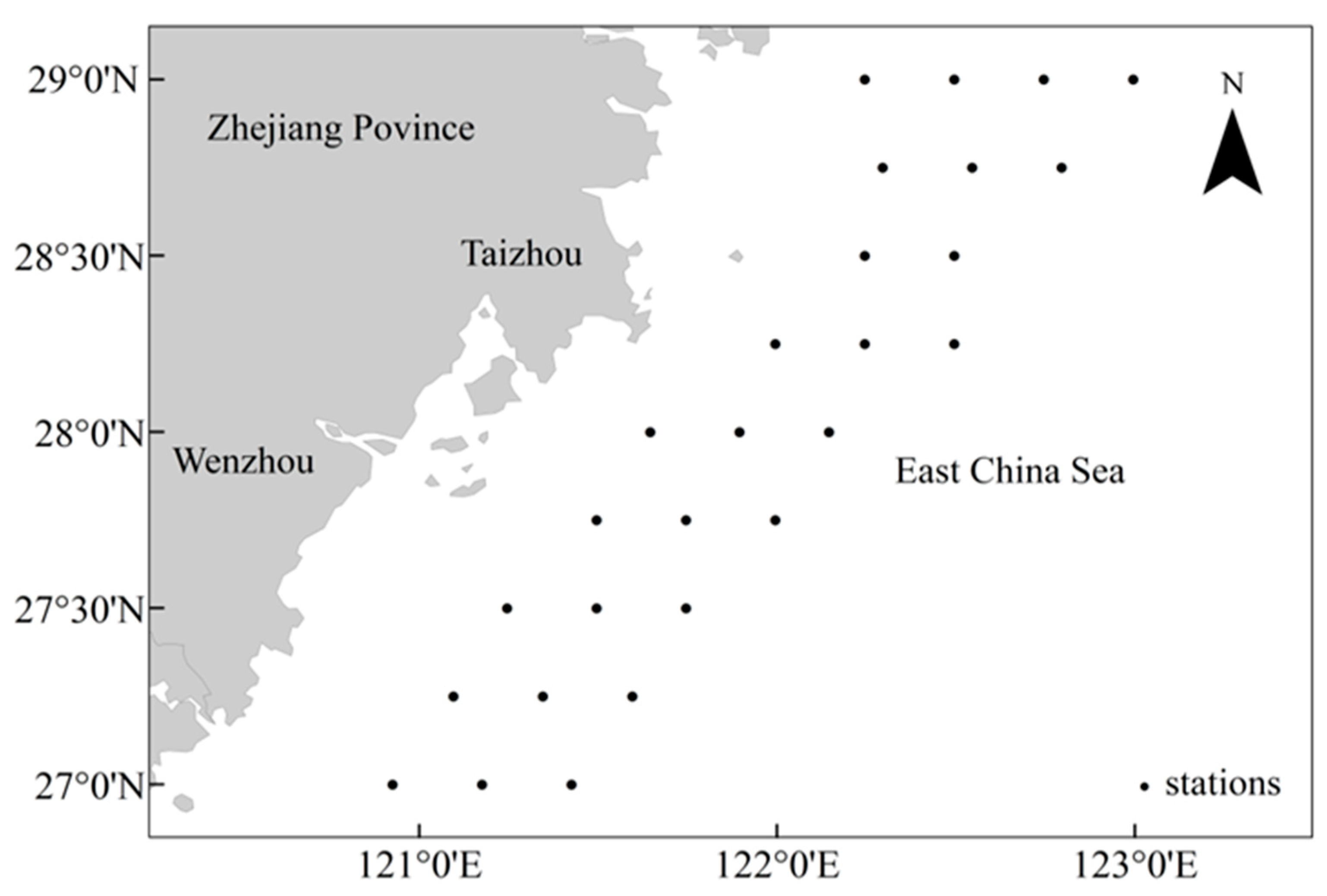
2.1. Data Sources
2.2. Modeling Process
2.2.1. Selection of Explanatory Variables
2.2.2. Tweedie-GAM Development
2.2.3. Model Selection
2.2.4. Model Evaluation
- (1)
- Cross-validation. The predictive performance of the different models was compared by randomly selecting 80% of the data as the training set and the remaining 20% as the test set and running it 1000 times. For each cross-validation, a linear regression model was used to construct a linear relationship between the predicted and observed values, and the root mean square error (RMSE) and mean absolute error (MAE) between the predicted and observed values, as well as the mean of the coefficient of determination (R2), were calculated. When the RMSE and MAE are smaller and the R2 value is closer to 1, the model predicts better [32,33,34]. The regression equation is as follows:where y is the predicted value and Y is the observed value of the model. When a = 0 and b = 1, the predicted value and the observed value (i.e., test data) have a similar spatial pattern.The equation for calculating the MAE is [33]:where n is the number of observations, is the ith observed value, and is the ith predicted value.
- (2)
- To evaluate the fitting effect between different models, this study used two different modeling approaches to predict the density of each station in each year and season and calculated the coefficient of determination (R2) and the significance (P) between the predicted and observed values.
- (3)
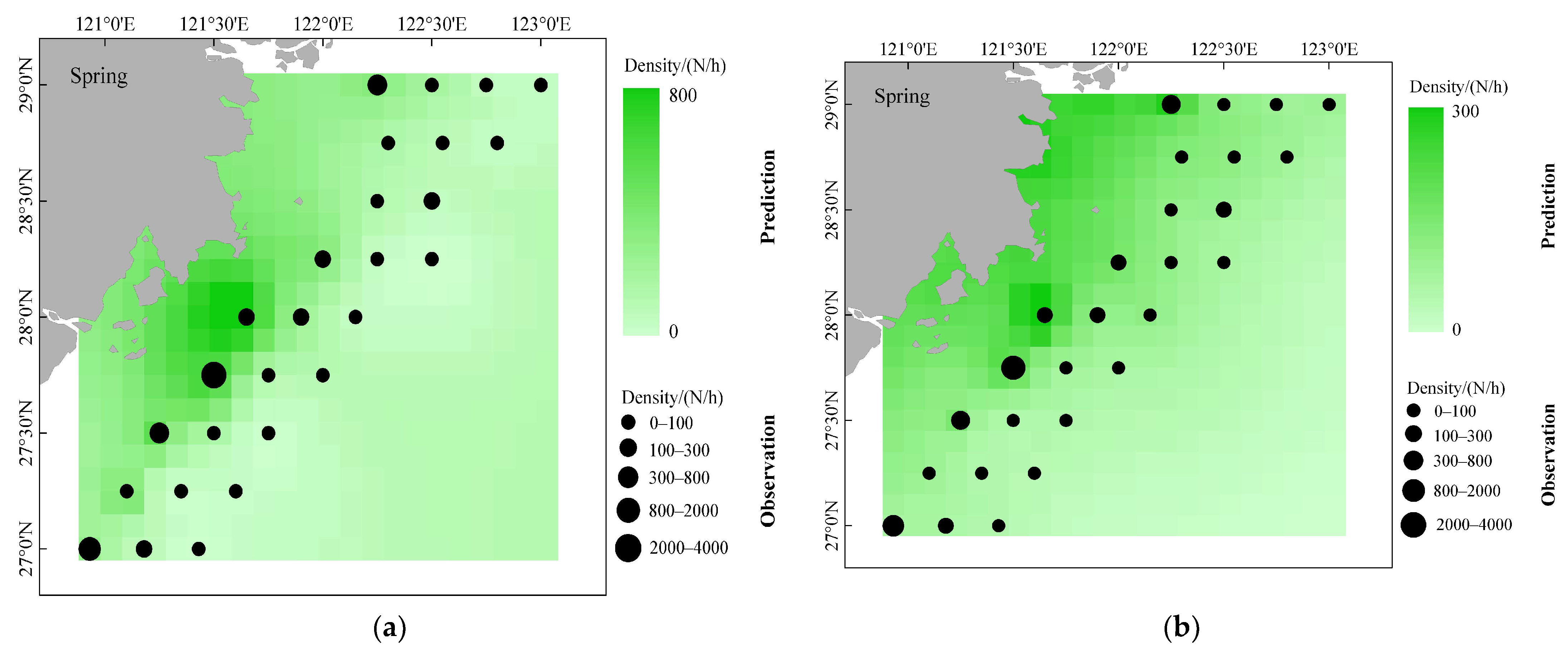
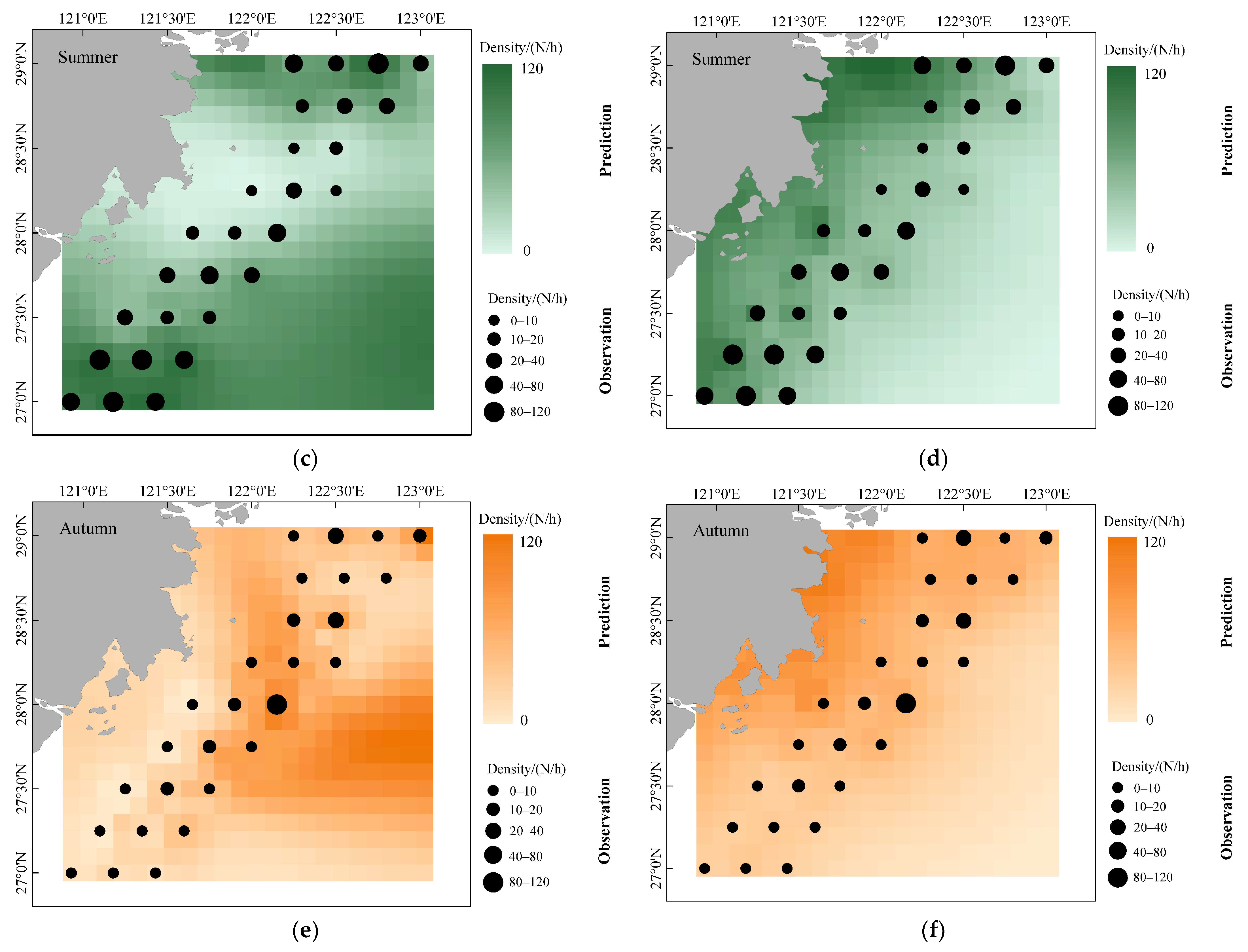
- To assess the differences in the accuracy of predicting the spatial distribution of fishery resources by different modeling approaches, this study predicts the spatial distribution of D. maruadsi in different seasons by using two modeling approaches. A total of 420 quadrilateral grids were created for model prediction in the study area with a spatial resolution of 0.1° × 0.1°. The environmental factors in each grid were interpolated by using inverse distance weighting (IDW). The mean values of the observed and predicted values of D. maruadsi in different seasons from 2015 to 2020 were also superimposed and combined with the living habits of D. maruadsi to evaluate the prediction effects of the two modeling approaches.
3. Results
3.1. Collinearity Test
3.2. Optimal Model
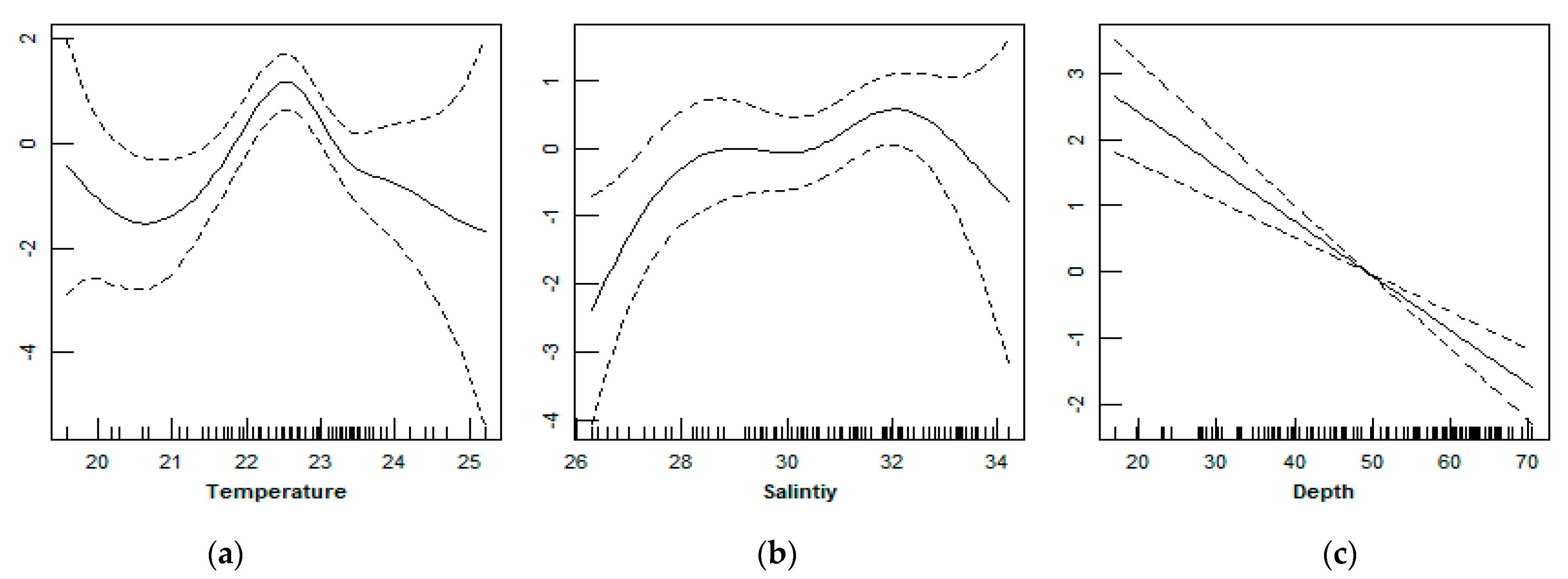
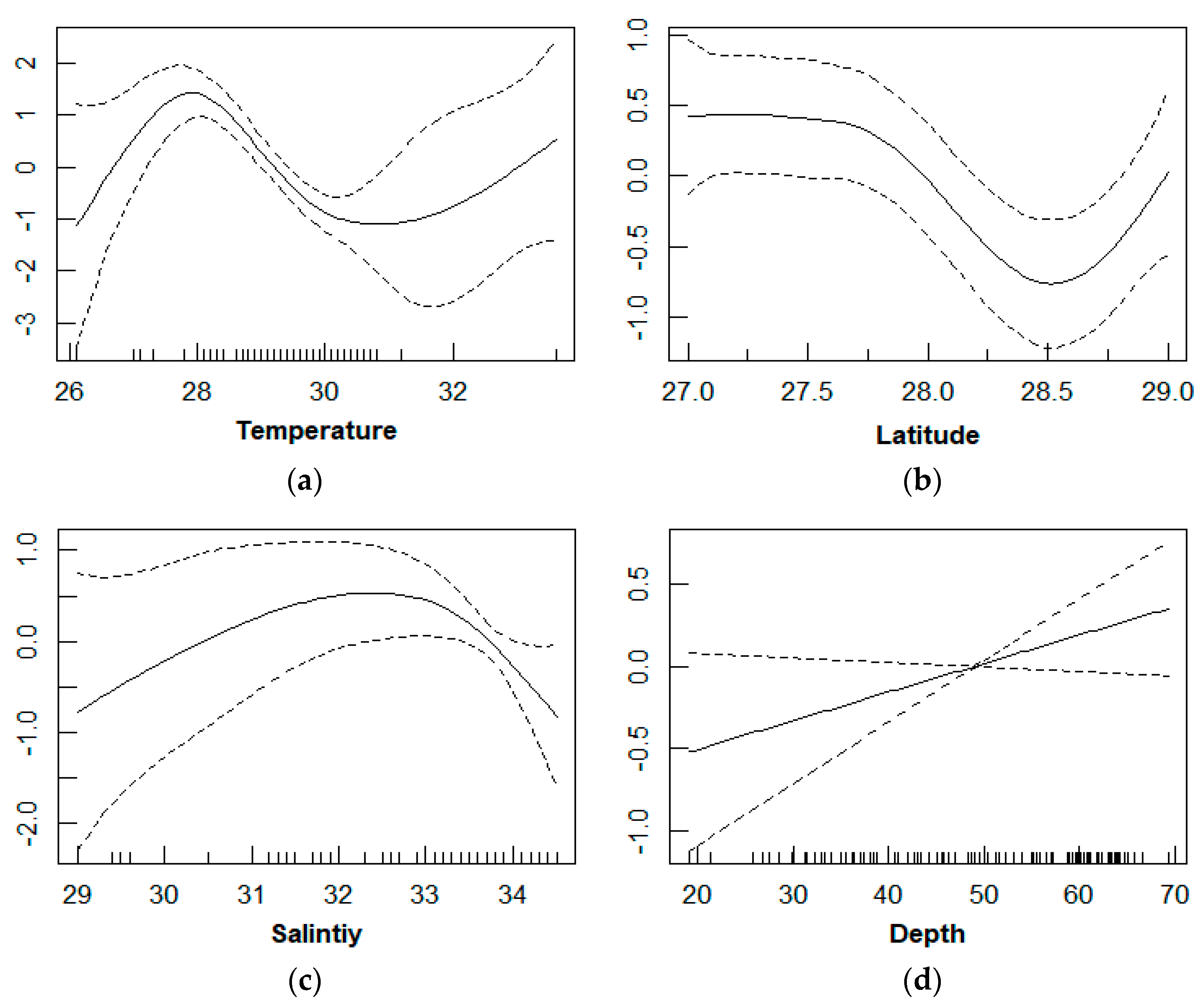
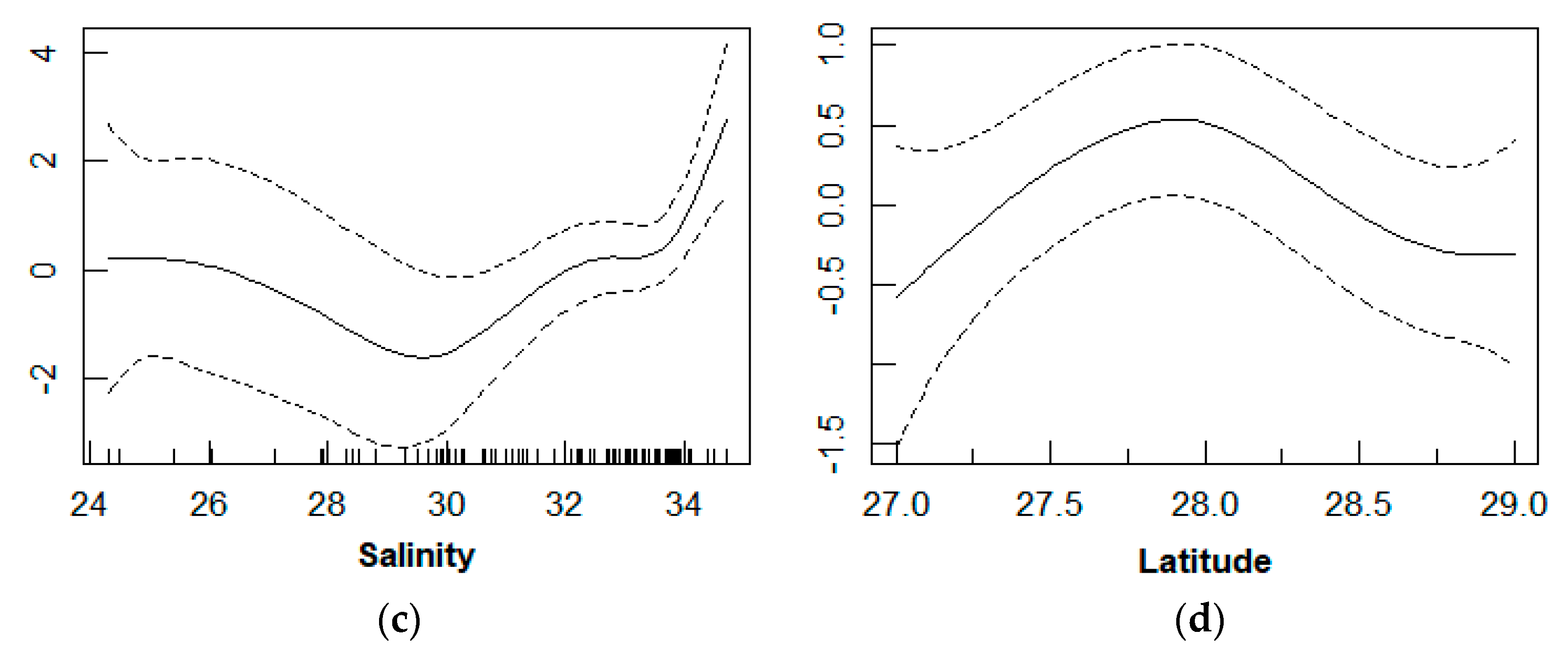
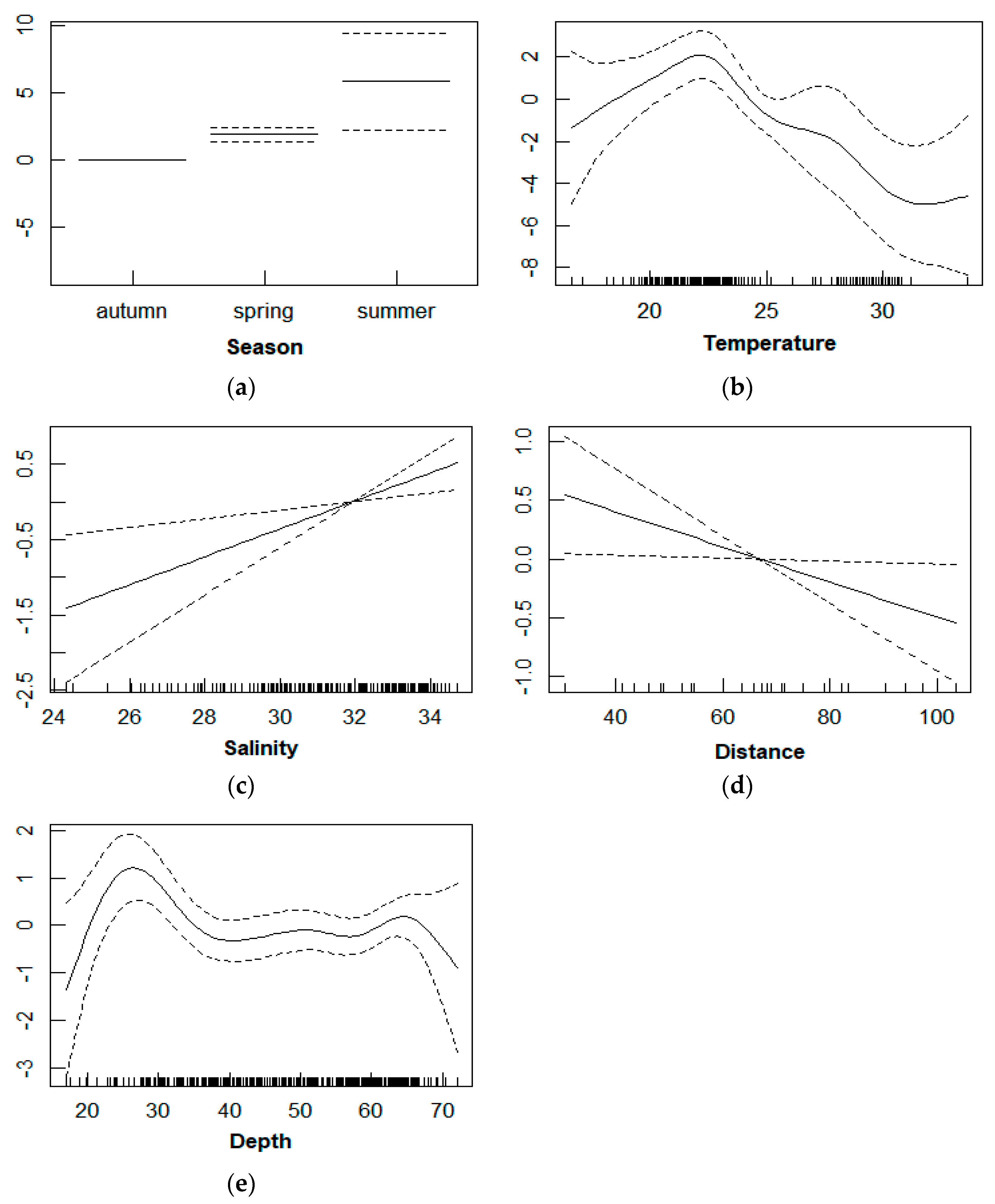
3.3. Relationship between D. maruadsi Density and Environmental Factors
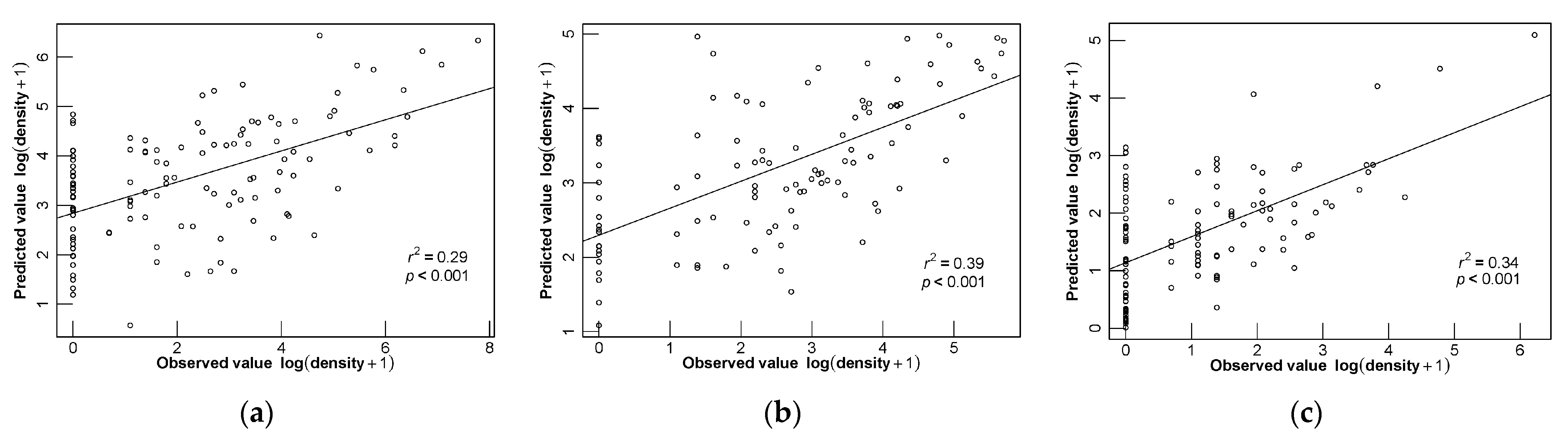
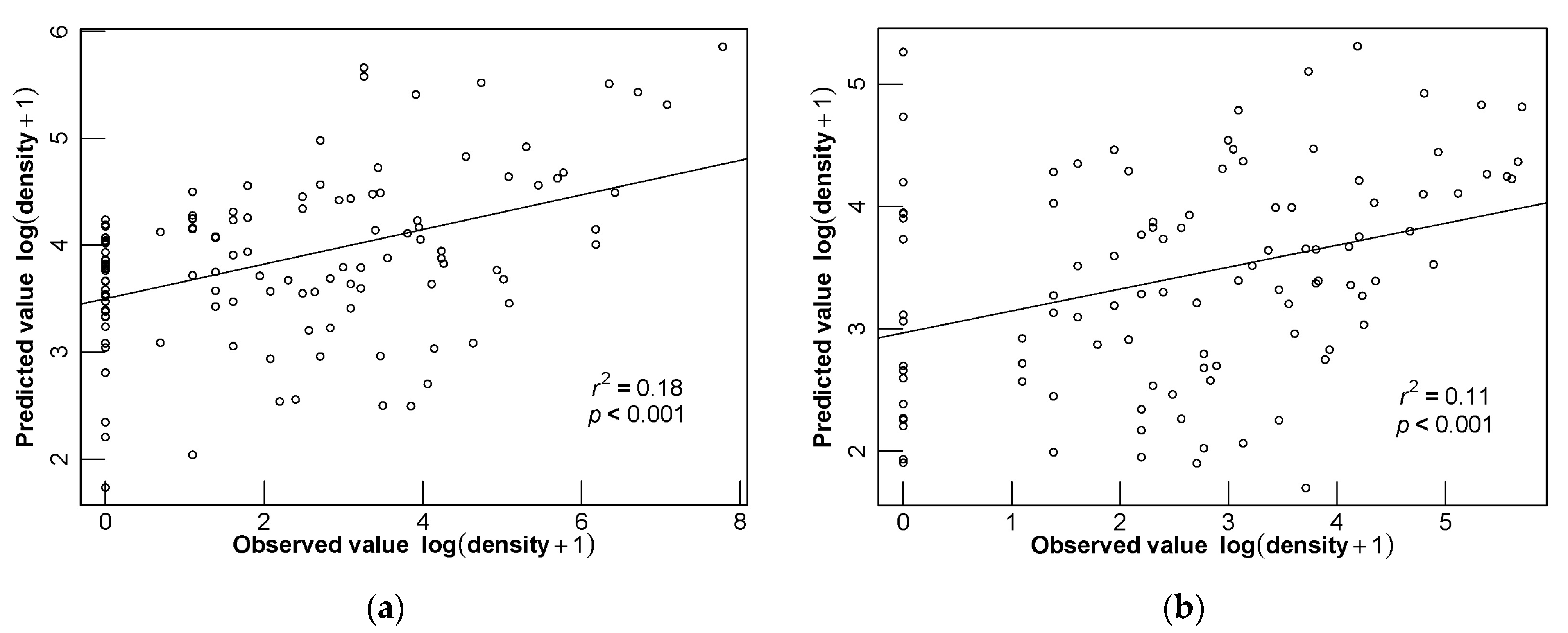
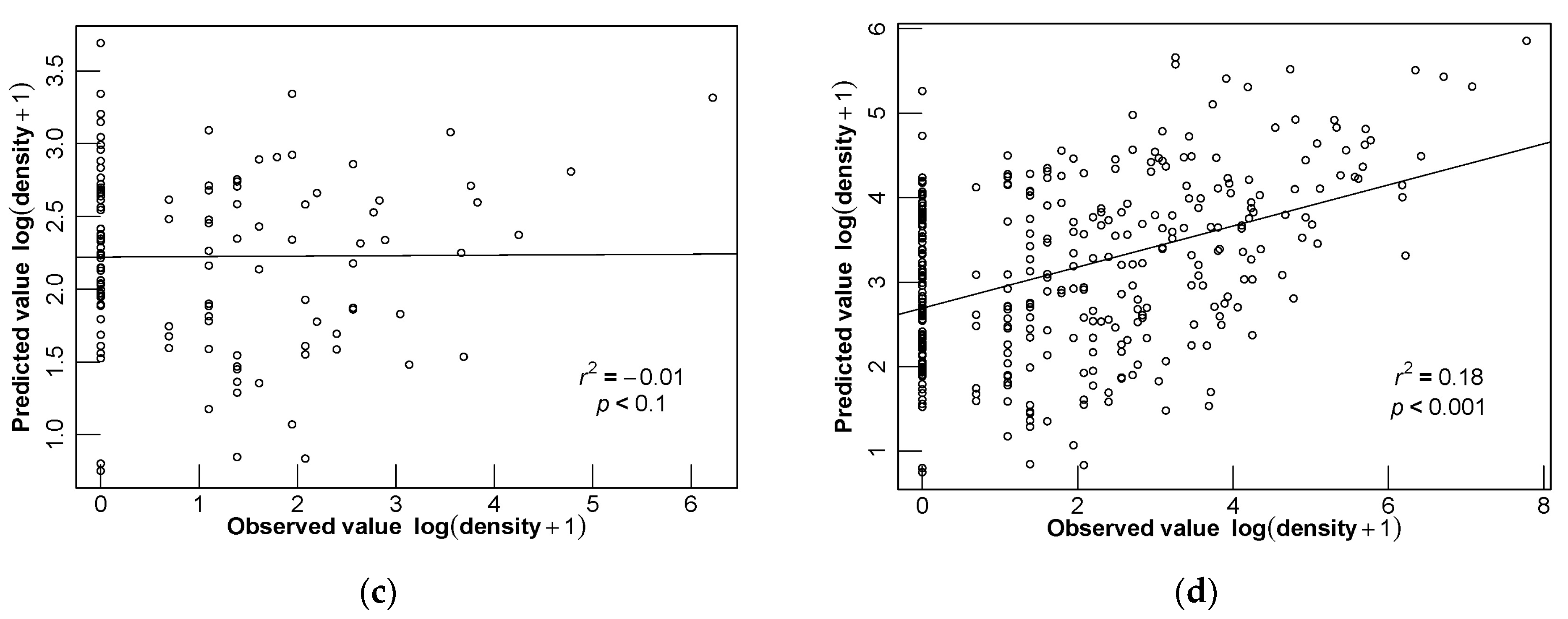
3.4. Model Evaluation
3.5. Different Seasonal Distributions of D. maruadsi and Predictive Performance of Different Models
4. Discussion
4.1. Model Comparison
4.2. Spatial and Temporal Distribution of D. maruadsi Density
4.3. Relationship between D. maruadsi Density and Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, J.; Cao, J.; Tian, S.; Chen, Y.; Zhang, S.; Wang, Z.; Zhou, X. A comparison between two GAM models in quantifying relationships of environmental variables with fish richness and diversity indices. Aquat. Ecol. 2014, 48, 297–312. [Google Scholar] [CrossRef]
- Long, X.Y.; Wan, R.; Li, Z.G.; Ren, Y.P.; Song, P.B.; Tian, Y.J.; Xu, B.D.; Xue, Y. Spatio-temporal distribution of Konosirus punctatus spawning and nursing ground in the South Yellow Sea. Acta Oceanol. Sin. 2021, 40, 133–144. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, Y.T.; Dong, C.M.; Li, Z.L. Spatiotemporal distribution of Decapterus maruadsi in spring and autumn in response to environmental variation in the northern South China Sea. Reg. Stud. Mar. Sci. 2021, 45, 101811. [Google Scholar] [CrossRef]
- Pan, S.Y.; Tian, S.Q.; Wang, X.F.; Dai, L.B.; Gao, C.X.; Tong, J.F. Comparing different spatial interpolation methods to predict the distribution of fishes: A case study of Coilia nasus in the Changjiang River Estuary. Acta Oceanol. Sin. 2021, 40, 119–132. [Google Scholar] [CrossRef]
- Tian, S.Q.; Chen, X.J.; Chen, Y.; Xu, L.X.; Dai, X.J. Evaluating habitat suitability indices derived from CPUE and fishing effort data for Ommatrephes bratramii in the northwestern Pacific Ocean. Fish. Res. 2009, 95, 181–188. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, C.L.; Xu, B.D.; Ji, Y.P.; Ren, Y.P.; Xue, Y. Impacts of trophic interactions on the prediction of spatio-temporal distribution of mid-trophic level fishes. Ecol. Indic. 2022, 138, 108826. [Google Scholar] [CrossRef]
- Yu, H.; Cooper, A.R.; Infante, D.M. Improving species distribution model predictive accuracy using species abundance: Application with boosted regression trees. Ecol. Model. 2020, 432, 109202. [Google Scholar] [CrossRef]
- Liu, X.X.; Wang, J.; Zhang, Y.L.; Yu, H.M.; Xu, B.D.; Zhang, C.L.; Ren, Y.P.; Xue, Y. Comparison between two GAMs in quantifying the spatial distribution of Hexagrammos otakii in Haizhou Bay, China. Fish. Res. 2019, 218, 209–217. [Google Scholar] [CrossRef]
- Liu, X.X.; Gao, C.X.; Zhao, J.; Tian, S.Q.; Ye, S.; Ma, J. Modeling and comparison of count data containing zero values: A case study of Setipinna taty in the south inshore of Zhejiang, China. Environ. Sci. Pollut. Res. 2021, 28, 46827–46837. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Luan, J.; Zhang, C.L.; Ji, Y.P.; Xu, B.D.; Xue, Y.; Ren, Y.P. Matching data types to the objectives of species distribution modeling: An evaluation with marine fish species. Front. Mar. Sci. 2021. [Google Scholar] [CrossRef]
- Meng, W.Z.; Gong, Y.H.; Wang, X.F.; Tong, J.F.; Han, D.Y.; Chen, J.H.; Wu, J.H. Influence of spatial scale selection of environmental factors on the prediction of distribution of Coilia nasus in Changjiang River Estuary. Fishes 2021, 6, 48. [Google Scholar] [CrossRef]
- Bouska, K.L.; Whitledge, G.W.; Lant, C. Development and evaluation of species distribution models for fourteen native central U.S. fish species. Hydrobiologia 2015, 747, 159–176. [Google Scholar] [CrossRef]
- Dai, L.B.; Hodgdon, C.; Tian, S.Q.; Chen, J.H.; Gao, C.X.; Han, D.Y.; Kindong, R.; Ma, Q.Y.; Wang, X.F. Comparative performance of modelling approaches for predicting fish species richness in the Yangtze River Estuary. Reg. Stud. Mar. Sci. 2020, 35, 101161. [Google Scholar] [CrossRef]
- Chang, J.H.; Chen, Y.; Holland, D.; Grabowski, J. Estimating spatial distribution of American lobster Homarus americanus using habitat variables. Mar. Ecol. Prog. Ser. 2010, 420, 145–156. [Google Scholar] [CrossRef]
- Hua, C.X.; Zhu, Q.C.; Shi, Y.C.; Liu, Y. Comparative analysis of CPUE standardization of Chinese Pacific saury (Cololabis saira) fishery based on GLM and GAM. Acta Oceanol. Sin. 2019, 38, 100–110. [Google Scholar] [CrossRef]
- Bellido, J.M.; Pierce, G.J.; Wang, J. Modelling intra-annual variation in abundance of squid Loligo forbesi in Scottish waters using generalised additive models. Fish. Res. 2001, 52, 23–39. [Google Scholar] [CrossRef]
- Shono, H. Application of the Tweedie distribution to zero-catch data in CPUE analysis. Fish. Res. 2008, 93, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.J.; Song, L.M.; Yuan, H.C.; Song, B.; Ebango Ngando, N. A comparative study on habitat models for adult bigeye tuna in the Indian Ocean based on gridded tuna longline fishery data. Fish. Oceanogr. 2021, 30, 584–607. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Cheng, J.H.; Xu, H.X.; Shen, X.Q.; Yu, G.P.; Zheng, Y.J. Fishery Resources and Their Sustainable Utilization in the East China Sea; Fudan University: Shanghai, China, 2007. [Google Scholar]
- Yu, J.; Liu, Z.N.; Chen, P.M.; Yao, L.J. Environmental factors affecting the spatiotemporal distribution of Decapterus maruadsi in the western Guangdong waters, China. Appl. Ecol. Env. Res. 2019, 17, 8485–8499. [Google Scholar] [CrossRef]
- Jiang, R.J.; Xu, H.X.; Jin, H.W.; Zhou, Y.D.; He, Z.T. Feeding habits of blue mackerel scad Decapterus maruadsi Temminck et Schlegel in the East China Sea. J. Fish. China 2012, 36, 216–227. [Google Scholar]
- Xu, W.; Yang, R.; Chen, G.; Gao, C.X.; Ye, S.; Han, D.Y. Feeding ecology of Decapterus maruadsi in the southern coastal area of Zhejiang based on stomach contents and stable isotope analysis. J. Appl. Ecol. 2022, 1–10. [Google Scholar] [CrossRef]
- Cui, M.Y.; Chen, W.F.; Dai, L.B.; Ma, Q.Y. Growth heterogeneity and natural mortality of Japanese scad in offshore waters of southern Zhejiang. J. Fish. Sci. China 2020, 27, 1427–1437. [Google Scholar]
- General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People’s Republic of China. National Standard (Recommended) of the People’s Republic of China: Specifications for Oceanographic Survey—Part 6: Marine Biological Survey, GB/T 12763.6–2007; Standards Press of China: Beijing, China, 2008.
- General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China. GB17378.3–2007 the Specification for Marine Monitoring—Part 3: Sample Collection, Storage and Transportation; China Standards Press: Beijing, China, 2008.
- Tanaka, K.R.; Chang, J.H.; Xue, Y.; Li, Z.G.; Jacobson, L.; Chen, Y. Mesoscale climatic impacts on the distribution of Homarus americanus in the US inshore Gulf of Maine. Can. J. Fish. Aquat. Sci. 2019, 76, 608–625. [Google Scholar] [CrossRef] [Green Version]
- Berg, C.W.; Nielsen, A.; Kristensen, K. Evaluation of alternative age–based methods for estimating relative abundance from survey data in relation to assessment models. Fish. Res. 2014, 151, 91–99. [Google Scholar] [CrossRef]
- Pebesma, E.; Bivand, R.S. S classes and methods for spatial data: The sp package. R News 2005, 5, 9–13. [Google Scholar]
- Bui, D.T.; Lofman, O.; Revhaug, I.; Dick, O. Landslide susceptibility analysis in the Hoa Binh province of Vietnam using statistical index and logistic regression. Nat. Hazards 2011, 59, 1413–1444. [Google Scholar] [CrossRef]
- Planque, B.; Bellier, E.; Lazure, P. Modelling potential spawning habitat of sardine (Sardina pilchardus) and anchovy (Engraulis encrasicolus) in the Bay of Biscay. Fish. Oceanogr. 2007, 16, 16–30. [Google Scholar] [CrossRef]
- Stow, C.A.; Jolliff, J.; McGillicuddy, D.J.; Doney, S.C.; Allen, J.I.; Friedrichs, M.A.M.; Rose, K.A.; Wallhead, P. Skill assessment for coupled biological/physical models of marine systems. J. Mar. Syst. 2009, 76, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Willmott, C.J.; Matsuura, K. Advantages of the mean absolute error (MAE) over the root mean square error (RMSE) in assessing average model performance. Clim. Res. 2005, 30, 79–82. [Google Scholar] [CrossRef]
- Ma, W.; Gao, C.X.; Tang, W.; Qin, S.; Ma, J.; Zhao, J. Relationship between Engraulis japonicus resources and environmental factors based on multi-model comparison in offshore waters of southern Zhejiang, China. J. Mar. Sci. Eng. 2022, 10, 657. [Google Scholar] [CrossRef]
- Wood, S.N.; Pya, N.; Säfken, B. Smoothing parameter and model selection for general smooth models. J. Am. Stat. Assoc. 2016, 111, 1548–1563. [Google Scholar] [CrossRef]
- Franca, S.; Cabral, H.N. Predicting fish species richness in estuaries: Which modelling technique to use? Environ. Model. Softw. 2015, 66, 17–26. [Google Scholar] [CrossRef]
- Zeng, Y.; Yeo, D.C.J. Assessing the aggregated risk of invasive crayfish and climate change to freshwater crabs: A Southeast Asian case study. Biol. Conserv. 2018, 223, 58–67. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Yu, H.M.; Yu, H.Q.; Xu, B.D.; Zhang, C.L.; Ren, Y.P.; Xue, Y.; Xu, L.L. Optimization of environmental variables in habitat suitability modeling for mantis shrimp Oratosquilla oratoria in the Haizhou Bay and adjacent waters. Acta Oceanol. Sin. 2020, 39, 36–47. [Google Scholar] [CrossRef]
- Valavanis, V.D.; Georgakarakos, S.; Kapantagakis, A.; Palialexis, A.; Katara, I. A GIS environmental modelling approach to essential fish habitat designation. Ecol. Model. 2004, 178, 417–427. [Google Scholar] [CrossRef]
- Zhang, C.; Shang, S.; Hong, H. Spatial patterns of annual cycles in surface chlorophyll a in the Taiwan Strait. Acta Oceanol. Sin. 2006, 28, 165–170. [Google Scholar]
- Liu, X.X.; Gao, C.X.; Tian, S.Q.; Qin, S.; Ma, J.; Zhao, J. Distribution of Setipinna taty optimal habitats in the South inshore area of Zhejiang Province based on the habitat suitability index. J. Fish. Sci. China 2020, 27, 1485–1495. [Google Scholar]
- He, L.X.; Fu, D.Y.; Li, Z.L.; Wang, H.; Sun, Y.; Liu, B.; Yu, G. Spatio–temporal distribution of Decapterus maruadsi and its relationship with environmental factors in the northwestern South China Sea. Prog. Fish. Sci. 2022, 1–12. [Google Scholar] [CrossRef]
- Chen, W.F.; Peng, X.; Wang, Z.H.; Xie, Q.L.; Chen, S.B.; Ye, S.; Chen, Y.; Ai, W.M. Community structure characteristics of fishes in the coastal area of south Zhejiang during autumn and winter. Ocean Dev. Manag. 2017, 34, 111–119. [Google Scholar]
- Zheng, Y.J.; Li, J.S.; Zhang, Q.Y.; Hong, W.S. Research progresses of resource biology of important marine pelagic food fishes in China. J. Fish. China 2014, 38, 149–160. [Google Scholar]
- Kindong, R.; Chen, J.H.; Dai, L.B.; Gao, C.X.; Han, D.Y.; Tian, S.Q.; Wu, J.H.; Ma, Q.Y.; Tang, J.Y. The effect of environmental conditions on seasonal and inter–annual abundance of two species in the Yangtze River estuary. Mar. Freshw. Res. 2021, 72, 493–506. [Google Scholar] [CrossRef]
- Lewin, W.C.; Mehner, T.; Ritterbusch, D.; Brämick, U. The influence of anthropogenic shoreline changes on the littoral abundance of fish species in German lowland lakes varying in depth as determined by boosted regression trees. Hydrobiologia 2014, 724, 293–306. [Google Scholar] [CrossRef]
- Fan, J.T.; Huang, Z.R.; Xu, Y.W.; Sun, M.S.; Chen, G.B.; Chen, Z.Z. Habitat model analysis for Decapterus maruadsi in northern south China sea based on remote sensing data. Trans. Oceanol. Limnol. 2018, 3, 142–147. [Google Scholar]
- Niu, S.F.; Wu, R.X.; Zhai, Y.; Zhang, H.R.; Li, Z.L.; Liang, Z.B.; Chen, Y.H. Demographic history and population genetic analysis of Decapterus maruadsi from the northern South China Sea based on mitochondrial control region sequence. PeerJ 2019, 7, e7953. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.L.; Song, H.T.; Bo, Z.L.; Wu, Z.J. A study on mackerel and round scad fishing ground off Zhejiang coast in the summer–autumn season. Mar. Sci. Bull. 1984, 2, 62–70. [Google Scholar]
- Feng, Y.T.; Shi, H.Y.; Hou, G.; Zhao, H.; Dong, C.M. Relationships between environmental variables and spatial and temporal distribution of jack mackerel (Trachurus japonicus) in the Beibu Gulf, South China Sea. PeerJ 2021, 9, e12337. [Google Scholar] [CrossRef]
- Yalcin, E.; Gurbet, R. Environmental Influences on the spatio–temporal distribution of European Hake (Merluccius merluccius) in Izmir Bay, Aegean Sea. Turk. J. Fish. Aquat. Sci. 2016, 16, 1–14. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | VIF | |||||
|---|---|---|---|---|---|---|
| T | S | Depth | Distance | Lon | Lat | |
| Spring | 1.40 | 1.88 | 5.25 | 4.25 | 43.8 | 36.1 |
| 1.38 | 1.85 | 3.78 | 2.78 | - | 1.57 | |
| Summer | 1.06 | 1.54 | 3.91 | 4.74 | 42.83 | 37.8 |
| 1.02 | 1.42 | 3.05 | 2.36 | - | 1.89 | |
| Autumn | 1.71 | 2.35 | 4.70 | 4.28 | 38.68 | 30.91 |
| 1.71 | 2.34 | 4.26 | 1.67 | - | 1.61 | |
| Year | 1.35 | 1.75 | 3.84 | 4.22 | 39.50 | 32.47 |
| 1.35 | 1.75 | 2.78 | 2.28 | - | 1.24 | |
| Model | Optimal Model | Degree of Freedom | p Value | Cumulative Deviance Explained | Deviance Explanation of Each Factor | AIC |
|---|---|---|---|---|---|---|
| Spring-GAM | temperature | 5.129 | <0.001 *** | 18.6% | 18.6% | 1015.15 |
| salinity | 3.745 | 0.04 * | 22.4% | 3.8% | ||
| depth | 1.0001 | <0.001 *** | 42.4% | 20.0% | ||
| Summer-GAM | temperature | 4.524 | <0.001 *** | 35.8% | 35.8% | 878.79 |
| latitude | 3.595 | 0.02 * | 40.8% | 5.0% | ||
| salinity | 2.686 | 0.07 | 44.6% | 3.8% | ||
| depth | 1.001 | 0.09 | 46.5% | 1.9% | ||
| Autumn-GAM | depth | 2.697 | 0.25 | 24.6% | 24.6% | 622.72 |
| temperature | 1.000 | 0.19 | 27.9% | 3.3% | ||
| salinity | 5.052 | 0.01 * | 39.0% | 11.1% | ||
| latitude | 2.684 | 0.19 | 43.7% | 4.7% | ||
| Yearly-GAM | season | - | - | 13.2% | 13.2% | 2601.80 |
| temperature | 6.775 | <0.001 *** | 25.5% | 12.3% | ||
| salinity | 4.308 | <0.01 ** | 27.5% | 2.0% | ||
| depth | 7.975 | <0.01 *** | 33.6% | 6.1% | ||
| distance | 1.000 | 0.029 * | 34.3% | 0.7% |
| Model | MAE | RMSE | R2 |
|---|---|---|---|
| Spring-GAM | 99.10 | 219.6 | 0.31 |
| Summer-GAM | 34.80 | 58.66 | 0.27 |
| Autumn-GAM | 9.14 | 23.03 | 0.47 |
| Yearly-GAM | 42.88 | 147.37 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, W.; Gao, C.; Qin, S.; Ma, J.; Zhao, J. Do Two Different Approaches to the Season in Modeling Affect the Predicted Distribution of Fish? A Case Study for Decapterus maruadsi in the Offshore Waters of Southern Zhejiang, China. Fishes 2022, 7, 153. https://doi.org/10.3390/fishes7040153
Ma W, Gao C, Qin S, Ma J, Zhao J. Do Two Different Approaches to the Season in Modeling Affect the Predicted Distribution of Fish? A Case Study for Decapterus maruadsi in the Offshore Waters of Southern Zhejiang, China. Fishes. 2022; 7(4):153. https://doi.org/10.3390/fishes7040153
Chicago/Turabian StyleMa, Wen, Chunxia Gao, Song Qin, Jin Ma, and Jing Zhao. 2022. "Do Two Different Approaches to the Season in Modeling Affect the Predicted Distribution of Fish? A Case Study for Decapterus maruadsi in the Offshore Waters of Southern Zhejiang, China" Fishes 7, no. 4: 153. https://doi.org/10.3390/fishes7040153
APA StyleMa, W., Gao, C., Qin, S., Ma, J., & Zhao, J. (2022). Do Two Different Approaches to the Season in Modeling Affect the Predicted Distribution of Fish? A Case Study for Decapterus maruadsi in the Offshore Waters of Southern Zhejiang, China. Fishes, 7(4), 153. https://doi.org/10.3390/fishes7040153