
Phylogenetic Relationships of the Pseudogobionini Group (Teleostei: Cyprinidae) with Selection Pressure Analyses to Genes of Mitochondrial Genome

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Samples Collection and DNA Extraction
2.2. PCR Amplification and Sequencing
2.3. Sequence Assembly and Analysis
2.4. Phylogenetic Analysis
2.5. Selection Pressure Analysis
3. Results
3.1. Mitogenome Characteristics and Sequence Variation of the Pseudogobionini Group
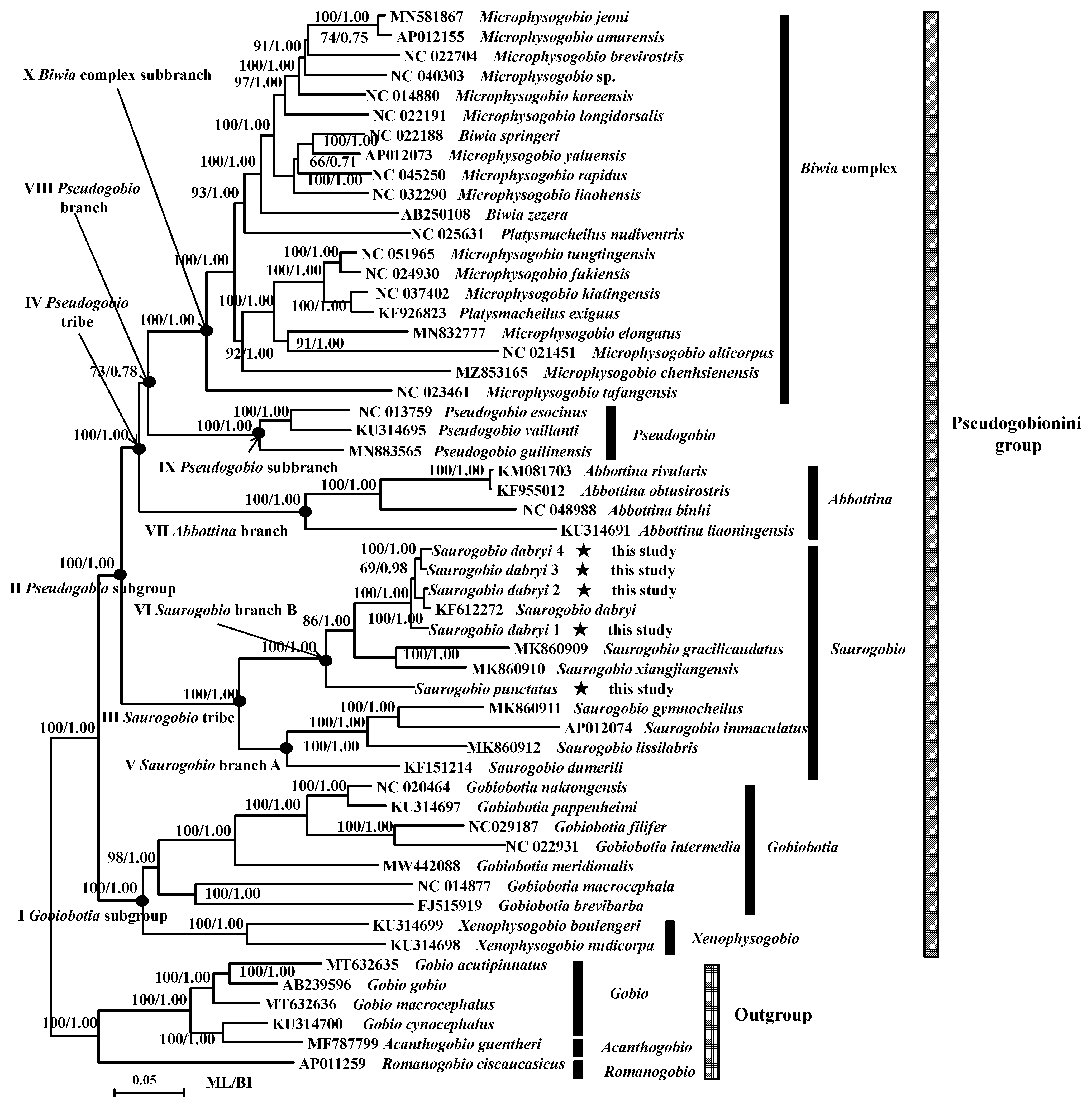
3.2. Phylogenetic Analysis
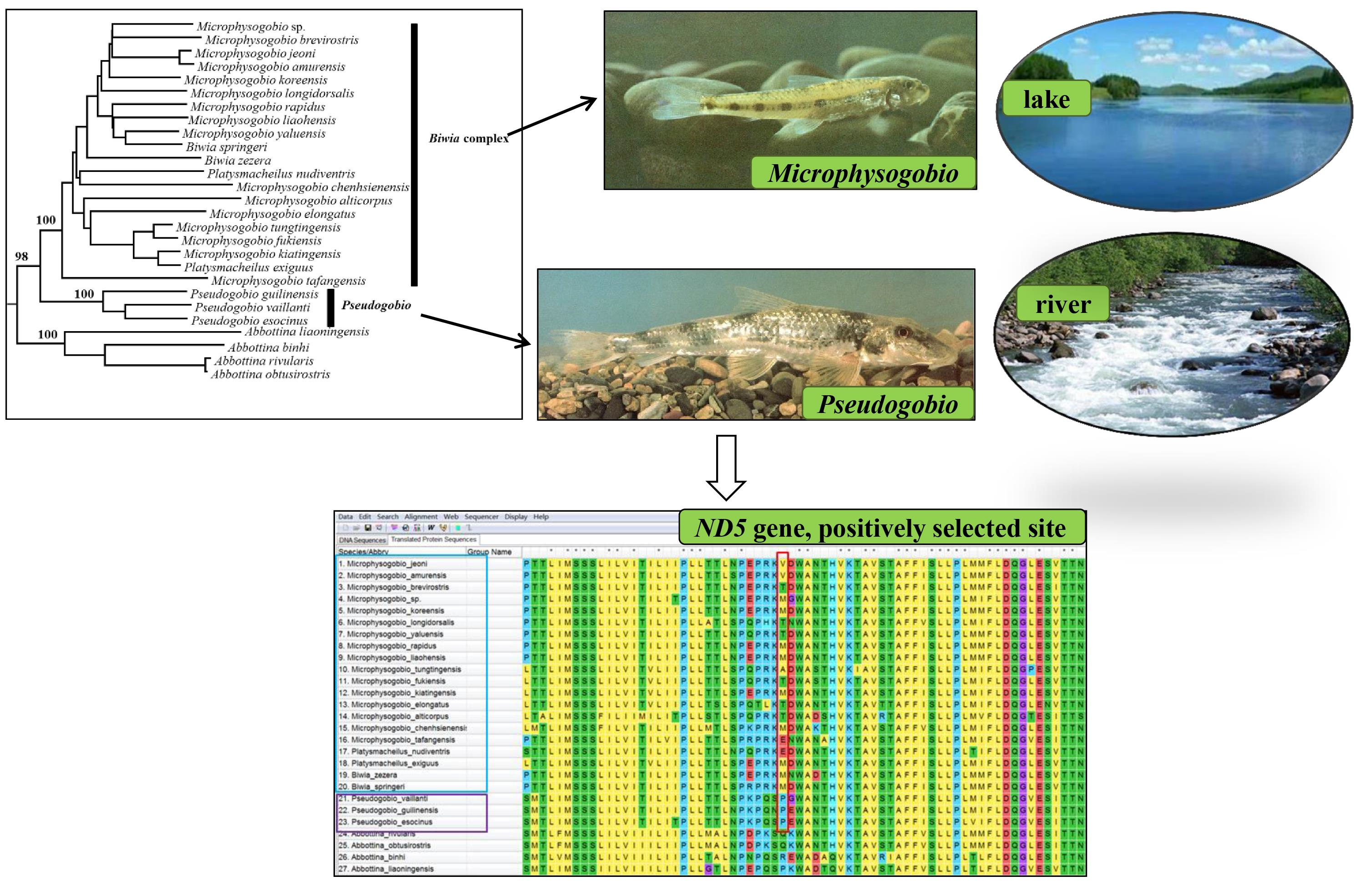
3.3. Selection Pressure Analyses
4. Discussion
4.1. Structural Features of Mitogenomes of the Pseudogobionini Group
4.2. Phylogenetic Relationships of the Pseudogobionini Group
4.3. Classification of the Pseudogobionini Group
- Pseudogobionini group
- ●
- Gobiobotia subgroup
- ●
- Pseudogobio subgroup
- ■
- Saurogobio tribe
- ◆
- Saurogobio branch A
- ◆
- Saurogobio branch B
- ■
- Pseudogobio tribe
- ◆
- Abbottina branch
- ◆
- Pseudogobio branch
- ➢
- Pseudogobio subbranch
- ➢
- Biwia complex subbranch
4.4. Mitochondrial Gene Evolution and Group Differentiation and Adaptations of the Pseudogobionini Fishes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Brown, K. Fish mitochondrial genomics: Sequence, inheritance and functional variation. J. Fish Biol. 2008, 72, 355–374. [Google Scholar] [CrossRef]
- Satoh, T.P.; Miya, M.; Mabuchi, K.; Nishida, M.J. Structure and variation of the mitochondrial genome of fishes. BMC Genom. 2016, 17, 1–20. [Google Scholar] [CrossRef]
- Miya, M.; Nishida, M. The mitogenomic contributions to molecular phylogenetics and evolution of fishes: A 15-year retrospect. Ichthyol Res 2015, 62, 29–71. [Google Scholar] [CrossRef]
- Saitoh, K.; Sado, T.; Mayden, R.; Hanzawa, N.; Nakamura, K.; Nishida, M.; Miya, M. Mitogenomic evolution and interrelationships of the Cypriniformes (Actinopterygii: Ostariophysi): The first evidence toward resolution of higher-level relationships of the world’s largest freshwater fish clade based on 59 whole mitogenome sequences. J. Mol. Evol. 2006, 63, 826–841. [Google Scholar] [CrossRef] [PubMed]
- Tabata, R.; Kakioka, R.; Tominaga, K.; Komiya, T.; Watanabe, K. Phylogeny and historical demography of endemic fishes in Lake Biwa: The ancient lake as a promoter of evolution and diversification of freshwater fishes in western Japan. Ecol. Evol. 2016, 6, 2601–2623. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Zhou, L.; Yang, W.T.; Miao, L.J.; Li, Z.; Zhang, X.J.; Wang, Y.; Gui, J.F. Comparative mitogenome analyses uncover mitogenome features and phylogenetic implications of the subfamily Cobitinae. BMC Genom. 2021, 22, 1–19. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, D.; Zhao, K.; Diogo, R.; Yang, J.; Peng, Z. The origin and divergence of Gobioninae fishes (Teleostei: Cyprinidae) based on complete mitochondrial genome sequences. J. Appl. Ichthyol. 2016, 32, 32–39. [Google Scholar] [CrossRef]
- Da Fonseca, R.R.; Johnson, W.E.; O’Brien, S.J.; Ramos, M.J.; Antunes, A. The adaptive evolution of the mammalian mitochondrial genome. BMC Genom. 2008, 9, 1–22. [Google Scholar] [CrossRef]
- Liu, S.Q.; Mayden, R.L.; Zhang, J.B.; Yu, D.; Tang, Q.Y.; Deng, X.; Liu, H.Z. Phylogenetic relationships of the Cobitoidea (Teleostei: Cypriniformes) inferred from mitochondrial and nuclear genes with analyses of gene evolution. Gene 2012, 508, 60–72. [Google Scholar] [CrossRef]
- Noll, D.; Leon, F.; Brandt, D.; Pistorius, P.; Le Bohec, C.; Bonadonna, F.; Trathan, P.N.; Barbosa, A.; Raya Rey, A.; Dantas, G.P.M.; et al. Positive selection over the mitochondrial genome and its role in the diversification of gentoo penguins in response to adaptation in isolation. Sci. Rep. 2022, 12, 3767. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, S.; Wu, X.; Wei, Q.; Shang, Y.; Sun, G.; Mei, X.; Dong, Y.; Sha, W.; Zhang, H.; et al. High-altitude adaptation in vertebrates as revealed by mitochondrial genome analyses. Ecol. Evol. 2021, 11, 15077–15084. [Google Scholar] [CrossRef]
- Yang, Z.; Wong, W.S.; Nielsen, R. Bayes empirical Bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Yu, L.R.; Yue, P.Q. Origin and evolution of the Pseudogobiini fishes in Eastern Asia. Acta Hydrobiol. Sin. 1996, 20, 1–7. (In Chinese) [Google Scholar]
- Yu, L.R.; Yue, P.Q. Studies on phylogeny of the Pseudogobiini fishes (Cypriniformes: Cyprinidae). Acta Zootaxon. Sin. 1996, 21, 244–255. (In Chinese) [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 17 March 2022).
- Froese, R.; Pauly, D.; FishBase. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 16 March 2022).
- Bănărescu, P. A critical updated checklist of Gobioninae (Pisces, Cyprinidae). Trav. Du Muséum D’histoire Nat. “Grigore Antipa” 1992, 32, 303–330. [Google Scholar]
- Bănărescu, P.; Nalbant, T. Pisces Teleostei Cyprinidae (Gobioninae); Das Tierreich, Lieferung 93; Walter de Guryter: Berlin, Germany, 1973; pp. 1–304. [Google Scholar]
- Yue, P.Q. Gobioninae. In Fauna Sinica, Osteichthyes, Cypriniformes (II); Chen, Y.Y., Ed.; Science Press: Beijing, China, 1998; pp. 232–389. (In Chinese) [Google Scholar]
- Jordan, D.S.; Fowler, H.W. A review of the cyprinoid fishes of Japan. Proc. U.S. Natl. Mus. 1903, 26, 811–862. [Google Scholar] [CrossRef]
- Kawase, S.; Hosoya, K. Biwia yodoensis, a new species from the Lake Biwa/Yodo River Basin, Japan (Teleostei: Cyprinidae). Ichthyol. Explor. Freshw. 2010, 21, 1–7. [Google Scholar]
- Tang, K.L.; Agnew, M.K.; Chen, W.J.; Vincent Hirt, M.; Raley, M.E.; Sado, T.; Schneider, L.M.; Yang, L.; Bart, H.L.; He, S.; et al. Phylogeny of the gudgeons (Teleostei: Cyprinidae: Gobioninae). Mol. Phylogenet. Evol. 2011, 61, 103–124. [Google Scholar] [CrossRef]
- Hosoya, K. Interrelationships of the Gobioninae (Cyprinidae). In Indo-Pacific Fish Biology: Proceedings of the Second International Conference on Indo-Pacific Fishes; Uyeno, T., Arai, R., Taniuchi, T., Matsuura, K., Eds.; Ichthyological Society of Japan: Tokyo, Japan, 1986; pp. 484–501. [Google Scholar]
- Chen, A.H. Studies on Molecular Phylogeny of the Gobioninae (Teleostei: Cyprinidae) (Unpublished Data). Master’s Thesis, Fudan University, Shanghai, China, 2014. [Google Scholar]
- Zhang, R.Y.; Tang, Q.; Deng, L. The complete mitochondrial genome of Microphysogobio elongatus (Teleostei, Cyprinidae) and its phylogenetic implications. ZooKeys 2021, 1061, 57. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Freyhof, J.; Xiong, B.X.; Liu, H.Z. Multiple invasions of Europe by East Asian cobitid loaches (Teleostei: Cobitidae). Hydrobiologia 2008, 605, 17–28. [Google Scholar] [CrossRef]
- Xu, H.; Xue, Y.; Li, X.Y.; Luo, X.; Li, Y.; Li, Y. Complete mitochondrial genome of the Chinese lizard gudgeon, Saurogobio dabryi (Teleostei: Cypriniformes: Cyprinidae). Mitochondrial DNA 2015, 26, 840–841. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Swindell, S.R.; Plasterer, T.N. Seqman. In Sequence Data Analysis Guidebook; Humana Press: New York, NY, USA, 1997; pp. 75–89. [Google Scholar] [CrossRef]
- Perna, N.; Kocher, T. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Watrous, L.E.; Wheeler, Q.D. The out-group comparison method of character analysis. Syst. Biol. 1981, 30, 1–11. [Google Scholar] [CrossRef]
- Yang, J.Q.; He, S.P.; Freyhof, J.; Witte, K.; Liu, H.Z. The phylogenetic relationships of the Gobioninae (Teleostei: Cyprinidae) inferred from mitochondrial cytochrome b gene sequences. Hydrobiologia 2006, 553, 255–266. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol Ecol Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Luo, A.; Qiao, H.; Zhang, Y.; Shi, W.; Ho, S.Y.; Xu, W.; Zhang, A.; Zhu, C. Performance of criteria for selecting evolutionary models in phylogenetics: A comprehensive study based on simulated datasets. BMC Evol. Biol. 2010, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Xu, B.; Yang, Z. PAMLX: A graphical user interface for PAML. Mol. Biol. Evol. 2013, 30, 2723–2724. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H. PAML: A program package for phylogenetic analysis by maximum likelihood. Comput. Appl. Biosci. 1997, 13, 555–556. [Google Scholar] [CrossRef]
- Gao, F.; Chen, C.; Arab, D.A.; Du, Z.; He, Y.; Ho, S.Y. EasyCodeML: A visual tool for analysis of selection using CodeML. Ecol. Evol. 2019, 9, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H. Likelihood ratio tests for detecting positive selection and application to primate lysozyme evolution. Mol. Biol. Evol. 1998, 15, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Swanson, W.J. Codon-substitution models to detect adaptive evolution that account for heterogeneous selective pressures among site classes. Mol. Biol. Evol. 2002, 19, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Sun, Y.B.; Shen, Y.Y.; Irwin, D.M.; Zhang, Y.P. Evaluating the roles of energetic functional constraints on teleost mitochondrial-encoded protein evolution. Mol. Biol. Evol. 2011, 28, 39–44. [Google Scholar] [CrossRef]
- Peng, R.; Zeng, B.; Meng, X.; Yue, B.; Zhang, Z.; Zou, F. The complete mitochondrial genome and phylogenetic analysis of the giant panda (Ailuropoda melanoleuca). Gene 2007, 397, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Peng, Y.; Zhang, S.; Liu, Y.; Zhang, K.; Chen, J.; Zhang, H.; Zhang, C.; Liu, B.J. The complete mitochondrial genome of Parachiloglanis hodgarti and its phylogenetic position within Sisoridae. J. Oceanol. Limnol. 2023, 41, 267–279. [Google Scholar] [CrossRef]
- Wang, Z.L.; Li, C.; Fang, W.Y.; Yu, X.P. The complete mitochondrial genome of two Tetragnatha spiders (Araneae: Tetragnathidae): Severe truncation of tRNAs and novel gene rearrangements in Araneae. Int. J. Biol. Sci. 2016, 12, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Z.; Yang, J.Q.; Tang, Q.Y. Estimated evolutionary tempo of East Asian gobionid fishes (Teleostei: Cyprinidae) from mitochondrial DNA sequence data. Chin. Sci. Bull. 2010, 55, 1501–1510. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Li, X.B.; Yu, D.; Zhu, Y.R.; Ding, B.Q.; Liu, H.Z.; Danley, P.D. Saurogobio punctatus sp. nov., a new cyprinid gudgeon (Teleostei: Cypriniformes) from the Yangtze River, based on both morphological and molecular data. J. Fish Biol. 2018, 92, 347–364. [Google Scholar] [CrossRef]
- Zhou, T.; Shen, X.; Irwin, D.M.; Shen, Y.; Zhang, Y. Mitogenomic analyses propose positive selection in mitochondrial genes for high-altitude adaptation in galliform birds. Mitochondrion 2014, 18, 70–75. [Google Scholar] [CrossRef]
- Nakamura, M. Cyprinid Fishes of Japan—Studies on the life history of cyprinid fishes of Japan, Special Publications of the Research Institute for Natural Resources, No. 4; Research Institute for Natural Resources: Tokyo, Japan, 1969; 1–455. (In Japanese)
- Nakajima, J.; Onikura, N. Spawning behaviour and male mating success of Pike Gudgeon, Pseudogobio esocinus (Cypriniformes, Cyprinidae), in an experimental tank. Ichthyol. Res. 2016, 63, 39–45. [Google Scholar] [CrossRef]
- Tominaga, K.; Kawase, S. Two new species of Pseudogobio pike gudgeon (Cypriniformes: Cyprinidae: Gobioninae) from Japan, and redescription of P. esocinus (Temminck and Schlegel 1846). Ichthyol. Res. 2019, 66, 488–508. [Google Scholar] [CrossRef]
- Vercellino, I.; Sazanov, L.A. The assembly, regulation and function of the mitochondrial respiratory chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Genus | Species | GenBank Accession Number | Length (bp) |
|---|---|---|---|
| Pseudogobio | Pseudogobio esocinus | NC013759 | 16,609 |
| Pseudogobio vaillanti | KU314695 | 16,607 | |
| Pseudogobio guilinensis | MN883565 | 16,609 | |
| Saurogobio | Saurogobio dumerili | KF151214 | 16,604 |
| Saurogobio dabryi | KF612272 | 16,601 | |
| Saurogobio immaculatus | AP012074 | 16,988 | |
| Saurogobio lissilabris | MK860912 | 16,594 | |
| Saurogobio gracilicaudatus | MK860909 | 16,608 | |
| Saurogobio xiangjiangensis | MK860910 | 16,600 | |
| Saurogobio gymnocheilus | MK860911 | 16,604 | |
| Saurogobio dabryi1 * | ON533885 | 16,601 | |
| Saurogobio dabryi2 * | ON533886 | 16,601 | |
| Saurogobio dabryi3 * | ON533887 | 16,600 | |
| Saurogobio dabryi4 * | ON533888 | 16,600 | |
| Saurogobio punctatus * | ON533884 | 16,604 | |
| Abbottina | Abbottina rivularis | KM081703 | 16,597 |
| Abbottina obtusirostris | KF955012 | 16,605 | |
| Abbottina liaoningensis | KU314691 | 16,608 | |
| Abbottina binhi | NC048988 | 16,609 | |
| Biwia | Biwia zezera | AB250108 | 16,599 |
| Biwia springeri | NC022188 | 16,606 | |
| Gobiobotia | Gobiobotia pappenheimi | KU314697 | 16,605 |
| Gobiobotia filifer | NC029187 | 16,613 | |
| Gobiobotia brevibarba | FJ515919 | 16,594 | |
| Gobiobotia macrocephala | NC014877 | 16,610 | |
| Gobiobotia naktongensis | NC020464 | 16,609 | |
| Gobiobotia intermedia | NC022931 | 16,608 | |
| Gobiobotia meridionalis | MW442088 | 16,609 | |
| Microphysogobio | Microphysogobio brevirostris | NC022704 | 16,608 |
| Microphysogobio fukiensis | NC024930 | 16,600 | |
| Microphysogobio tungtingensis | NC051965 | 16,627 | |
| Microphysogobio yaluensis | AP012073 | 16,603 | |
| Microphysogobio kiatingensis | NC037402 | 16,603 | |
| Microphysogobio koreensis | NC014880 | 16,606 | |
| Microphysogobio tafangensis | NC023461 | 16,605 | |
| Microphysogobio longidorsalis | NC022191 | 16,603 | |
| Microphysogobio amurensis | AP012155 | 16,599 | |
| Microphysogobio chenhsienensis | MZ853165 | 16,602 | |
| Microphysogobio alticorpus | NC021451 | 16,568 | |
| Microphysogobio elongatus | MN832777 | 16,612 | |
| Microphysogobio liaohensis | NC032290 | 16,609 | |
| Microphysogobio jeoni | MN581867 | 16,602 | |
| Microphysogobio rapidus | NC045250 | 16,603 | |
| Microphysogobio sp. | NC040303 | 16,607 | |
| Platysmacheilus | Platysmacheilus exiguus | KF926823 | 16,604 |
| Platysmacheilus nudiventris | KM502565 | 16,603 | |
| Xenophysogobio | Xenophysogobio boulengeri | KU314699 | 16,615 |
| Xenophysogobio nudicorpa | KU314698 | 16,617 | |
| Gobio | Gobio gobio | AB239596 | 16,607 |
| Gobio cynocephalus | KU314700 | 16605 | |
| Gobio macrocephalus | MT632636 | 16,609 | |
| Gobio acutipinnatus | MT632635 | 16,609 | |
| Acanthogobio | Acanthogobio guentheri | MF787799 | 16,604 |
| Romanogobio | Romanogobio ciscaucasicus | AP011259 | 16,603 |
| Gobiobotia subgroup four pairs of barbels | Gobiobotia tribe | ||
| urohyal vertical plate is considerably high | |||
| Pseudogobio subgroup one pair of barbels or be absent | Saurogobio tribe urohyal vertical plate is medium high; pre-dorsal length/post-dorsal length < 1; the air-bladder anterior chamber enclosed by the bony capsule | Saurogobio branch A lips smooth or with degenerated papillae | |
| Saurogobio branch B lips with developed papillae | |||
| Pseudogobio tribe urohyal vertical plate is medium high or low; pre-dorsal length/post-dorsal length ≥ 1; the air-bladder anterior chamber enclosed by the membranous capsule | Abbottina branch supraorbital bones absent; curve rob-shape end of 4th vertebral pleural rib | ||
| Pseudogobio branch supraorbital bones present; wing-like lateral expansion end of 4th vertebral pleural rib | Pseudogobio subbranch snout long and prominent; two rows of pharyngeal teeth | ||
| Biwia complex subbranch snout short and blunt; one row of pharyngeal teeth |
| Gene | Parameters | Node I # | Node II | Node III # | Node IV | Node V # | Node VI | Node VII # | Node VIII | Node IX # | Node X |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ND1 | dN/dS | 0.0280 | 0.0376 | 0.0267 | 0.0533 | ||||||
| dN | 0.0079 ** | 0.0019 | 0.0041 | 0.0029 | |||||||
| dS | 0.3053 ** | 0.0889 | 0.2542 | 0.1753 | |||||||
| ND2 | dN/dS | 0.0646 | 0.0687 | 0.0470 | 0.0609 | ||||||
| dN | 0.0136 | 0.0084 | 0.0091 | 0.0070 | |||||||
| dS | 0.2246 | 0.1531 | 0.2775 | 0.1596 | |||||||
| COXI | dN/dS | 0.0082 | 0.1560 | 1.0465 ** | 0.0270 | ||||||
| dN | 0.0006 | 0.0023 | 0.0150** | 0.0003 | |||||||
| dS | 0.1007 | 0.0986 | 0.1537 | 0.0946 | |||||||
| COXII | dN/dS | 0.0184 | 0.0288 | 0.0386 ** | 0.0092 | 0.2157 ** | 0.0061 | ||||
| dN | 0.0022 | 0.0041 | 0.0060 ** | 0.0003 | 0.0341 ** | 0.0006 | |||||
| dS | 0.0957 | 0.0984 | 0.1782 ** | 0.0451 | 0.1987 * | 0.0896 | |||||
| ATP8 | dN/dS | 0.0793 | 0.1924 | 0.0301 | 0.1227 | ||||||
| dN | 0.0101 | 0.0203 | 0.0038 | 0.0082 | |||||||
| dS | 0.1314 | 0.1116 | 0.1421 | 0.0990 | |||||||
| ATP6 | dN/dS | 0.0328 | 0.0405 | 0.0236 | 0.0547 | ||||||
| dN | 0.0082 | 0.0019 | 0.0027 | 0.0031 | |||||||
| dS | 0.2250 ** | 0.0642 | 0.2254 | 0.1489 | |||||||
| COXIII | dN/dS | 0.0213 | 0.0136 | ||||||||
| dN | 0.0042 * | 0.0011 | |||||||||
| dS | 0.1396 | 0.0962 | |||||||||
| ND4 | dN/dS | 0.3164 | 0.0466 | 0.0456 | 0.0385 | ||||||
| dN | 0.0071 | 0.0058 | 0.0072 | 0.0052 | |||||||
| dS | 0.2256 * | 0.1489 | 0.1557 | 0.1534 | |||||||
| ND5 | dN/dS | 0.0544 | 0.0459 | ||||||||
| dN | 0.0109 * | 0.0036 | |||||||||
| dS | 0.2182 ** | 0.0730 | |||||||||
| ND6 | dN/dS | 0.0411 | 0.0531 | ||||||||
| dN | 0.0090 | 0.0077 | |||||||||
| dS | 0.2511 | 0.1695 | |||||||||
| Cyt b | dN/dS | 0.0281 | 0.0148 | 0.0379 ** | 0.0076 | ||||||
| dN | 0.0063 ** | 0.0008 | 0.0103 ** | 0.0014 | |||||||
| dS | 0.2722 * | 0.1066 | 0.2408 | 0.1608 |
| Model | Ln L | Estimates of Parameters | Model Compared | LRT p-Value | Positively Selected Sites | ||
|---|---|---|---|---|---|---|---|
| M3 | −5078.253651 | p0 = 0.84992 | p1 = 0.14517 | p2 = 0.00491 | M0 vs. M3 | 0.00000 | [-] |
| ω0 = 0.0062 | ω1 = 0.19028 | ω2 = 1.62404 | |||||
| M0 | −5130.798044 | ω0 = 0.03624 | Not Allowed | ||||
| M2a | −5096.161404 | p0 = 0.96931 | p1 = 0.03069 | p2 = 0.00000 | M1a vs. M2a | 1.00000 | [-] |
| ω0 = 0.02251 | ω1 = 1.00000 | ω2 = 36.02152 | |||||
| M1a | −5096.161404 | p0 = 0.96931 | p1 = 0.03069 | Not Allowed | |||
| ω0 = 0.02251 | ω1 = 1.00000 | ||||||
| M8 | −5081.175068 | p0 = 0.98839 | p1 = 0.04111 | q = 0.36593 | M7 vs. M8 | 0.00013 | 186 F 0.706, 193 S 0.952 * |
| (p1 = 0.01161) | ω = 1.00000 | ||||||
| M7 | −5090.122897 | p = 0.04073 | q = 0.32459 | Not Allowed | |||
| M8a | −5081.175065 | p0 = 0.98839 | p = 0.04101 | q = 0.36457 | M8a vs. M8 | 0.99804 | Not Allowed |
| (p1 = 0.01161) | ω = 1.00000 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Zhang, X.; Liu, H. Phylogenetic Relationships of the Pseudogobionini Group (Teleostei: Cyprinidae) with Selection Pressure Analyses to Genes of Mitochondrial Genome. Fishes 2023, 8, 201. https://doi.org/10.3390/fishes8040201
Chen L, Zhang X, Liu H. Phylogenetic Relationships of the Pseudogobionini Group (Teleostei: Cyprinidae) with Selection Pressure Analyses to Genes of Mitochondrial Genome. Fishes. 2023; 8(4):201. https://doi.org/10.3390/fishes8040201
Chicago/Turabian StyleChen, Lin, Xiaoyu Zhang, and Huanzhang Liu. 2023. "Phylogenetic Relationships of the Pseudogobionini Group (Teleostei: Cyprinidae) with Selection Pressure Analyses to Genes of Mitochondrial Genome" Fishes 8, no. 4: 201. https://doi.org/10.3390/fishes8040201
APA StyleChen, L., Zhang, X., & Liu, H. (2023). Phylogenetic Relationships of the Pseudogobionini Group (Teleostei: Cyprinidae) with Selection Pressure Analyses to Genes of Mitochondrial Genome. Fishes, 8(4), 201. https://doi.org/10.3390/fishes8040201