
Temporal Variation in Von Bertalanffy Growth Curves and Generation Time of Southern Gulf of St. Lawrence Spring and Fall Spawning Atlantic Herring (Clupea harengus)


Abstract
:1. Introduction
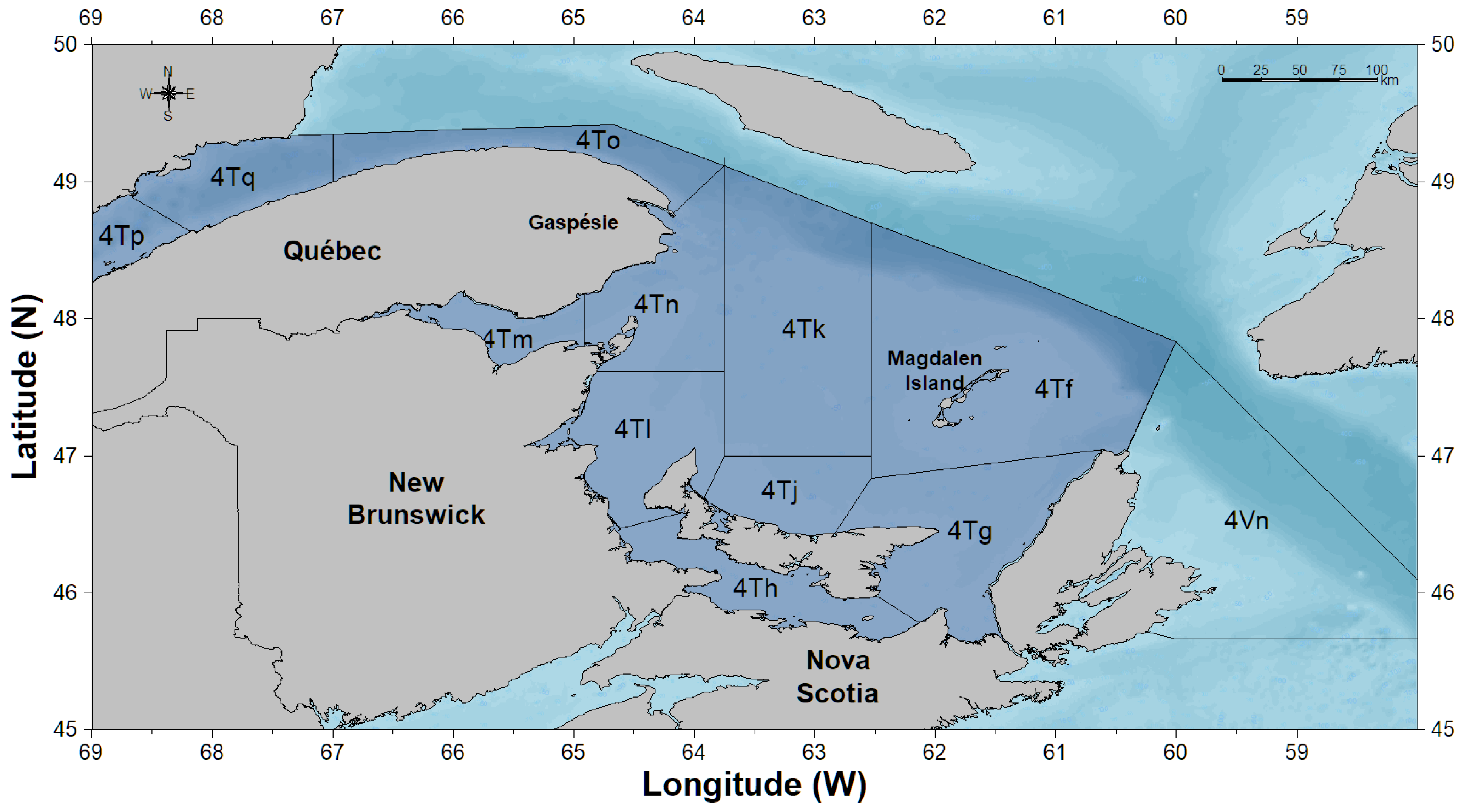
2. Materials and Methods
3. Results
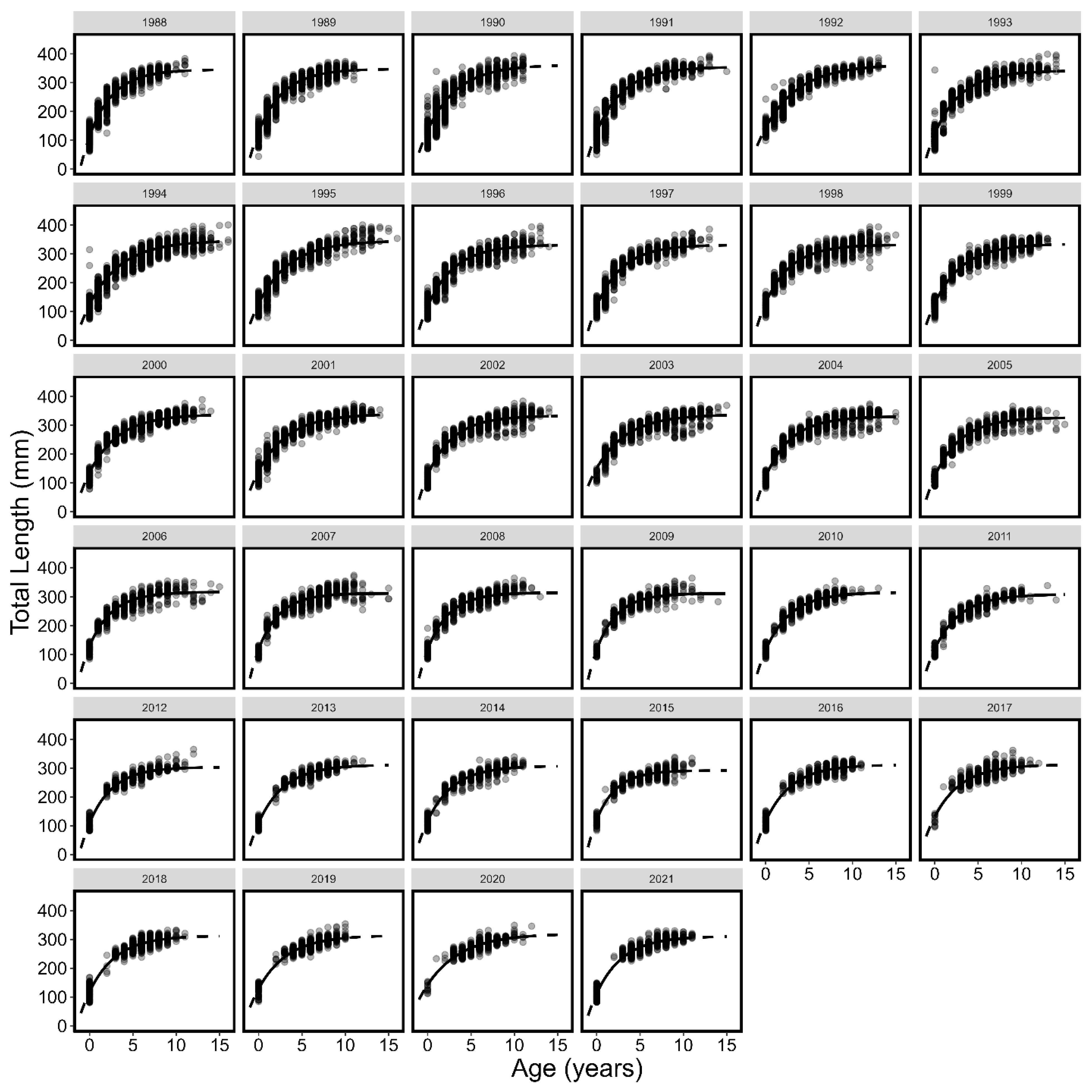
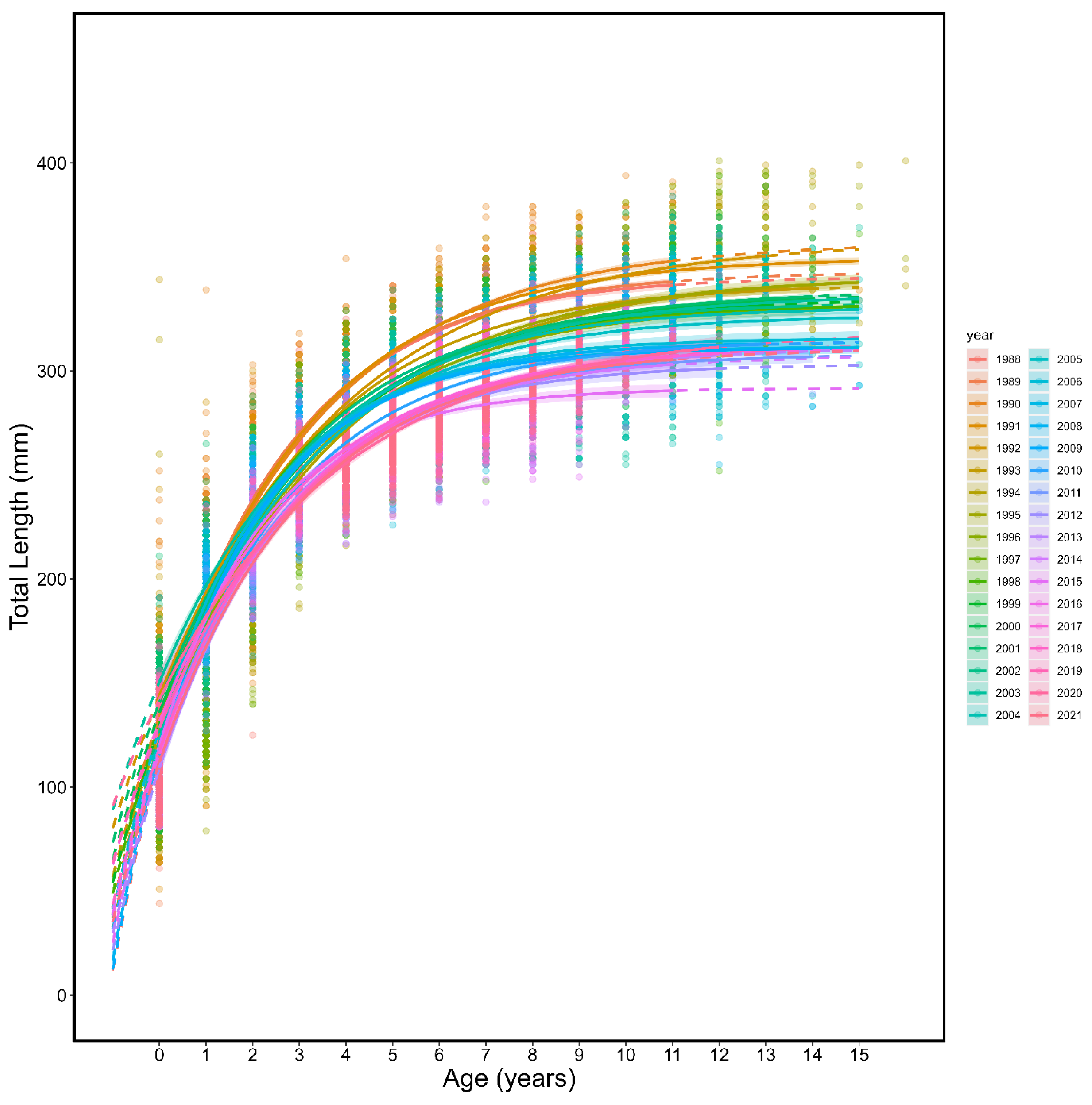
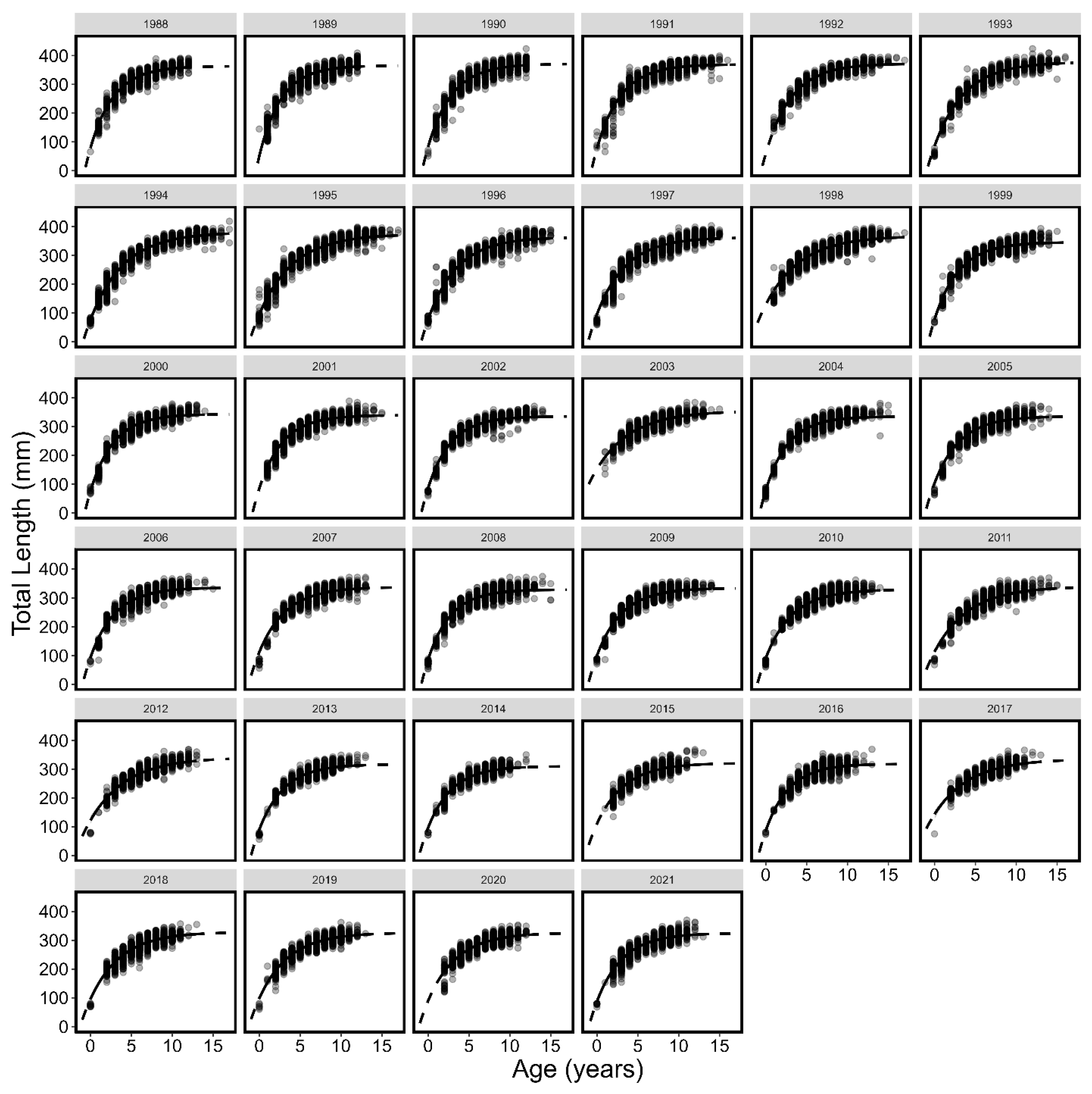
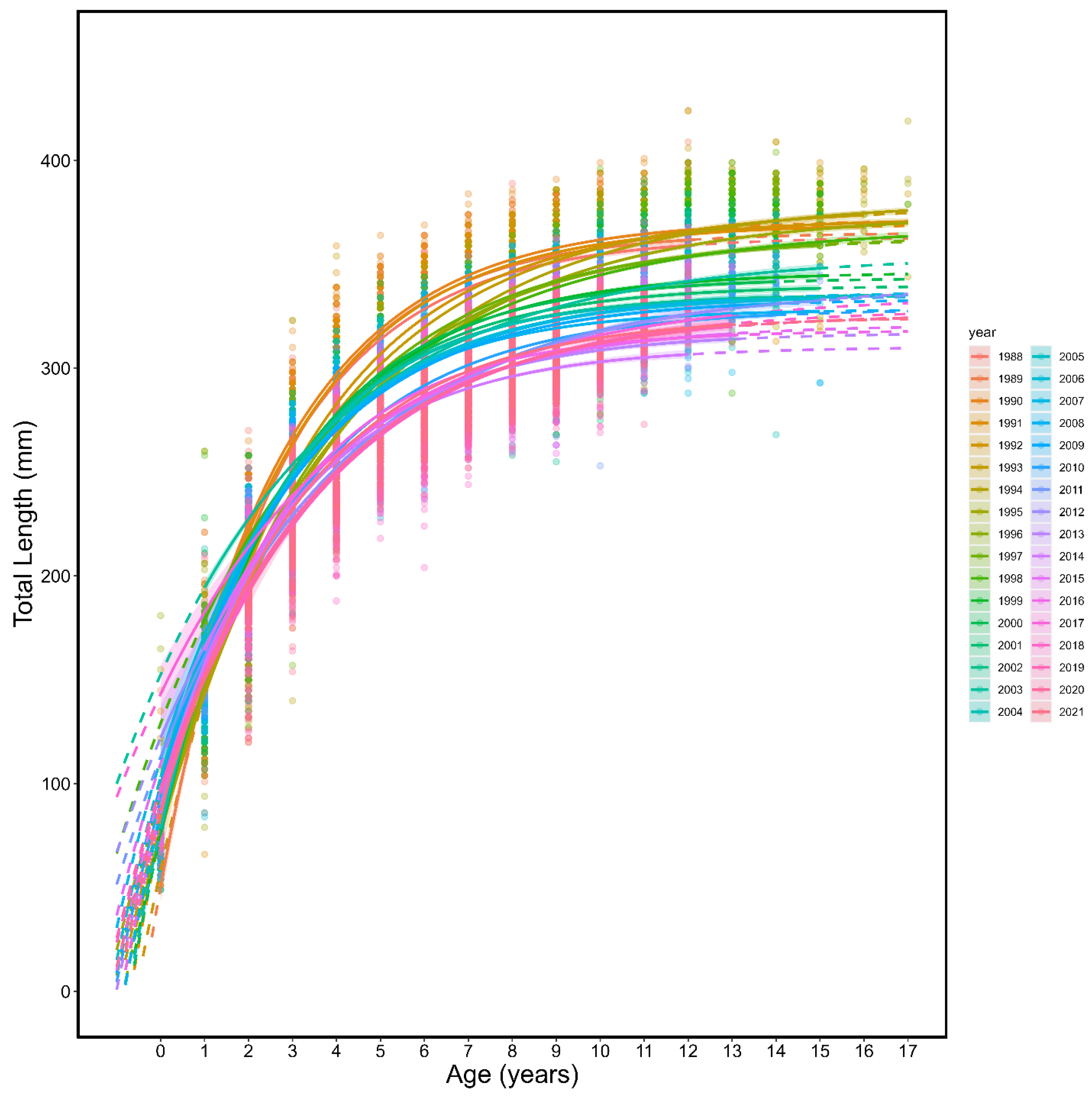
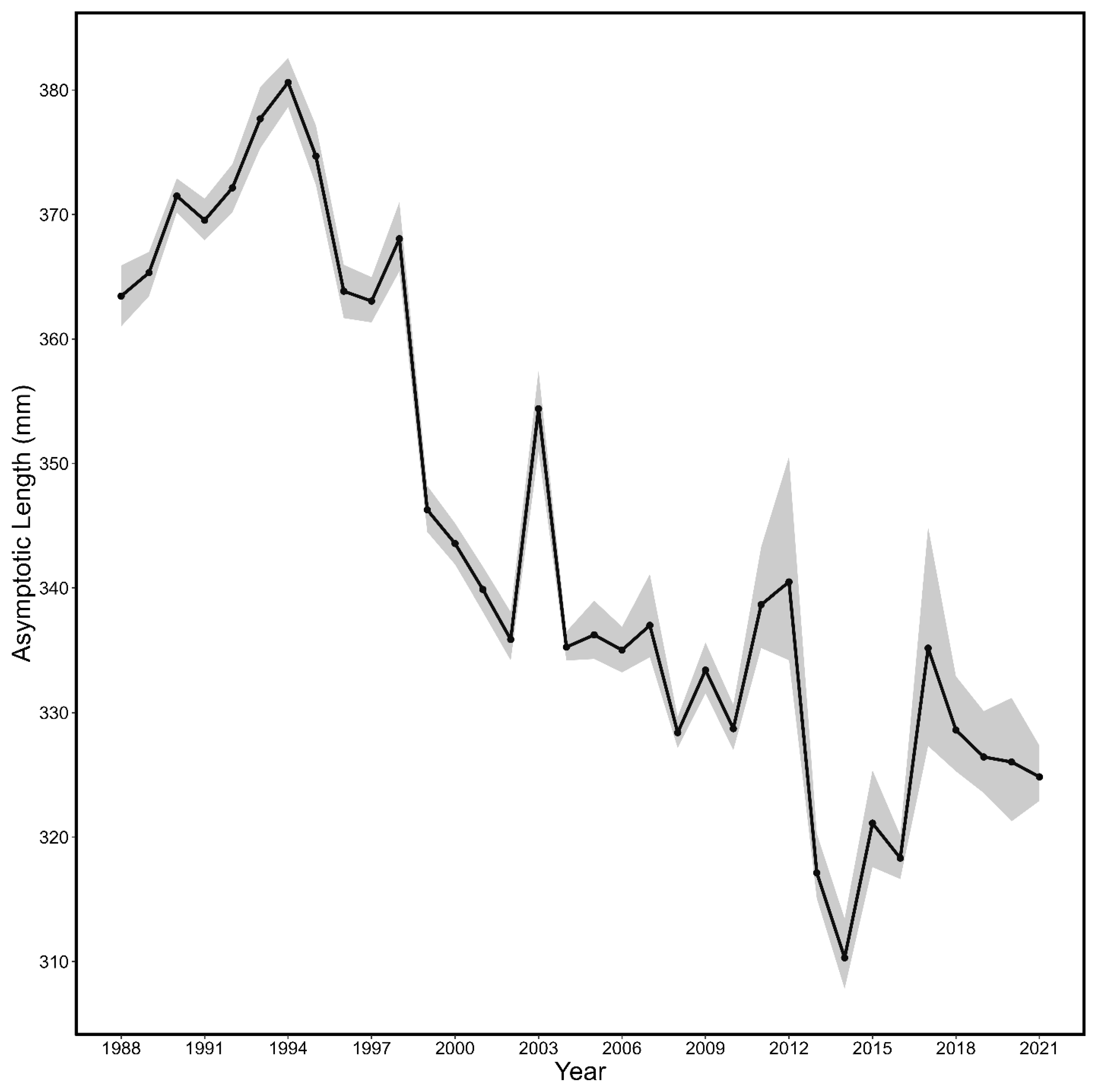
3.1. Von Bertalanffy Growth Curves
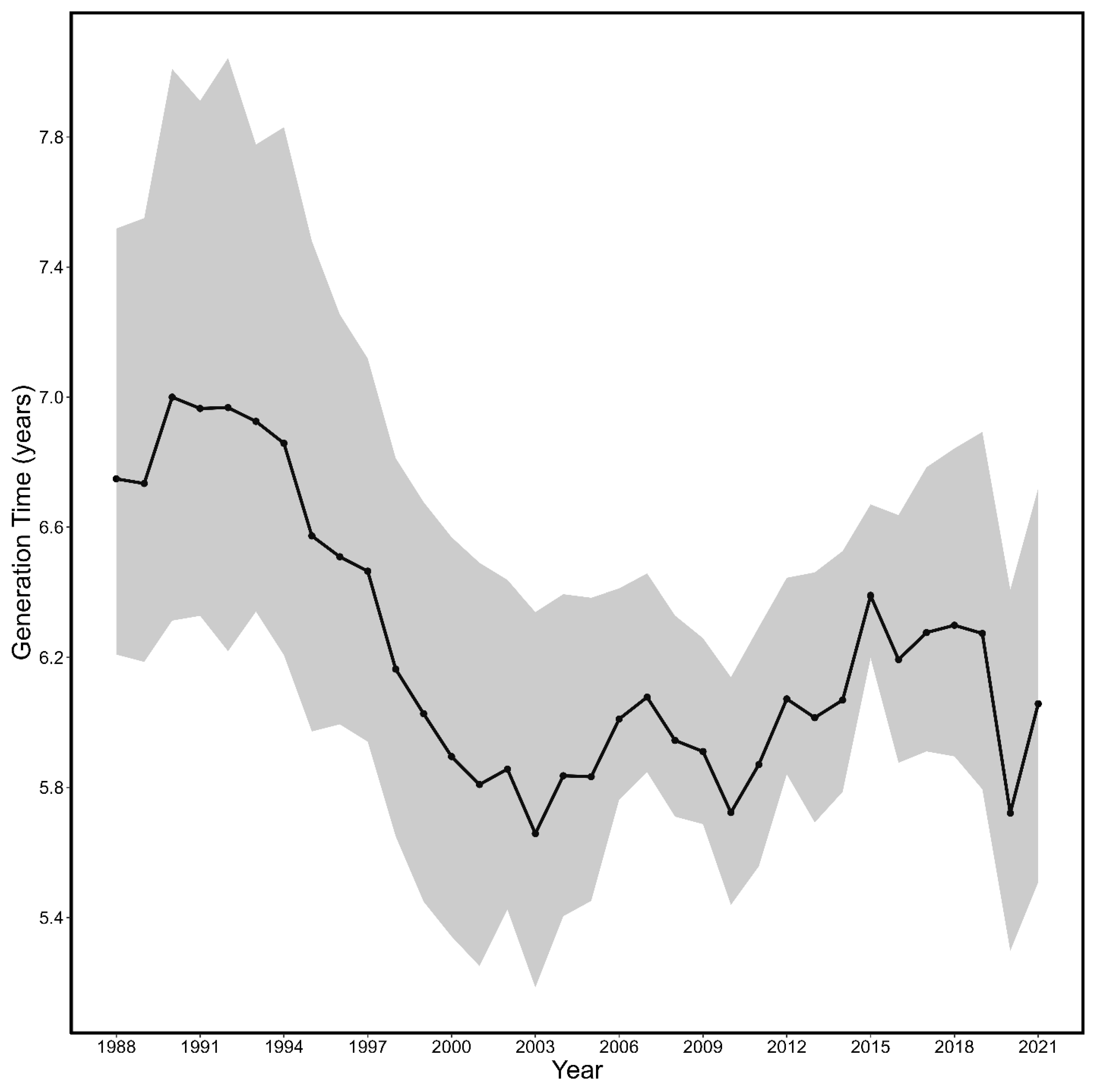
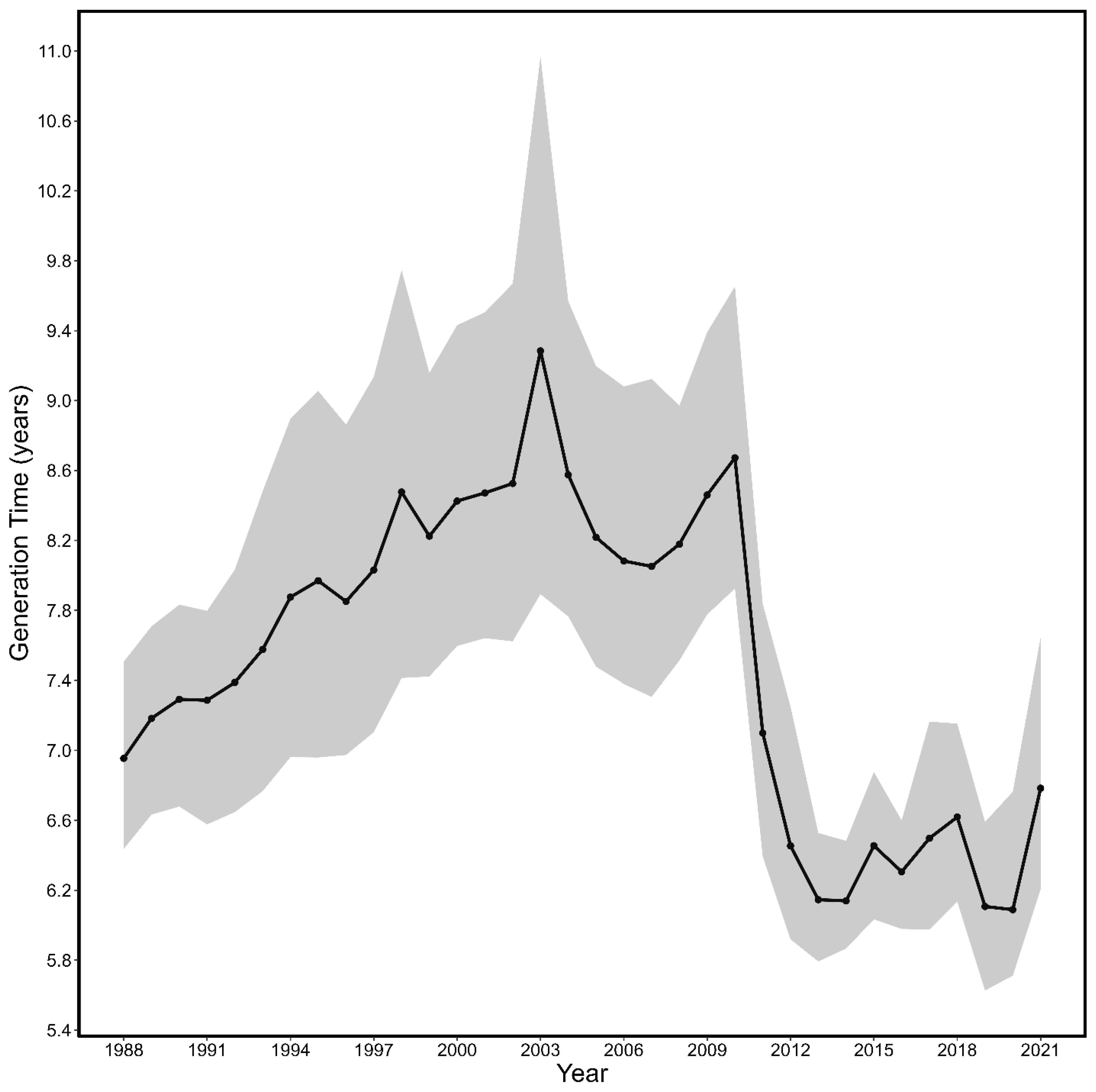
3.2. Generation Time
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rolland, N.; Turcotte, F.; Mcdermid, J.L.; Dejong, R.A.; Landry, L. Assessment of the NAFO Division 4TVn Southern Gulf of St. Lawrence Atlantic Herring (Clupea Harengus) in 2020-2021. DFO Can. Sci. Advis. Sec. Res. Doc. 2022, 2022/068, xii + 142 p. [Google Scholar]
- Claytor, R.R. Fishery Acoustic Indices for Assessing Atlantic Herring Populations. Can. Tech. Rep. Fish. Aquat. Sci. 2001, 2359, 213 p. [Google Scholar]
- Lamichhaney, S.; Fuentes-Pardo, A.P.; Rafati, N.; Ryman, N.; McCracken, G.R.; Bourne, C.; Singh, R.; Ruzzante, D.E.; Andersson, L. Parallel Adaptive Evolution of Geographically Distant Herring Populations on Both Sides of the North Atlantic Ocean. Proc. Natl. Acad. Sci. USA 2017, 114, E3452–E3461. [Google Scholar] [CrossRef] [PubMed]
- Messieh, S.N. Some Characteristics of Atlantic Herring (Clupea harengus) Spawning in the Southern Gulf of St. Lawrence. NAFO Sci. Counc. Stud. 1987, 11, 53–61. [Google Scholar]
- Simon, J.; Stobo, W.T. The 1982-1983 4Vn Herring Biological Update. DFO CAFSAC Res. Doc. 1983, 83/49, 28 p. [Google Scholar]
- Turcotte, F.; Swain, D.P.; McDermid, J.L.; Dejong, R.A. Assessment of the NAFO Division 4TVn Southern Gulf of St. Lawrence Atlantic Herring (Clupea Harengus) in 2018-2019. DFO Can. Sci. Advis. Sec. Res. Doc. 2021, 2021/030, xiv + 158 p. [Google Scholar]
- Jennings, S.; Reynolds, J.D.; Mills, S.C. Life History Correlates of Responses to Fisheries Exploitation. Proc. R. Soc. B Biol. Sci. 1998, 265, 333–339. [Google Scholar] [CrossRef]
- Winemiller, K.O. Life History Strategies, Population Regulation, and Implications for Fisheries Management. Can. J. Fish. Aquat. Sci. 2005, 62, 872–885. [Google Scholar] [CrossRef]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations; Chapman and Hall: London, UK, 1957. [Google Scholar]
- Froese, R.; Palomares, M.L.; Pauly, D. Estimation of Life-History Key Facts. Available online: https://www.fishbase.se/manual/key%20facts.htm (accessed on 3 January 2023).
- Beaudry-Sylvestre, M.; Benoit, H.P.; Hutchings, J.A. Combined Effects of Exploitation and Environmental Change on Life History: A Comparative Analysis on Atlantic Herring. ICES J. Mar. Sci. 2022, 79, 2728–2740. [Google Scholar] [CrossRef]
- Ogle, D.H.; Brenden, T.O.; McCormick, J.L. Growth Estimation: Growth Models and Statistical Inference. In Age and Growth of Fishes: Principles and Techniques; Quist, M.C., Isermann, D.A., Eds.; American Fisheries Society: Bathesda, MD, USA, 2017; pp. 265–359. [Google Scholar]
- Ogle, D.H. Introductory Fisheries Analyses with R; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- R Development Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-Project.Org/ (accessed on 3 January 2023).
- Beverton, R.J.H. Patterns of Reproductive Strategy Parameters in Some Marine Teleost Fishes. J. Fish Biol. 1992, 41, 137–160. [Google Scholar] [CrossRef]
- Pauly, D. Gill Size and Temperature as Governing Factors in Fish Growth: A Generalization of von Bertalanffy’s Growth Formula. Ber. Inst. f. Meereskd. Univ. Kiel 1979, 63, xv + 156 p. [Google Scholar]
- DFO A Harvest Strategy Compliant With the Precautionary Approach. Can. Sci. Advis. Secr. Sci. Advis. Rep. 2006, 2006/023.
- DFO 2018 Assessment of 4VWX Herring. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2018, 2018/052, 1–26.
- DFO Assessment of the West Coast of Newfoundland (NAFO Division 4R) Herring ( Clupea Harengus) Stocks in 2021. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2022, 2022/020, 1–18.
- DFO Assessment of the Quebec North Shore (Division 4S) Herring Stocks in 2020. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2021, 2021/037, 1–15.
- Bourne, C.; Squires, B.; O’Keefe, B.; Schofield, M. Assessment of Newfoundland East and South Coast Atlantic Herring (Clupea Harengus) Stock Complexes to 2018. DFO Can. Sci. Advis. Sec. Res. Doc. 2023, 2023/013, iv + 41 p. [Google Scholar]
- Wheeler, J.P.; Purchase, C.F.; Macdonald, P.D.M.; Fill, R.; Jacks, L.; Wang, H.; Ye, C. Temporal Changes in Maturation, Mean Length-at-Age, and Condition of Spring-Spawning Atlantic Herring (Clupea harengus) in Newfoundland Waters. ICES J. Mar. Sci. 2009, 66, 1800–1807. [Google Scholar] [CrossRef]
- Barrett, T.J.; Hordyk, A.R.; Barrett, M.A.; van den Heuvel, M.R. Spatial and Temporal Differences in Fecundity of Atlantic Herring (Clupea harengus) off Nova Scotia and Consequences for Biological Reference Points. Can. J. Fish. Aquat. Sci. 2022, 79, 1086–1096. [Google Scholar] [CrossRef]
- Burbank, J.; Drake, D.A.R.; Power, M. Silver Shiner, Notropis Photogenis Fecundity and Parasite Infections during the Reproductive Period. Can. J. Zool. 2022, 100, 826–833. [Google Scholar] [CrossRef]
- Messieh, S.N. Fecundity Studies on Atlantic Herring from the Southern Gulf of St. Lawrence and along the Nova Scotia Coast. Trans. Am. Fish. Soc. 1976, 105, 384–394. [Google Scholar] [CrossRef]
- Marshall, D.J.; Bode, M.; Mangel, M.; Arlinghaus, R.; Dick, E.J. Reproductive Hyperallometry and Managing the World’s Fisheries. Proc. Natl. Acad. Sci. USA 2021, 118, e2100695118. [Google Scholar] [CrossRef] [PubMed]
- Óskarsson, G.J. Pre-Spawning Factors and Recruitment Variation in Atlantic Herring (Clupeidae; Clupea harengus, L.): A Comparative Approach. Ph.D. thesis, Dalhousie University, Halifax, NS, Canada, 2005. [Google Scholar]
- Dickey-Collas, M.; Nash, R.D.M.; Brunel, T.; Van Damme, C.J.G.; Marshall, C.T.; Payne, M.R.; Corten, A.; Geffen, A.J.; Peck, M.A.; Hatfield, E.M.C.; et al. Lessons Learned from Stock Collapse and Recovery of North Sea Herring: A Review. ICES J. Mar. Sci. 2010, 67, 1875–1886. [Google Scholar] [CrossRef]
- Burbank, J.; Dejong, R.A.; Turcotte, F.; Rolland, N. Understanding Factors Influencing Atlantic Herring (Clupea harengus) Recruitment: From Egg Deposition to Juveniles. Fish. Oceanogr. 2022, 32, 147–159. [Google Scholar] [CrossRef]
- Óskarsson, G.J.; Taggart, C.T. Variation in Reproductive Potential and Influence on Icelandic Herring Recruitment. Fish. Oceanogr. 2010, 19, 412–426. [Google Scholar] [CrossRef]
- Ottersen, G.; Loeng, H. Covariability in early growth and year-class strength of Barents Sea cod, haddock, and herring: The environmental link. ICES J. Mar. Sci. 2000, 57, 339–348. [Google Scholar] [CrossRef]
- Lorenzen, K.; Enberg, K. Density-dependent growth as a key mechanism in the regulation of fish populations: Evidence from among-population comparisons. Proc. R. Soc. Lond. B 2002, 269, 49–54. [Google Scholar] [CrossRef]
- Stenevik, E.K.; Hølleland, S.; Enberg, K.; Høines, Å.; Salthaug, A.; Slotte, A.; Vatnehol, S.; Aanes, S. Predicting density-dependent somatic growth in Norwegian spring-spawning herring. ICES J. Mar. Sci. 2022, 79, 1340–1352. [Google Scholar] [CrossRef]
- Zimmermann, F.; Ricard, D.; Heino, M. Density regulation in Northeast Atlantic fish populations: Density dependence is stronger in recruitment than in somatic growth. J. Anim. Ecol. 2018, 87, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, F. Environmental effects on recruitment dynamics and population projections of NAFO Division 4TVn Spring Spawning Atlantic Herring. DFO Can. Sci. Advis. Sec. Res. Doc. 2022, 2022/047, iv + 25 p. [Google Scholar]
- Turcotte, F.; Swain, D.P.; McDermid, J.L. NAFO 4TVn Atlantic Herring Population Models: From Virtual Population Analysis to Statistical Catch-at-Age Estimating Time-Varying Natural Mortality. DFO Can. Sci. Advis. Sec. Res. Doc. 2021, 2021/029, vi + 52 p. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | L∞ | K | t0 | L∞LCI | L∞UCI | KLCI | KUCI | t0LCI | t0UCI |
|---|---|---|---|---|---|---|---|---|---|
| 1988 | 345.55 | 0.36 | −1.10 | 342.47 | 348.68 | 0.35 | 0.38 | −1.14 | −1.06 |
| 1989 | 347.90 | 0.35 | −1.34 | 345.67 | 349.92 | 0.33 | 0.36 | −1.40 | −1.29 |
| 1990 | 362.26 | 0.29 | −1.59 | 359.10 | 365.62 | 0.28 | 0.30 | −1.65 | −1.54 |
| 1991 | 354.63 | 0.32 | −1.39 | 352.79 | 356.73 | 0.31 | 0.33 | −1.44 | −1.35 |
| 1992 | 363.22 | 0.26 | −1.98 | 360.81 | 365.58 | 0.25 | 0.26 | −2.06 | −1.90 |
| 1993 | 341.90 | 0.33 | −1.36 | 339.50 | 344.54 | 0.31 | 0.34 | −1.44 | −1.28 |
| 1994 | 346.28 | 0.27 | −1.63 | 343.41 | 349.38 | 0.26 | 0.28 | −1.70 | −1.57 |
| 1995 | 345.92 | 0.28 | −1.63 | 341.55 | 350.42 | 0.27 | 0.29 | −1.71 | −1.57 |
| 1996 | 332.04 | 0.32 | −1.37 | 328.49 | 335.87 | 0.31 | 0.34 | −1.45 | −1.29 |
| 1997 | 331.74 | 0.34 | −1.31 | 329.41 | 334.13 | 0.33 | 0.35 | −1.37 | −1.24 |
| 1998 | 332.23 | 0.34 | −1.47 | 330.42 | 334.06 | 0.33 | 0.35 | −1.56 | −1.39 |
| 1999 | 335.17 | 0.31 | −1.56 | 333.41 | 337.17 | 0.30 | 0.32 | −1.65 | −1.50 |
| 2000 | 337.84 | 0.30 | −1.71 | 335.77 | 339.71 | 0.29 | 0.31 | −1.82 | −1.62 |
| 2001 | 339.13 | 0.29 | −1.83 | 337.57 | 340.73 | 0.29 | 0.30 | −1.90 | −1.77 |
| 2002 | 332.97 | 0.34 | −1.39 | 331.04 | 335.00 | 0.33 | 0.35 | −1.46 | −1.32 |
| 2003 | 337.35 | 0.29 | −2.08 | 334.21 | 340.75 | 0.27 | 0.30 | −2.28 | −1.90 |
| 2004 | 330.85 | 0.34 | −1.35 | 328.41 | 333.43 | 0.33 | 0.35 | −1.43 | −1.29 |
| 2005 | 326.96 | 0.33 | −1.41 | 323.63 | 330.23 | 0.32 | 0.35 | −1.49 | −1.34 |
| 2006 | 315.97 | 0.39 | −1.33 | 311.79 | 320.62 | 0.36 | 0.42 | −1.46 | −1.19 |
| 2007 | 311.77 | 0.42 | −1.13 | 308.20 | 315.47 | 0.39 | 0.44 | −1.23 | −1.04 |
| 2008 | 313.99 | 0.40 | −1.14 | 310.94 | 317.14 | 0.38 | 0.43 | −1.22 | −1.07 |
| 2009 | 311.23 | 0.42 | −1.10 | 307.77 | 314.71 | 0.40 | 0.45 | −1.17 | −1.03 |
| 2010 | 314.96 | 0.35 | −1.31 | 311.86 | 317.98 | 0.33 | 0.37 | −1.39 | −1.23 |
| 2011 | 308.48 | 0.34 | −1.43 | 305.19 | 312.11 | 0.32 | 0.36 | −1.55 | −1.31 |
| 2012 | 303.39 | 0.37 | −1.20 | 298.77 | 308.88 | 0.34 | 0.40 | −1.34 | −1.08 |
| 2013 | 311.40 | 0.33 | −1.30 | 307.87 | 314.55 | 0.32 | 0.35 | −1.41 | −1.21 |
| 2014 | 307.74 | 0.34 | −1.41 | 303.30 | 312.23 | 0.31 | 0.36 | −1.53 | −1.30 |
| 2015 | 291.87 | 0.44 | −1.20 | 288.47 | 295.81 | 0.41 | 0.47 | −1.33 | −1.08 |
| 2016 | 311.61 | 0.33 | −1.39 | 307.83 | 315.94 | 0.31 | 0.36 | −1.51 | −1.26 |
| 2017 | 312.21 | 0.32 | −1.70 | 307.66 | 317.16 | 0.29 | 0.36 | −2.14 | −1.39 |
| 2018 | 312.92 | 0.32 | −1.46 | 308.99 | 316.94 | 0.30 | 0.34 | −1.56 | −1.37 |
| 2019 | 313.94 | 0.30 | −1.76 | 309.72 | 319.20 | 0.28 | 0.32 | −1.91 | −1.60 |
| 2020 | 319.94 | 0.25 | −2.32 | 313.12 | 330.72 | 0.21 | 0.29 | −2.98 | −1.96 |
| 2021 | 311.04 | 0.33 | −1.37 | 307.96 | 314.41 | 0.31 | 0.34 | −1.44 | −1.30 |
| All | 330.86 | 0.34 | −1.42 | 330.36 | 331.37 | 0.33 | 0.34 | −1.43 | −1.41 |
| Year | L∞ | K | t0 | L∞LCI | L∞UCI | KLCI | KUCI | t0LCI | t0UCI |
|---|---|---|---|---|---|---|---|---|---|
| 1988 | 363.45 | 0.35 | −0.70 | 361.01 | 365.91 | 0.33 | 0.36 | −0.82 | −0.59 |
| 1989 | 365.33 | 0.37 | −0.40 | 363.43 | 367.01 | 0.36 | 0.39 | −0.48 | −0.32 |
| 1990 | 371.50 | 0.34 | −0.74 | 370.17 | 372.91 | 0.33 | 0.35 | −0.84 | −0.67 |
| 1991 | 369.54 | 0.34 | −0.68 | 367.93 | 371.28 | 0.33 | 0.36 | −0.85 | −0.51 |
| 1992 | 372.15 | 0.31 | −0.55 | 370.18 | 374.06 | 0.30 | 0.32 | −0.66 | −0.43 |
| 1993 | 377.70 | 0.27 | −0.95 | 375.33 | 380.25 | 0.26 | 0.28 | −1.08 | −0.85 |
| 1994 | 380.62 | 0.25 | −0.91 | 378.63 | 382.57 | 0.24 | 0.25 | −0.96 | −0.86 |
| 1995 | 374.69 | 0.24 | −1.23 | 372.36 | 377.17 | 0.23 | 0.25 | −1.30 | −1.16 |
| 1996 | 363.84 | 0.28 | −0.85 | 361.68 | 365.97 | 0.28 | 0.29 | −0.90 | −0.79 |
| 1997 | 363.05 | 0.28 | −1.08 | 361.33 | 364.97 | 0.27 | 0.29 | −1.19 | −0.99 |
| 1998 | 368.06 | 0.23 | −1.85 | 365.44 | 371.01 | 0.22 | 0.24 | −2.05 | −1.69 |
| 1999 | 346.28 | 0.33 | −0.76 | 344.50 | 348.22 | 0.32 | 0.34 | −0.85 | −0.68 |
| 2000 | 343.57 | 0.35 | −0.71 | 341.88 | 345.20 | 0.34 | 0.36 | −0.78 | −0.64 |
| 2001 | 339.88 | 0.35 | −0.83 | 338.08 | 341.72 | 0.34 | 0.36 | −0.92 | −0.74 |
| 2002 | 335.88 | 0.36 | −0.83 | 334.19 | 338.09 | 0.34 | 0.37 | −0.95 | −0.75 |
| 2003 | 354.39 | 0.23 | −2.43 | 351.00 | 357.48 | 0.22 | 0.25 | −2.68 | −2.15 |
| 2004 | 335.27 | 0.36 | −0.73 | 334.19 | 336.51 | 0.36 | 0.37 | −0.78 | −0.69 |
| 2005 | 336.25 | 0.32 | −1.14 | 334.31 | 339.00 | 0.30 | 0.33 | −1.32 | −1.02 |
| 2006 | 335.03 | 0.34 | −0.97 | 333.23 | 336.91 | 0.32 | 0.35 | −1.10 | −0.87 |
| 2007 | 337.02 | 0.31 | −1.31 | 334.45 | 341.10 | 0.28 | 0.33 | −1.63 | −1.12 |
| 2008 | 328.39 | 0.37 | −0.83 | 327.16 | 329.66 | 0.35 | 0.37 | −0.90 | −0.77 |
| 2009 | 333.42 | 0.33 | −1.08 | 331.55 | 335.64 | 0.31 | 0.34 | −1.24 | −0.97 |
| 2010 | 328.71 | 0.31 | −1.04 | 326.98 | 330.64 | 0.30 | 0.32 | −1.15 | −0.97 |
| 2011 | 338.68 | 0.24 | −1.69 | 335.19 | 343.30 | 0.22 | 0.25 | −1.94 | −1.50 |
| 2012 | 340.48 | 0.23 | −1.97 | 334.21 | 350.55 | 0.19 | 0.25 | −2.58 | −1.59 |
| 2013 | 317.13 | 0.33 | −1.01 | 315.06 | 320.29 | 0.31 | 0.34 | −1.24 | −0.89 |
| 2014 | 310.30 | 0.34 | −1.10 | 307.80 | 313.46 | 0.32 | 0.36 | −1.30 | −0.96 |
| 2015 | 321.12 | 0.30 | −1.41 | 317.59 | 325.38 | 0.27 | 0.32 | −1.71 | −1.15 |
| 2016 | 318.31 | 0.35 | −0.89 | 316.63 | 320.16 | 0.34 | 0.36 | −1.03 | −0.81 |
| 2017 | 335.18 | 0.23 | −2.42 | 327.32 | 344.90 | 0.20 | 0.27 | −3.07 | −1.83 |
| 2018 | 328.60 | 0.27 | −1.28 | 325.28 | 332.93 | 0.25 | 0.28 | −1.54 | −1.11 |
| 2019 | 326.44 | 0.28 | −1.30 | 323.54 | 330.12 | 0.26 | 0.29 | −1.55 | −1.10 |
| 2020 | 326.04 | 0.28 | −1.13 | 321.26 | 331.18 | 0.25 | 0.32 | −1.67 | −0.62 |
| 2021 | 324.84 | 0.31 | −0.99 | 322.91 | 327.36 | 0.30 | 0.33 | −1.14 | −0.90 |
| All | 345.83 | 0.30 | −1.10 | 345.36 | 346.30 | 0.30 | 0.30 | −1.12 | −1.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burbank, J.; McDermid, J.L.; Turcotte, F.; Rolland, N. Temporal Variation in Von Bertalanffy Growth Curves and Generation Time of Southern Gulf of St. Lawrence Spring and Fall Spawning Atlantic Herring (Clupea harengus). Fishes 2023, 8, 205. https://doi.org/10.3390/fishes8040205
Burbank J, McDermid JL, Turcotte F, Rolland N. Temporal Variation in Von Bertalanffy Growth Curves and Generation Time of Southern Gulf of St. Lawrence Spring and Fall Spawning Atlantic Herring (Clupea harengus). Fishes. 2023; 8(4):205. https://doi.org/10.3390/fishes8040205
Chicago/Turabian StyleBurbank, Jacob, Jenni L. McDermid, François Turcotte, and Nicolas Rolland. 2023. "Temporal Variation in Von Bertalanffy Growth Curves and Generation Time of Southern Gulf of St. Lawrence Spring and Fall Spawning Atlantic Herring (Clupea harengus)" Fishes 8, no. 4: 205. https://doi.org/10.3390/fishes8040205
APA StyleBurbank, J., McDermid, J. L., Turcotte, F., & Rolland, N. (2023). Temporal Variation in Von Bertalanffy Growth Curves and Generation Time of Southern Gulf of St. Lawrence Spring and Fall Spawning Atlantic Herring (Clupea harengus). Fishes, 8(4), 205. https://doi.org/10.3390/fishes8040205