
Effects of IGF1 and IGF2 on In Vitro Ovarian Follicle Maturation in Rainbow Trout, Oncorhynchus mykiss


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Husbandry
2.2. Tissue Collection and In Vitro Bioassay
2.3. Experimental Design
2.4. Statistical Analysis
3. Results
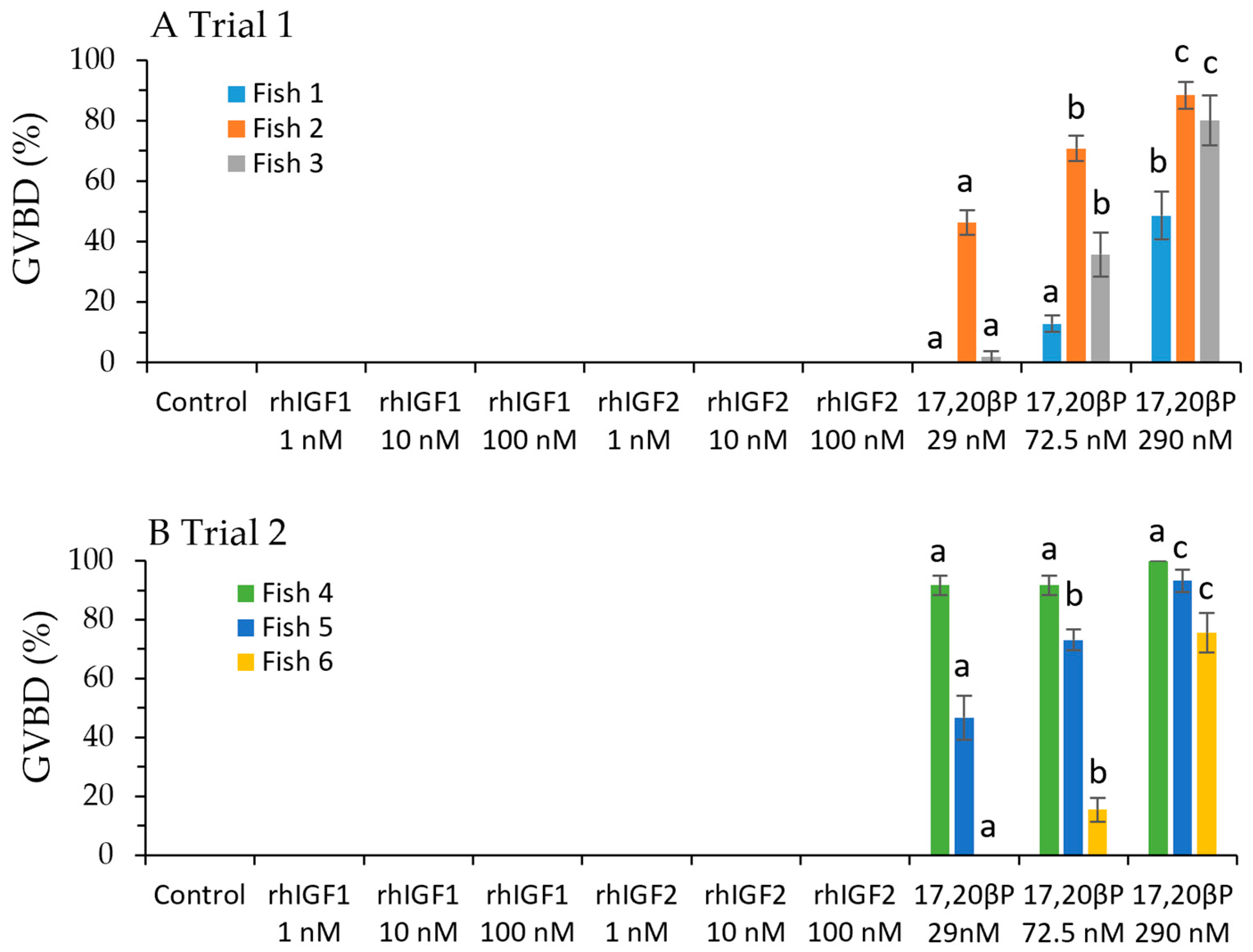
3.1. Effects of rhIGF1 and rhIGF2 on Induction of GVBD In Vitro
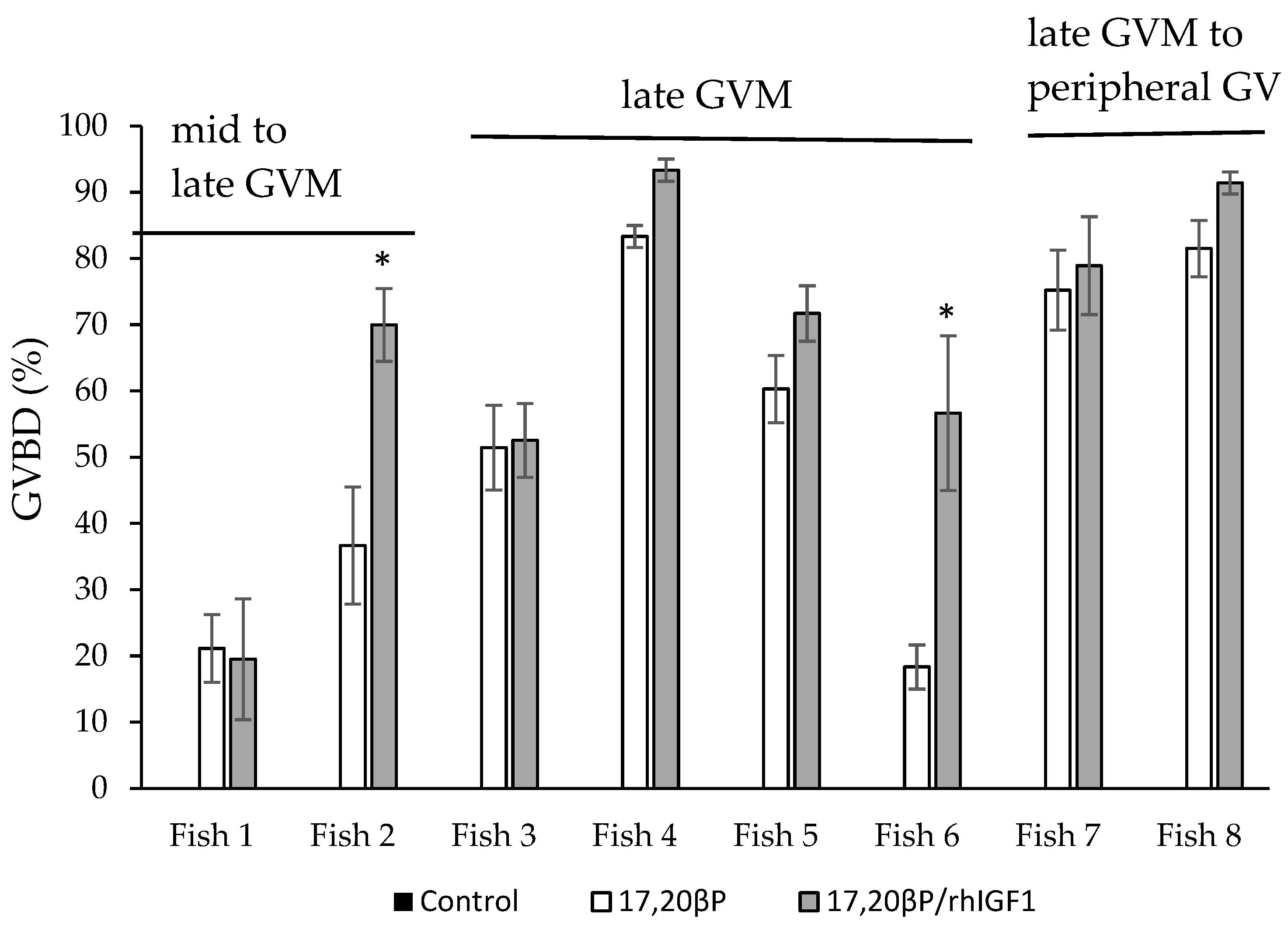
3.2. Effects of rhIGF1 on MIH-Induced GVBD In Vitro
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patiño, R.; Yoshizaki, G.; Thomas, P.; Kagawa, H. Gonadotropic control of ovarian follicle maturation: The two-stage concept and its mechanisms. Comp. Biochem. Physiol. Part B 2001, 129, 427–439. [Google Scholar] [CrossRef]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50, S195–S219. [Google Scholar]
- Lubzens, E.; Young, G.; Bobe, J.; Cerdà, J. Oogenesis in teleosts: How fish eggs are formed. Gen. Comp. Endocrinol. 2010, 165, 367–389. [Google Scholar]
- Mohajer, L.E.; Bulteau, R.; Fontaine, P.; Milla, S. Maturation Inducing Hormones in teleosts: Are progestogens always the first to be nominated? Aquaculture 2022, 546, 737315. [Google Scholar]
- Lubzens, E.; Bobe, J.; Young, G.; Sullivan, C.V. Maternal investment in fish oocytes and eggs: The molecular cargo and its contributions to fertility and early development. Aquaculture 2017, 472, 107–143. [Google Scholar]
- Das, D.; Khan, P.P.; Maitra, S. Endocrine and paracrine regulation of meiotic cell cycle progression in teleost oocytes: cAMP at the centre of complex intra-oocyte signalling events. Gen. Comp. Endocrinol. 2017, 241, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Reinecke, M. Insulin-like growth factors and fish reproduction. Biol. Reprod. 2010, 82, 656–661. [Google Scholar] [PubMed] [Green Version]
- Li, J.; Liu, Z.; Kang, T.; Li, M.; Wang, D.; Cheng, C.H.K. Igf3: A novel player in fish reproduction. Biol. Reprod. 2021, 104, 1194–1204. [Google Scholar]
- Ndandala, C.B.; Dai, M.; Mustapha, U.F.; Li, X.; Liu, J.; Huang, H.; Li, G.; Chen, H. Current research and future perspectives of GH and IGFs family genes in somatic growth and reproduction of teleost fish. Aquac. Rep. 2022, 26, 101289. [Google Scholar] [CrossRef]
- Bobe, J.; Maugars, G.; Nguyen, T.; Rime, H.; Jalabert, B. Rainbow trout follicular maturational competence acquisition is associated with an increased expression of follicle stimulating hormone receptor and insulin-like growth factor 2 messenger RNAs. Mol. Reprod. Dev. 2003, 66, 46–53. [Google Scholar] [CrossRef]
- Bobe, J.; Nguyen, T.; Jalabert, B. Targeted gene expression profiling in the rainbow trout (Oncorhynchus mykiss) ovary during maturational competence acquisition and oocyte maturation. Biol. Reprod. 2004, 71, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamangar, B.B.; Gabillard, J.C.; Bobe, J. Insulin-like growth factor-binding protein (IGFBP)-1,-2,-3,-4,-5, and -6 and IGFBP-related protein 1 during rainbow trout postvitellogenesis and oocyte maturation: Molecular characterization, expression profiles, and hormonal regulation. Endocrinology 2006, 147, 2399–2410. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yu, S.M.; Ge, W. Expression and functional characterization of intrafollicular GH-IGF system in the zebrafish ovary. Gen. Comp. Endocrinol. 2016, 232, 32–42. [Google Scholar] [CrossRef]
- Maestro, M.A.; Planas, J.V.; Moriyama, S.; Gutiérrez, J.; Planas, J.; Swanson, P. Ovarian receptors for insulin and insulin-like growth factor I (IGF-I) and effects of IGF-I on steroid production by isolated follicular layers of the preovulatory coho salmon ovarian follicle. Gen. Comp. Endocrinol. 1997, 106, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Maestro, M.A.; Méndez, E.; Planas, J.V.; Gutiérrez, J.; Swanson, P. Dynamics of insulin and insulin-like growth factor-I (IGF-I) ovarian receptors during maturation in the brown trout (Salmo trutta). Fish Physiol. Biochem. 1999, 20, 341–349. [Google Scholar] [CrossRef]
- Young, G.; Adachi, S.; Nagahama, Y. Role of ovarian theca-interstitial layer and granulosa cells in gonadotropin induced synthesis of a salmonid maturation-inducing substance (17α, 20β-dihydroxy-4-pregnen-3-one). Dev. Biol. 1986, 118, 1–8. [Google Scholar] [CrossRef]
- Nagahama, Y. Gonadotropin action on gametogenesis and steroidogenesis in teleost gonads. Zool. Sci. 1987, 4, 209–222. [Google Scholar]
- Maestro, M.A.; Planas, J.V.; Gutiérrez, J.; Moriyama, S.; Swanson, P. Effects of insulin-like growth-factor-I (Igf-I) on steroid-production by isolated ovarian theca and granulosa layers of preovulatory coho salmon. Neth. J. Zool. 1995, 45, 143–146. [Google Scholar]
- Weber, G.M.; Sullivan, C.V. Effects of insulin-like growth factor-I on in vitro final oocyte maturation and ovarian steroidogenesis in striped bass, Morone saxatilis. Biol. Reprod. 2000, 63, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.M.; Sullivan, C.V. Insulin-like growth factor-I induces oocyte maturational competence but not meiotic resumption in white bass (Morone chrysops) follicles in vitro: Evidence for rapid evolution of Insulin-like growth factor action. Biol. Reprod. 2005, 72, 1177–1186. [Google Scholar]
- Weber, G.M.; Moore, A.B.; Sullivan, C.V. In vitro actions of insulin-like growth factor-I on ovarian follicle maturation in white perch (Morone americana). Gen. Comp. Endocrinol. 2007, 151, 180–187. [Google Scholar] [CrossRef]
- Nelson, S.N.; Van Der Kraak, G. The role of the insulin-like growth factor (IGF) system in zebrafish (Danio rerio) ovarian development. Gen. Comp. Endocrinol. 2010, 168, 103–110. [Google Scholar] [CrossRef]
- Das, D.; Pal, S.; Maitra, S. Releasing prophase arrest in zebrafish oocyte: Synergism between maturational steroid and Igf1. Reproduction 2016, 151, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chu, L.; Sun, X.; Liu, Y.; Cheng, C.H.K. IGFs mediate the action of LH on oocyte maturation in zebrafish. Mol. Endocrinol. 2015, 29, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, H.; Kobayashi, M.; Hasegawa, Y.; Aida, K. Insulin and insulin-like growth factors I and II induce final maturation of oocytes of red seabream, Pagrus major, in vitro. Gen. Comp. Endocrinol. 1994, 95, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Mukherjee, D.; Sen, U.; Paul, S.; Bhattacharyya, S.P. In vitro effects of insulin-like growth factors and insulin on oocyte maturation and maturation-inducing steroid production in ovarian follicles of common carp, Cyprinus carpio. Comp. Biochem. Physiol. Part A 2006, 144, 63–77. [Google Scholar] [CrossRef]
- Dasgupta, S.; Basu, D.; Ravi Kumar, L.; Bhattacharya, S. Insulin alone can lead to a withdrawal of meiotic arrest in the carp oocyte. J. Biosci. 2001, 26, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Chourasia, T.K.; Joy, K.P. Estrogen-2/4-hydroxylase activity is stimulated during germinal vesicle breakdown induced by hCG, IGF-1, GH and insulin in the catfish Heteropneustes fossilis. Gen. Comp. Endocrinol. 2008, 155, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Hajra, S.; Das, D.; Ghosh, P.; Pal, S.; Nath, P.; Maitra, S. Regulation of recombinant human insulin-induced maturational events in Clarias batrachus (L.) oocytes in vitro. Zygote 2016, 24, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Pramanick, K.; Mukherjee, D.; Maiti, B.R. In vitro induction of oocyte maturation and steroidogenesis by gonadotropins, insulin, calcitonin and growth factor in an estuarine flat head grey mullet, Mugil cephalus L. Fish Physiol. Biochem. 2014, 40, 105–116. [Google Scholar] [CrossRef]
- Picha, M.E.; Shi, B.; Thomas, P. Dual role of IGF-II in oocyte maturation in southern flounder Paralichthys lethostigma: Up-regulation of mPRα and resumption of meiosis. Gen. Comp. Endocrinol. 2012, 177, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Negatu, Z.; Hsiao, S.M.; Wallace, R.A. Effects of insulin-like growth factor-I on final oocyte maturation and steroid production in Fundulus heteroclitus. Fish Physiol. Biochem. 1998, 19, 13–21. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Luckenbach, A.; Middleton, M.M.; Swanson, P. The spatiotemporal expression of multiple coho salmon ovarian connexin genes and their hormonal regulation in vitro during oogenesis. Reprod. Biol. Endocrinol. 2011, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalabert, B.; Fostier, A. The modulatory effect in vitro of oestradiol-17β, testosterone or cortisol on the output of 17α-hydroxy-20β-dihydroprogesterone by rainbow trout (Salmo gairdneri) ovarian follicles stimulated by the maturational gonadotropin s-GtH. Reprod. Nutr. Dev. 1984, 24, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Lankford, S.E.; Weber, G.M. The maturation-inducing hormone 17α,20β-dihydroxy-4-pregnen-3-one regulates gene expression of inhibin βA and bambi (bone morphogenetic protein and activin-membrane-bound inhibitor) in the rainbow trout ovary. Gen. Comp. Endocrinol. 2010, 168, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Patiño, R.; Kagawa, H. Regulation of gap junctions and oocyte maturational competence by gonadotropin and insulin-like growth factor-I in ovarian follicles of red seabream. Gen. Comp. Endocrinol. 1999, 115, 454–462. [Google Scholar] [CrossRef]
- Thomas, P.; Pinter, J.; Das, S. Upregulation of the maturation-inducing steroid membrane receptor in spotted seatrout ovaries by gonadotropin during oocyte maturation and its physiological significance. Biol. Reprod. 2001, 64, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, H.; Takagi, M. Design of serum-free medium for suspension culture of CHO cells on the basis of general commercial media. Cytotechnology 2015, 67, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, I.; Kusakabe, M.; Swanson, P.; Young, G. Regulation of sex steroid production and mRNAs encoding gonadotropin receptors and steroidogenic proteins by gonadotropins, cyclic AMP and insulin-like growth factor-I in ovarian follicles of rainbow trout (Oncorhynchus mykiss) at two stages of vitellogenesis. Comp. Biochem. Physiol. Part A 2016, 201, 132–140. [Google Scholar]



{kind=link}
{kind=link}
| Fish | Order | Reproduction Strategy | OMC Response (IGF Ligands) | OM Response (IGF Ligands) | Citations |
|---|---|---|---|---|---|
| Zebrafish (Danio rerio) | Cypriniformes | AS | Yes (IGF1) No (IGF2) | Yes (IGF1,2a,2b,3) | [22,23,24] |
| Red seabream (Pagrus major) | Perciformes | AS | Yes (IGF1,2) | Yes (IGF1,2) | [25] |
| Rainbow trout (Oncorhynchus mykiss) | Salmoniformes | GS-SC | No (IGF1) | No (IGF1) | Present |
| Common carp (Cyprinus carpio) | Cypriniformes | GS-SC | No (IGF1) | Yes (IGF1,2) | [26] |
| Rohu Indian carp (Labeo rohita) | Cypriniformes | GS-SC | ND | Yes (IGF1) | [27] |
| Stinging catfish (Heteropneustes fossilis) | Siluriformes | GS-SC | ND | Yes (IGF1) | [28] |
| Walking catfish (Clarias batrachus (L.)) | Siluriformes | GS-SC | ND | Yes (IGF1) | [29] |
| Grey mullet (Mugil cephalus) | Mugiliformes | GS-SC | ND | Yes (IGF1) | [30] |
| Striped bass (Morone saxatilis) | Perciformes | GS-SC | No (IGF1,2) | Yes (IGF1,2) | [19] |
| Southern flounder (Paralichthys lethostigma) | Pleuronectiformes | GS-MC | Yes (IGF2), No (IGF1) | Yes (IGF1,2) | [31] |
| Mummichog (Fundulus heteroclitus) | Cyprinodontiformes | GS-MC | ND | Yes (IGF1,2) | [32] |
| White bass (Morone chrysops) | Perciformes | GS-MC | Yes (IGF1) | No (IGF1) | [20] |
| White perch (Morone americana) | Perciformes | GS-MC | Yes (IGF1,2) | No (IGF1,2) | [21] |
| Spotted seatrout (Cynoscion nebulosus) | Acanthuriformes | GS-MC | Yes (IGF1) | ND | [33] |
| Mid GVM | Late GVM | Peripheral GV | Follicle Size (mm) Mean ± SEM | |
|---|---|---|---|---|
| Trial 1 | ||||
| Fish 1 | 5 | 20 | 5.09 ± 0.03 | |
| Fish 2 | 25 | 5.15 ± 0.05 | ||
| Fish 3 | 24 | 5.04 ± 0.04 | ||
| Trial 2 | ||||
| Fish 4 | 11 | 9 | 5.42 ± 0.04 | |
| Fish 5 | 7 | 13 | 5.09 ± 0.04 | |
| Fish 6 | 4 | 16 | 5.24 ± 0.03 |
| Stage | n | Follicle Size Range (mm) | Follicle Size (mm) Mean ± SEM |
|---|---|---|---|
| Incompetent | |||
| Central GV | 1 | 3.2 | - |
| Early to mid GVM | 6 | 3.00–4.21 | 3.60 ± 0.17 |
| Mid GVM | 14 | 2.99–4.49 | 3.97 ± 0.10 |
| Mid to late GVM | 5 | 3.65–4.36 | 3.99 ± 0.12 |
| All | 3.86 ± 0.08 | ||
| Competent | |||
| Mid to late GVM | 2 | 4.07–4.83 | 4.45 ± 0.38 |
| Late GVM | 4 | 3.96–4.96 | 4.37 ± 0.23 |
| Late GVM to peripheral GV | 2 | 4.02–4.05 | 4.04 ± 0.01 |
| All | 4.30 ± 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, G.M. Effects of IGF1 and IGF2 on In Vitro Ovarian Follicle Maturation in Rainbow Trout, Oncorhynchus mykiss. Fishes 2023, 8, 367. https://doi.org/10.3390/fishes8070367
Weber GM. Effects of IGF1 and IGF2 on In Vitro Ovarian Follicle Maturation in Rainbow Trout, Oncorhynchus mykiss. Fishes. 2023; 8(7):367. https://doi.org/10.3390/fishes8070367
Chicago/Turabian StyleWeber, Gregory M. 2023. "Effects of IGF1 and IGF2 on In Vitro Ovarian Follicle Maturation in Rainbow Trout, Oncorhynchus mykiss" Fishes 8, no. 7: 367. https://doi.org/10.3390/fishes8070367
APA StyleWeber, G. M. (2023). Effects of IGF1 and IGF2 on In Vitro Ovarian Follicle Maturation in Rainbow Trout, Oncorhynchus mykiss. Fishes, 8(7), 367. https://doi.org/10.3390/fishes8070367