


Genetic Population Structure and Diversity of the Whitetail Dogfish Squalus albicaudus (Chondrichthyes, Squaliformes) along the Brazilian Coast as Identified by SNP Markers

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Collection and DNA Extraction
2.2. ddRAD Library Preparation
2.3. SNP Analysis Filtering
2.4. Genetic Diversity and Population Structure
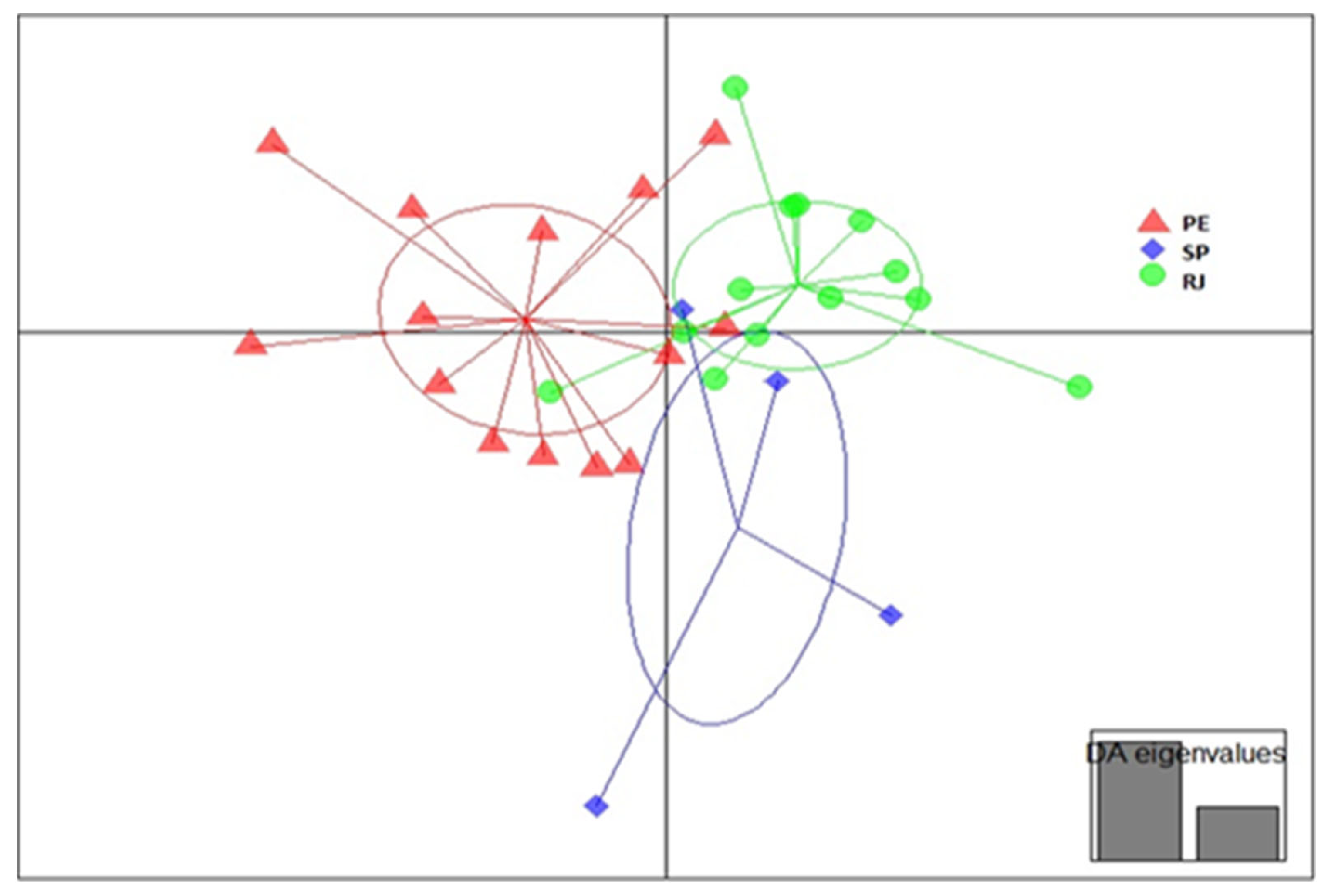
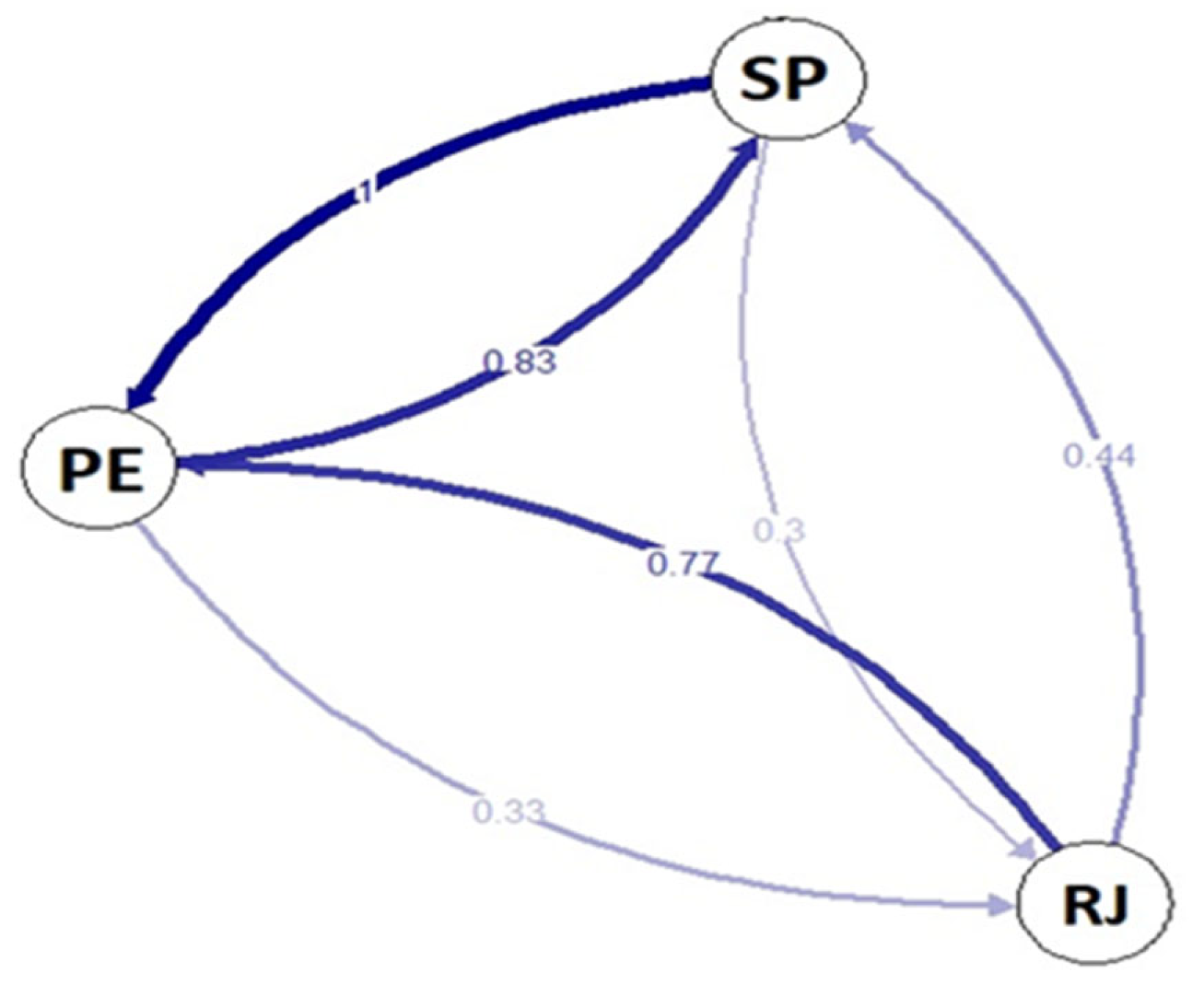
3. Results
Genetic Diversity and Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing Drives over One-Third of All Sharks and Rays toward a Global Extinction Crisis. Curr. Biol. 2021, 31, 4773–4787.e8. [Google Scholar] [CrossRef] [PubMed]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P.; et al. Half a Century of Global Decline in Oceanic Sharks and Rays. Nature 2021, 589, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Habibullah, M.S.; Din, B.H.; Tan, S.-H.; Zahid, H. Impact of Climate Change on Biodiversity Loss: Global Evidence. Environ. Sci. Pollut. Res. 2022, 29, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Cortés, E. Life History Patterns and Correlations in Sharks. Rev. Fish. Sci. 2000, 8, 299–344. [Google Scholar] [CrossRef]
- Conrath, C.; Musick, J. Reproductive Biology of Elasmobranchs. Biol. Sharks Relat. 2012, 2, 291–311. [Google Scholar]
- Allendorf, F.; England, P.; Luikart, G.; Ritchie, P.; Ryman, N. Genetic Effects of Harvest on Wild Animal Populations. Trends Ecol. Evol. 2008, 23, 327–337. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.M.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.; Fordham, S.V.; Francis, M.P.; et al. Extinction Risk and Conservation of the World’s Sharks and Rays. eLife 2014, 3, e00590. [Google Scholar] [CrossRef] [Green Version]
- MacNeil, M.A.; Chapman, D.D.; Heupel, M.; Simpfendorfer, C.A.; Heithaus, M.; Meekan, M.; Harvey, E.; Goetze, J.; Kiszka, J.; Bond, M.E.; et al. Global Status and Conservation Potential of Reef Sharks. Nature 2020, 583, 801–806. [Google Scholar] [CrossRef]
- Boussarie, G.; Momigliano, P.; Robbins, W.D.; Bonnin, L.; Cornu, J.; Fauvelot, C.; Kiszka, J.J.; Manel, S.; Mouillot, D.; Vigliola, L. Identifying Barriers to Gene Flow and Hierarchical Conservation Units from Seascape Genomics: A Modelling Framework Applied to a Marine Predator. Ecography 2022, 2022, e06158. [Google Scholar] [CrossRef]
- Sherman, C.S.; Simpfendorfer, C.A.; Pacoureau, N.; Matsushiba, J.H.; Yan, H.F.; Walls, R.H.L.; Rigby, C.L.; VanderWright, W.J.; Jabado, R.W.; Pollom, R.A.; et al. Half a Century of Rising Extinction Risk of Coral Reef Sharks and Rays. Nat. Commun. 2023, 14, 15. [Google Scholar] [CrossRef]
- Hohenlohe, P.A.; Funk, W.C.; Rajora, O.P. Population Genomics for Wildlife Conservation and Management. Mol. Ecol. 2021, 30, 62–82. [Google Scholar] [CrossRef] [PubMed]
- Delaval, A.; Frost, M.; Bendall, V.; Hetherington, S.J.; Stirling, D.; Hoarau, G.; Jones, C.S.; Noble, L.R. Population and Seascape Genomics of a Critically Endangered Benthic Elasmobranch, the Blue Skate Dipturus Batis. Evol. Appl. 2022, 15, 78–94. [Google Scholar] [CrossRef] [PubMed]
- Hogg, C.J.; Ottewell, K.; Latch, P.; Rossetto, M.; Biggs, J.; Gilbert, A.; Richmond, S.; Belov, K. Threatened Species Initiative: Empowering Conservation Action Using Genomic Resources. Proc. Natl. Acad. Sci. USA 2022, 119, e2115643118. [Google Scholar] [CrossRef] [PubMed]
- Pollom, R.; Rincon, G.; Herman, K. Squalus albicaudus. IUCN Red List. Threat. Species 2020, E.T129495269A129495382. [Google Scholar] [CrossRef]
- Casselberry, G.A.; Carlson, J.K. Endangered Species Act Status Review of the Daggernose Shark (Isogomphodon Oxyrhynchus). 2015. Available online: https://repository.library.noaa.gov/view/noaa/17700 (accessed on 20 February 2023).
- Lessa, R.; Batista, V.S.; Santana, F.M. Close to Extinction? The Collapse of the Endemic Daggernose Shark (Isogomphodon oxyrhynchus) off Brazil. Glob. Ecol. Conserv. 2016, 7, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Villwock de Miranda, L.; Vooren, C. Captura e Esforço Da Pesca de Elasmobranquios Demersais No Sul Oe Brasil Nos Anos 1975 a 1997.Catch and Effort of Demersal Elasmobranchs in South Brazil from 1975 to 1997. Frente Marítimo 2003, 19, 217–231. [Google Scholar]
- Lessa, R.; Vooren, C.M. Rhinobatos horkelii. IUCN Red List Threat. Species 2007, E.T41064A10396152. [Google Scholar]
- Feitosa, L.M.; Martins, A.P.B.; Giarrizzo, T.; Macedo, W.; Monteiro, I.L.; Gemaque, R.; Nunes, J.L.S.; Gomes, F.; Schneider, H.; Sampaio, I. DNA-Based Identification Reveals Illegal Trade of Threatened Shark Species in a Global Elasmobranch Conservation Hotspot. Sci. Rep. 2018, 8, 3347. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, M.; Solé-Cava, A.M.; Henning, F. What’s in a Name? Phylogenetic Species Identification Reveals Extensive Trade of Endangered Guitarfishes and Sharks. Biol. Conserv. 2021, 257, 109119. [Google Scholar] [CrossRef]
- da Silva Ferrette, B.L.; Mendonca, F.F.; Coelho, R.; de Oliveira, P.G.V.; Hazin, F.H.V.; Romanov, E.V.; Oliveira, C.; Santos, M.N.; Foresti, F. High Connectivity of the Crocodile Shark between the Atlantic and Southwest Indian Oceans: Highlights for Conservation. PLoS ONE 2015, 10, e0117549. [Google Scholar] [CrossRef] [Green Version]
- Bunholi, I.V.; da Silva Ferrette, B.L.; De Biasi, J.B.; de Oliveira Magalhães, C.; Rotundo, M.M.; Oliveira, C.; Foresti, F.; Mendonça, F.F. The Fishing and Illegal Trade of the Angelshark: DNA Barcoding against Misleading Identifications. Fish. Res. 2018, 206, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Ferrette, B.L.d.S.; Domingues, R.R.; Rotundo, M.M.; Miranda, M.P.; Bunholi, I.V.; De Biasi, J.B.; Oliveira, C.; Foresti, F.; Mendonça, F.F. DNA Barcode Reveals the Bycatch of Endangered Batoids Species in the Southwest Atlantic: Implications for Sustainable Fisheries Management and Conservation Efforts. Genes 2019, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Guimarães-Costa, A.; Machado, F.S.; Reis-Filho, J.A.; Andrade, M.; Araújo, R.G.; Corrêa, E.M.R.; Sampaio, I.; Giarrizzo, T. DNA Barcoding for the Assessment of the Taxonomy and Conservation Status of the Fish Bycatch of the Northern Brazilian Shrimp Trawl Fishery. Front. Mar. Sci. 2020, 7, 566021. [Google Scholar] [CrossRef]
- Bernardo, C.; Adachi, A.M.C.L.; Paes da Cruz, V.; Foresti, F.; Loose, R.H.; Bornatowski, H. The Label “Cação” Is a Shark or a Ray and Can Be a Threatened Species! Elasmobranch Trade in Southern Brazil Unveiled by DNA Barcoding. Mar. Policy 2020, 116, 103920. [Google Scholar] [CrossRef]
- Viana, S.T.D.F.; Carvalho, M.R.D.; Gomes, U.L. Taxonomy and Morphology of Species of the Genus Squalus Linnaeus, 1758 from the Southwestern Atlantic Ocean (Chondrichthyes: Squaliformes: Squalidae). Zootaxa 2016, 4133, 1. [Google Scholar] [CrossRef]
- Rincon, G.; Mazzoleni, R.C.; Palmeira, A.R.O.; Lessa, R. Deep-Water Sharks, Rays, and Chimaeras of Brazil. In Chondrichthyes: Multidisciplinary Approach’; Rodrigues-Filho, L.F., Sales, B.D., Eds.; InTech: Rijeka, Croatia, 2017; ISBN 978-953-51-3711-5. [Google Scholar]
- Sabadin, D.E.; Lucifora, L.O.; Barbini, S.A.; Figueroa, D.E.; Kittlein, M. Towards Regionalization of the Chondrichthyan Fauna of the Southwest Atlantic: A Spatial Framework for Conservation Planning. ICES J. Mar. Sci. 2020, 77, 1893–1905. [Google Scholar] [CrossRef]
- Ebert, D.A.; Dando, M.; Fowler, S. Sharks of the World: A Complete Guide; Princeton University Press: Princeton, NJ, USA, 2021; Volume 19, ISBN 0-691-20599-X. [Google Scholar]
- Merten Cruz, M.; Szynwelski, B.E.; Ochotorena de Freitas, T.R. Biodiversity on Sale: The Shark Meat Market Threatens Elasmobranchs in Brazil. Aquat. Conserv. 2021, 31, 3437–3450. [Google Scholar] [CrossRef]
- Hauser-Davis, R.A.; Pereira, C.F.; Pinto, F.; Torres, J.P.M.; Malm, O.; Vianna, M. Mercury Contamination in the Recently Described Brazilian White-Tail Dogfish Squalus albicaudus (Squalidae, Chondrichthyes). Chemosphere 2020, 250, 126228. [Google Scholar] [CrossRef]
- Luikart, G.; England, P.R.; Tallmon, D.; Jordan, S.; Taberlet, P. The Power and Promise of Population Genomics: From Genotyping to Genome Typing. Nat. Rev. Genet. 2003, 4, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Rajora, O.P. Population Genomics: Concepts, Approaches and Applications, 1st ed.; Spring International Publishing: Basileia, Swiss, 2019; ISBN 978-3-030-04587-6. [Google Scholar]
- Allendorf, F.W.; Funk, W.C.; Aitken, S.N.; Byrne, M.; Luikart, G. 113 Small Populations and Genetic Drift. In Conservation and the Genomics of Populations, 3rd ed.; Allendorf, F.W., Funk, W.C., Aitken, S.N., Byrne, M., Luikart, G., Antunes, A., Eds.; Oxford University Press: Oxford, UK, 2022; ISBN 978-0-19-885656-6. [Google Scholar]
- Cermakova, E.; Lencova, S.; Mukherjee, S.; Horka, P.; Vobruba, S.; Demnerova, K.; Zdenkova, K. Identification of Fish Species and Targeted Genetic Modifications Based on DNA Analysis: State of the Art. Foods 2023, 12, 228. [Google Scholar] [CrossRef]
- Kumar, G.; Kocour, M. Applications of Next-Generation Sequencing in Fisheries Research: A Review. Fish. Res. 2017, 186, 11–22. [Google Scholar] [CrossRef]
- Grummer, J.A.; Beheregaray, L.B.; Bernatchez, L.; Hand, B.K.; Luikart, G.; Narum, S.R.; Taylor, E.B. Aquatic Landscape Genomics and Environmental Effects on Genetic Variation. Trends Ecol. Evol. 2019, 34, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, M.; Yap, J.-Y.S.; Lemmon, J.; Bain, D.; Bragg, J.; Hogbin, P.; Gallagher, R.; Rutherford, S.; Summerell, B.; Wilson, T.C. A Conservation Genomics Workflow to Guide Practical Management Actions. Glob. Ecol. Conserv. 2021, 26, e01492. [Google Scholar] [CrossRef]
- Hoban, S.; Bruford, M.W.; da Silva, J.M.; Funk, W.C.; Frankham, R.; Gill, M.J.; Grueber, C.E.; Heuertz, M.; Hunter, M.E.; Kershaw, F.; et al. Genetic Diversity Goals and Targets Have Improved, but Remain Insufficient for Clear Implementation of the Post-2020 Global Biodiversity Framework. Conserv. Genet. 2023, 24, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Daly-Engel, T.; Grubbs, R.; Feldheim, K.; Bowen, B.; Toonen, R. Is Multiple Mating Beneficial or Unavoidable? Low Multiple Paternity and Genetic Diversity in the Shortspine Spurdog Squalus mitsukurii. Mar. Ecol. Prog. Ser. 2010, 403, 255–267. [Google Scholar] [CrossRef]
- Lamarca, F.; Vianna, M.; Vilasboa, A. The First Reproductive Parameters and Evidence of Multiple Paternity in One New Spiny Dogfish Species, Squalus albicaudus ( Squaliformes, Squalidae). J. Fish Biol. 2020, 97, 1268–1272. [Google Scholar] [CrossRef]
- Lage, C.R.; Petersen, C.W.; Forest, D.; Barnes, D.; Kornfield, I.; Wray, C. Evidence of Multiple Paternity in Spiny Dogfish (Squalus acanthias) Broods Based on Microsatellite Analysis. J. Fish Biol. 2008, 73, 2068–2074. [Google Scholar] [CrossRef]
- Veríssimo, A.; Mcdowell, J.R.; Graves, J.E. Global Population Structure of the Spiny Dogfish Squalus acanthias, a Temperate Shark with an Antitropical Distribution. Mol. Ecol. 2010, 19, 1651–1662. [Google Scholar] [CrossRef]
- Craven, K.S.; Webb, C.; Ragsdale, A.K.; Schrey, A.W. A Pilot Study of Multiple Paternity in Three Litters of Spiny Dogfish (Squalus acanthias) off the South Carolina/Georgia Coast. BIOS 2018, 89, 23–28. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and Extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Ariza, A.A.; Adachi, A.M.; Roque, P.; Hazin, F.H.; Vianna, M.; Rotundo, M.M.; Delpiani, S.M.; de Astarloa, J.M.D.; Delpiani, G.; Oliveira, C. DNA Barcoding and Species Delimitation for Dogfish Sharks Belonging to the Squalus Genus (Squaliformes: Squalidae). Diversity 2022, 14, 544. [Google Scholar] [CrossRef]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double Digest RADseq: An Inexpensive Method for de Novo SNP Discovery and Genotyping in Model and Non-Model Species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.; Conn, J.E.; Alonso, D.P.; Vinetz, J.M.; Emerson, K.J.; Ribolla, P.E.M. Microgeographical Structure in the Major Neotropical Malaria Vector Anopheles Darlingi Using Microsatellites and SNP Markers. Parasites Vectors 2017, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics; Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An Analysis Tool Set for Population Genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Rousset, F. Genepop’007: A Complete Re-implementation of the Genepop Software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New Tools for the Analysis of Genome-Wide SNP Data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Puechmaille, S.J. The Program Structure Does Not Reliably Recover the Correct Population Structure When Sampling Is Uneven: Subsampling and New Estimators Alleviate the Problem. Mol. Ecol. Resour. 2016, 16, 608–627. [Google Scholar] [CrossRef]
- Li, M.; Liu, J.; Yang, W.; Sun, X.; Guo, Z. Structure-Revealing Low-Light Image Enhancement via Robust Retinex Model. IEEE Trans. Image Process. 2018, 27, 2828–2841. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A Program for Identifying Clustering Modes and Packaging Population Structure Inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. DiveRsity: An R Package for the Estimation and Exploration of Population Genetics Parameters and Their Associated Errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, L.; Keenan, K.; Zackrisson, M.; Prodöhl, P.; Kleinhans, D. Directional Genetic Differentiation and Relative Migration. Ecol. Evol. 2016, 6, 3461–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junge, C.; Donnellan, S.C.; Huveneers, C.; Bradshaw, C.J.A.; Simon, A.; Drew, M.; Duffy, C.; Johnson, G.; Cliff, G.; Braccini, M.; et al. Comparative Population Genomics Confirms Little Population Structure in Two Commercially Targeted Carcharhinid Sharks. Mar. Biol. 2019, 166, 16. [Google Scholar] [CrossRef]
- Janes, J.K.; Miller, J.M.; Dupuis, J.R.; Malenfant, R.M.; Gorrell, J.C.; Cullingham, C.I.; Andrew, R.L. The K = 2 Conundrum. Mol. Ecol. 2017, 26, 3594–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.; Wen, W. Documentation for STRUCTURE Software: Version 2; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software STRUCTURE: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Gandra, M.; Assis, J.; Martins, M.R.; Abecasis, D. Reduced Global Genetic Differentiation of Exploited Marine Fish Species. Mol. Biol. Evol. 2021, 38, 1402–1412. [Google Scholar] [CrossRef]
- Shepherd, T.D.; Myers, R.A. Direct and Indirect Fishery Effects on Small Coastal Elasmobranchs in the Northern Gulf of Mexico. Ecol. Lett. 2005, 8, 1095–1104. [Google Scholar] [CrossRef]
- Hazin, F.H.; Fischer, A.F.; Broadhurst, M.K.; Veras, D.; Oliveira, P.G.; Burgess, G.H. Notes on the Reproduction of Squalus megalops off Northeastern Brazil. Fish. Res. 2006, 79, 251–257. [Google Scholar] [CrossRef]
- Jones, P.J. A Governance Analysis of the Galápagos Marine Reserve. Mar. Policy 2013, 41, 65–71. [Google Scholar] [CrossRef] [Green Version]
- van Oppen, M.J.H.; Coleman, M.A. Advancing the Protection of Marine Life through Genomics. PLoS Biol. 2022, 20, e3001801. [Google Scholar] [CrossRef] [PubMed]
- Domingues, R.R.; Hilsdorf, A.W.S.; Gadig, O.B.F. The Importance of Considering Genetic Diversity in Shark and Ray Conservation Policies. Conserv. Genet. 2018, 19, 501–525. [Google Scholar] [CrossRef] [Green Version]
- Hirschfeld, M.; Dudgeon, C.; Sheaves, M.; Barnett, A.; MacNeil, A. Barriers in a Sea of Elasmobranchs: From Fishing for Populations to Testing Hypotheses in Population Genetics. Glob. Ecol. Biogeogr. 2021, 30, 2147–2163. [Google Scholar] [CrossRef]
- Chabot, C.L. Microsatellite Loci Confirm a Lack of Population Connectivity among Globally Distributed Populations of the Tope Shark Galeorhinus galeus (Triakidae): Population Connectivity of Galeorhinus galeus. J. Fish. Biol. 2015, 87, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Daly-Engel, T.S.; Grubbs, R.D.; Holland, K.N.; Toonen, R.J.; Bowen, B.W. Assessment of Multiple Paternity in Single Litters from Three Species of Carcharhinid Sharks in Hawaii. Environ. Biol. Fishes 2006, 76, 419–424. [Google Scholar] [CrossRef]
- Hoekert, W.E.J.; Neufeglise, H.; Schouten, A.D.; Menken, S.B.J. Multiple Paternity and Female-Biased Mutation at a Microsatellite Locus in the Olive Ridley Sea Turtle (Lepidochelys Olivacea). Heredity 2002, 89, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Newcomer, S.D.; Zeh, J.A.; Zeh, D.W. Genetic Benefits Enhance the Reproductive Success of Polyandrous Females. Proc. Natl. Acad. Sci. USA 1999, 96, 10236–10241. [Google Scholar] [CrossRef]
- Chapman, D.D.; Prodöhl, P.A.; Gelsleichter, J.; Manire, C.A.; Shivji, M.S. Predominance of Genetic Monogamy by Females in a Hammerhead Shark, Sphyrna Tiburo: Implications for Shark Conservation. Mol. Ecol. 2004, 13, 1965–1974. [Google Scholar] [CrossRef]
- Ovenden, J.R.; Berry, O.; Welch, D.J.; Buckworth, R.C.; Dichmont, C.M. Ocean’s Eleven: A Critical Evaluation of the Role of Population, Evolutionary and Molecular Genetics in the Management of Wild Fisheries. Fish Fish. 2015, 16, 125–159. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Burrows, M.T.; Brown, C.J.; García Molinos, J.; Halpern, B.S.; Hoegh-Guldberg, O.; Kappel, C.V.; Moore, P.J.; Richardson, A.J.; Schoeman, D.S. Responses of Marine Organisms to Climate Change across Oceans. Front. Mar. Sci. 2016, 62. [Google Scholar] [CrossRef] [Green Version]
- Boyce, D.G.; Tittensor, D.P.; Garilao, C.; Henson, S.; Kaschner, K.; Kesner-Reyes, K.; Pigot, A.; Reyes Jr, R.B.; Reygondeau, G.; Schleit, K.E. A Climate Risk Index for Marine Life. Nat. Clim. Chang. 2022, 12, 854–862. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; d’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E. A Global Map of Human Impact on Marine Ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, B.S.; Frazier, M.; Potapenko, J.; Casey, K.S.; Koenig, K.; Longo, C.; Lowndes, J.S.; Rockwood, R.C.; Selig, E.R.; Selkoe, K.A. Spatial and Temporal Changes in Cumulative Human Impacts on the World’s Ocean. Nat. Commun. 2015, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, J.-P.; Jones, G.P.; Munday, P.L. Extinction Risk in Endemic Marine Fishes. Conserv. Biol. 2011, 25, 1053–1055. [Google Scholar] [CrossRef]
- O’Hara, C.C.; Frazier, M.; Halpern, B.S. At-Risk Marine Biodiversity Faces Extensive, Expanding, and Intensifying Human Impacts. Science 2021, 372, 84–87. [Google Scholar] [CrossRef]
- McCauley, D.J.; Pinsky, M.L.; Palumbi, S.R.; Estes, J.A.; Joyce, F.H.; Warner, R.R. Marine Defaunation: Animal Loss in the Global Ocean. Science 2015, 347, 1255641. [Google Scholar] [CrossRef] [Green Version]
- Lotze, H.K. Marine Biodiversity Conservation. Curr. Biol. 2021, 31, R1190–R1195. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adachi, A.M.C.L.; Roque, P.C.G.; Hazin, F.H.V.; Vianna, M.; Rotundo, M.M.; Oliveira, C.; Foresti, F.; Cruz, V.P. Genetic Population Structure and Diversity of the Whitetail Dogfish Squalus albicaudus (Chondrichthyes, Squaliformes) along the Brazilian Coast as Identified by SNP Markers. Fishes 2023, 8, 373. https://doi.org/10.3390/fishes8070373
Adachi AMCL, Roque PCG, Hazin FHV, Vianna M, Rotundo MM, Oliveira C, Foresti F, Cruz VP. Genetic Population Structure and Diversity of the Whitetail Dogfish Squalus albicaudus (Chondrichthyes, Squaliformes) along the Brazilian Coast as Identified by SNP Markers. Fishes. 2023; 8(7):373. https://doi.org/10.3390/fishes8070373
Chicago/Turabian StyleAdachi, Aisni M. C. L., Pollyana C. G. Roque, Fabio H. V. Hazin, Marcelo Vianna, Matheus M. Rotundo, Claudio Oliveira, Fausto Foresti, and Vanessa P. Cruz. 2023. "Genetic Population Structure and Diversity of the Whitetail Dogfish Squalus albicaudus (Chondrichthyes, Squaliformes) along the Brazilian Coast as Identified by SNP Markers" Fishes 8, no. 7: 373. https://doi.org/10.3390/fishes8070373
APA StyleAdachi, A. M. C. L., Roque, P. C. G., Hazin, F. H. V., Vianna, M., Rotundo, M. M., Oliveira, C., Foresti, F., & Cruz, V. P. (2023). Genetic Population Structure and Diversity of the Whitetail Dogfish Squalus albicaudus (Chondrichthyes, Squaliformes) along the Brazilian Coast as Identified by SNP Markers. Fishes, 8(7), 373. https://doi.org/10.3390/fishes8070373