


Year-Round Spawning, Filial Cannibalism, and Embryonic and Larval Development of the Coral Reef Fish Orchid Dottyback, Pseudochromis fridmani


Abstract
:1. Introduction
2. Materials and Methods
2.1. Brood Fish and Pair Formation
2.2. Reproductive Behaviour, Spawning, and Filial Cannibalism
2.3. Embryonic Development
2.4. Larval Development
2.4.1. Larval Rearing
2.4.2. Larvae Sampling
3. Results
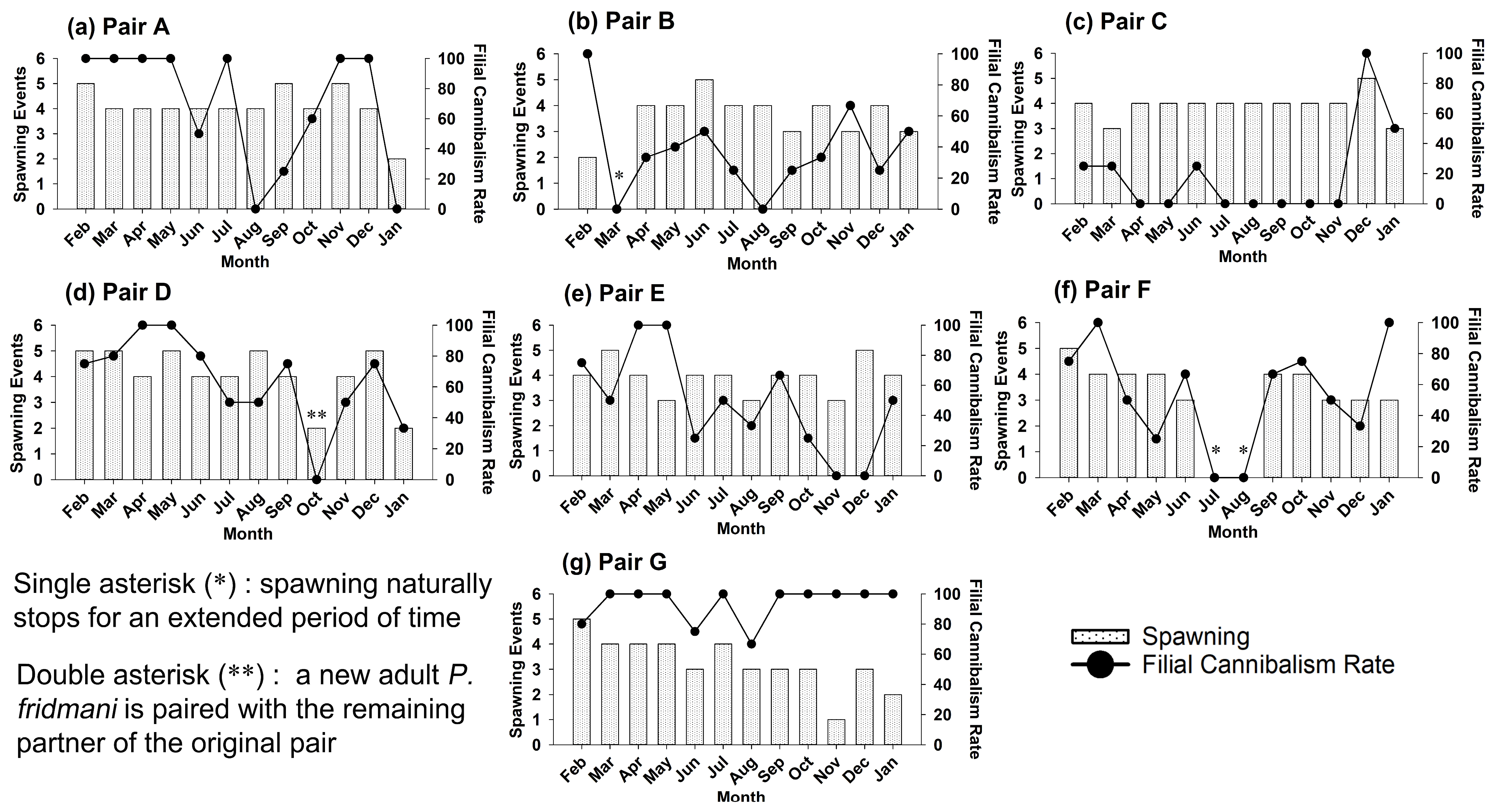
3.1. Reproductive Behaviour of Brood Pairs
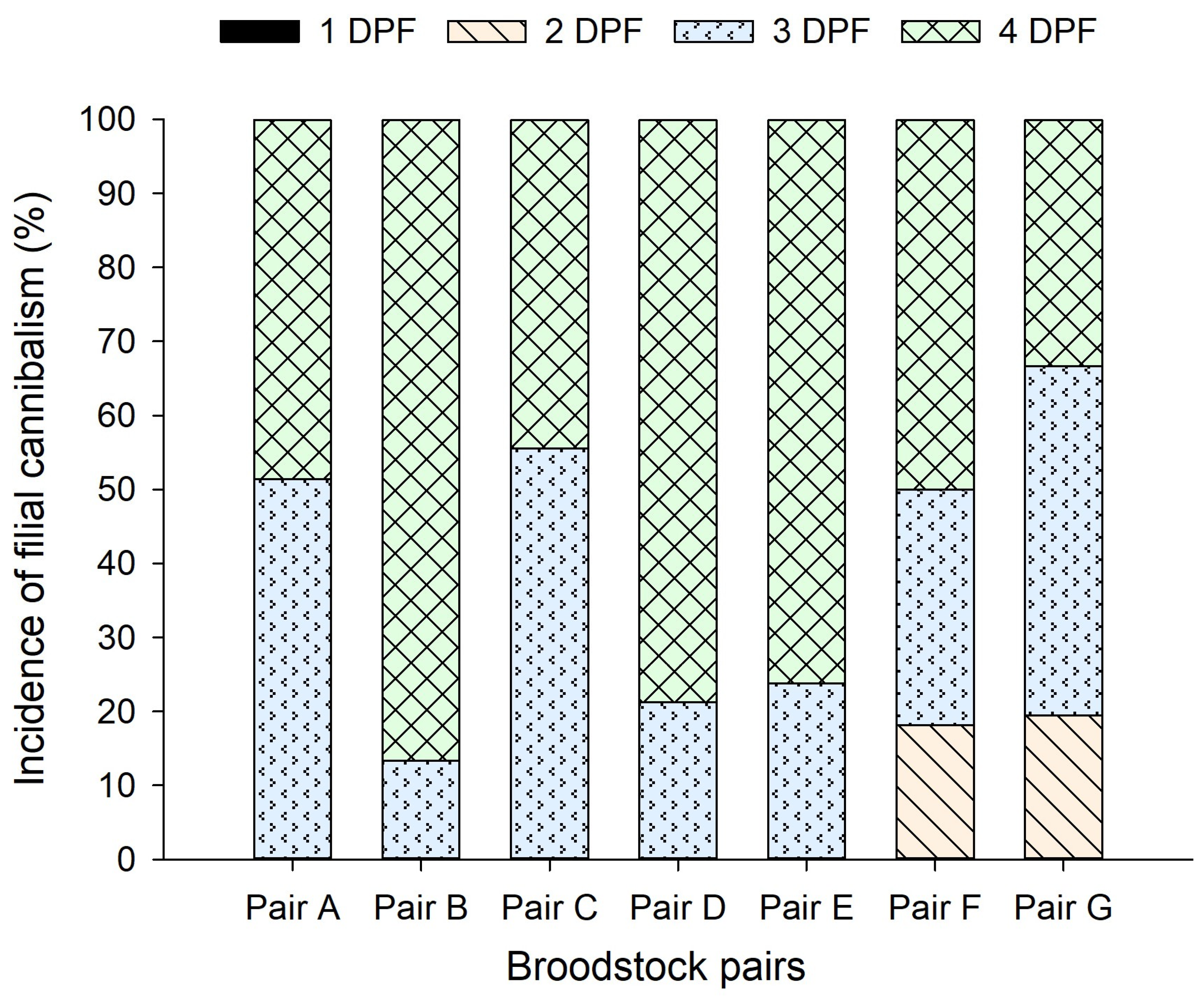
3.2. Captive Spawning and Filial Cannibalism
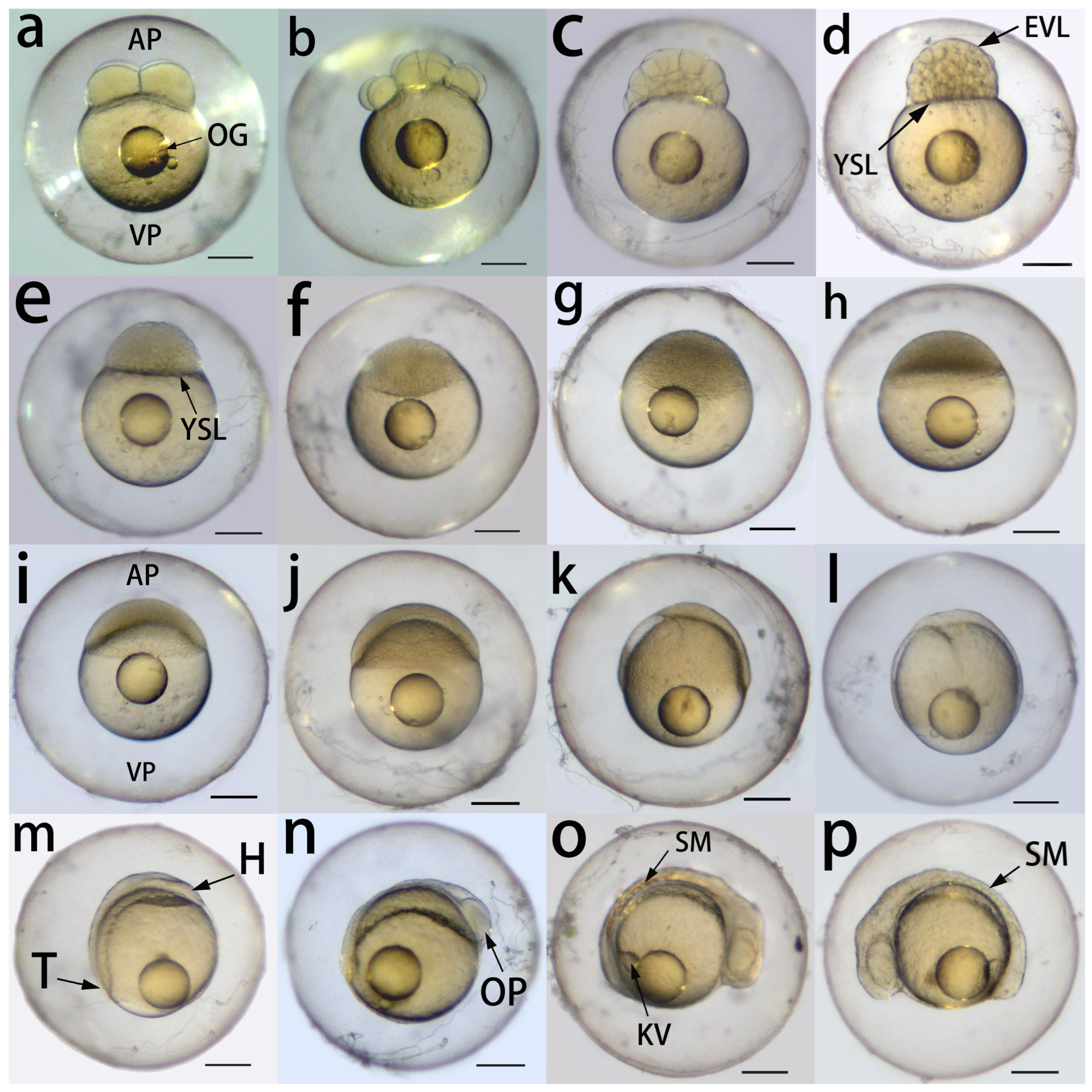
3.3. Embryonic Development
3.3.1. Cleavage Period (2- to 64-Cell Stage)
3.3.2. Blastula Period (128-Cell Stage to 30% Epiboly)
3.3.3. Gastrula Period (50% Epiboly to Bud Stage)
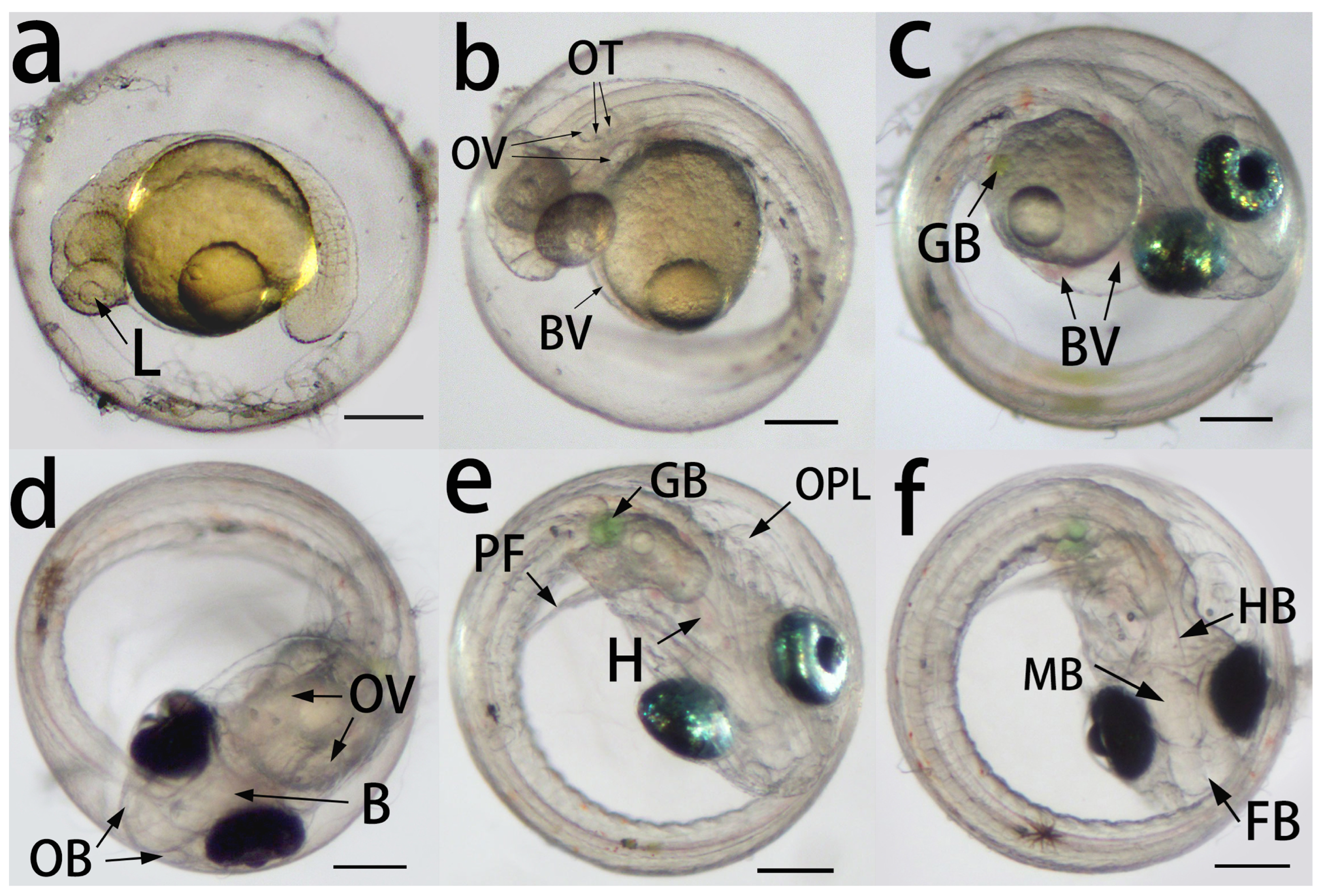
3.3.4. Segmentation and Pharyngula Period

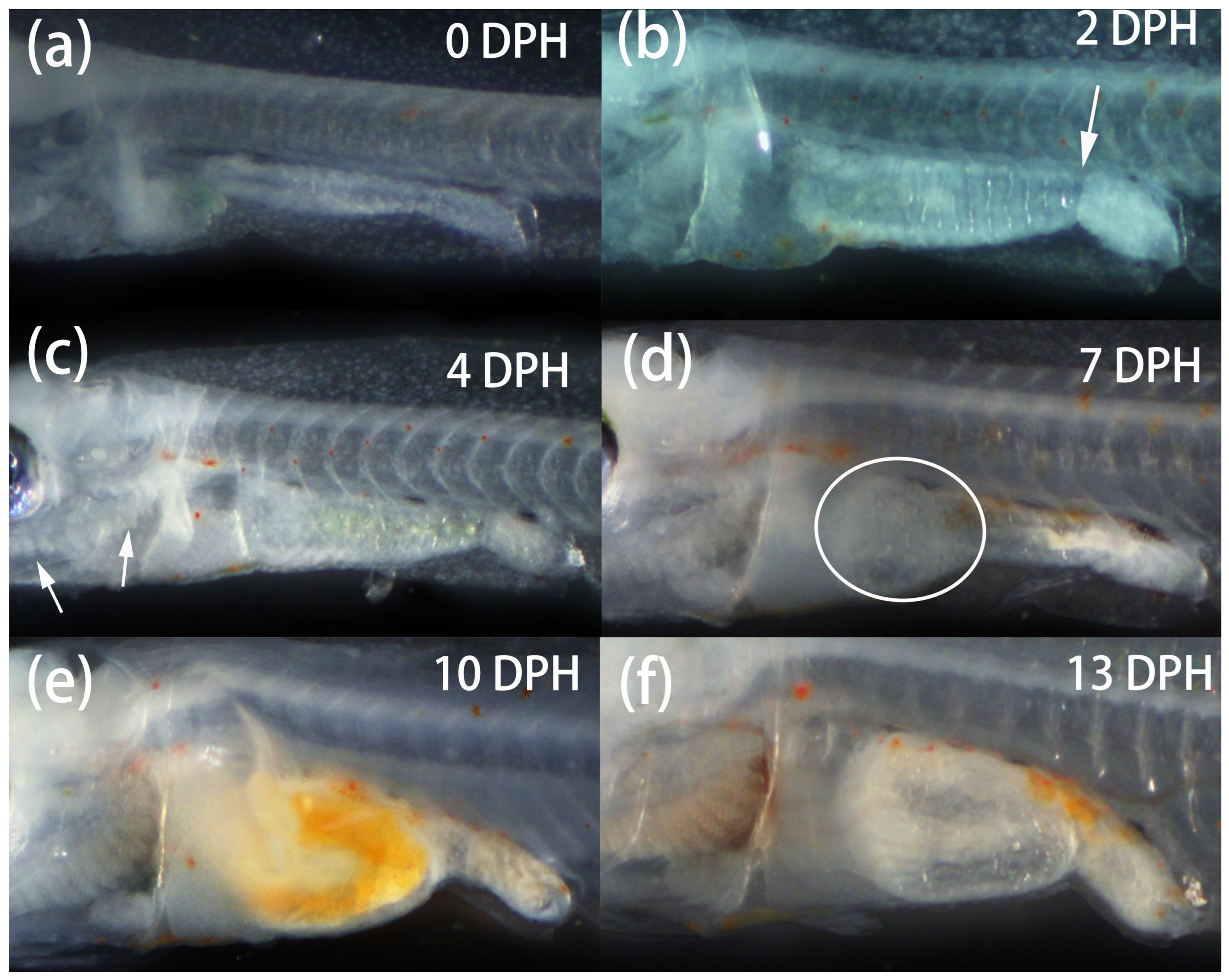
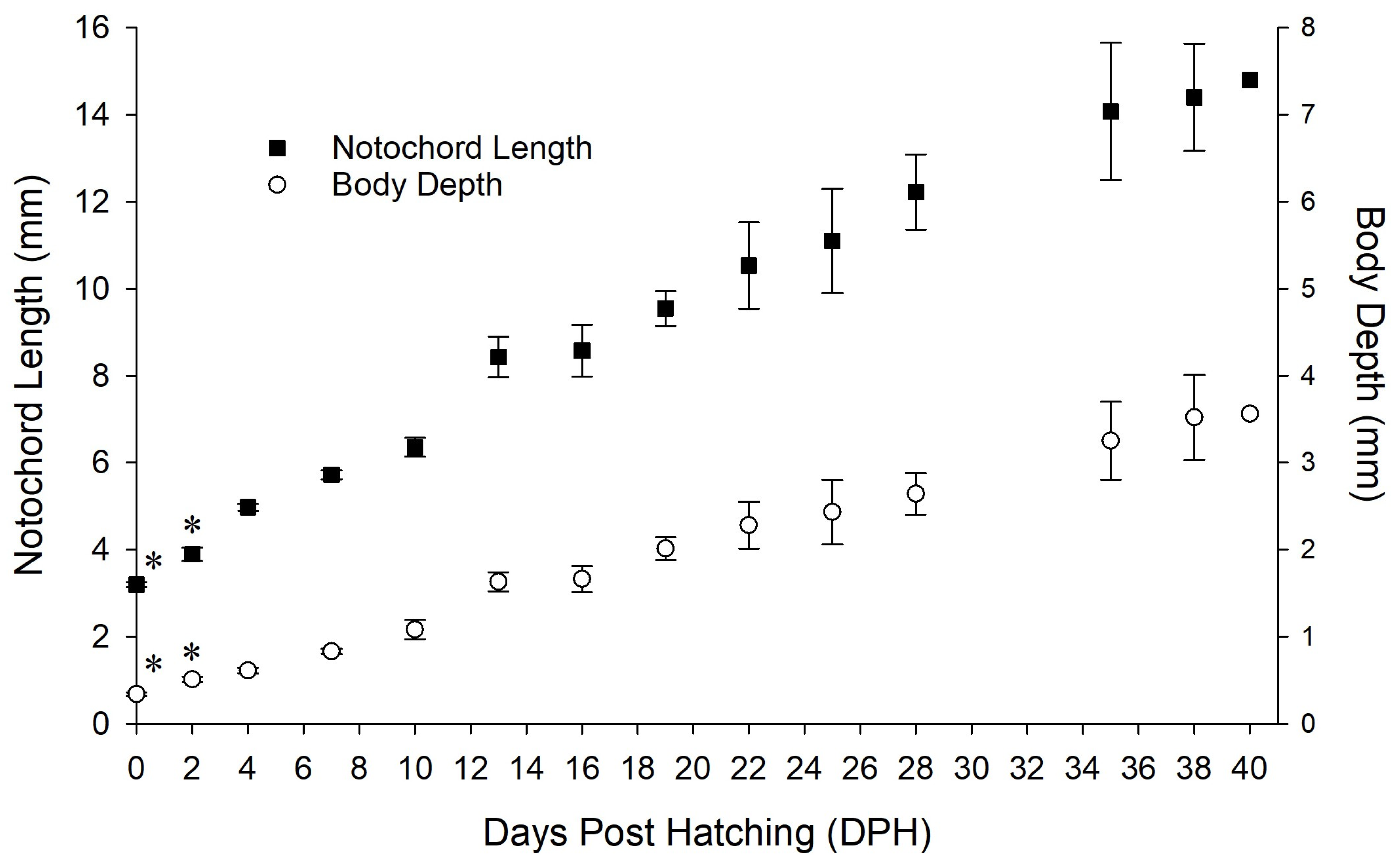
3.4. Larval Development and Associated Behavioural Observations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Froese, R.; Pauly, D. Family Pseudochromidae—Dottybacks. Available online: https://www.fishbase.se/summary/FamilySummary.php?ID=291 (accessed on 6 January 2023).
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Family PSEUDOCHROMIDAE (289)—Dottybacks. In Fishes of the World; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 339–340. [Google Scholar]
- Gill, A.C.; Allen, G.R.; Erdmann, M.V. Pseudochromis stellatus, a New Species of Dottyback from Indonesia (Teleostei: Pseudochromidae). Zootaxa 2017, 4338, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Gill, A.C.; Allen, G.R.; Erdmann, M. Two New Dottyback Species of the Genus Pseudochromis from Southern Indonesia (Teleostei: Pseudochromidae). Zootaxa 2012, 60, 53–60. [Google Scholar] [CrossRef]
- Lieske, E.; Myers, R. Dottybacks. In Coral Reef Fishes: Indo-Pacific & Caribbean; HarperCollins Publishers: London, UK, 1993; pp. 31–33. ISBN 0002199742. [Google Scholar]
- Kuiter, R.H. Dottybacks—Pseudochromidae. In Guide to Sea Fishes of Australia: A Comprehensive Reference for Divers and Fishermen; New Holland Publisher (Australia) Pty Ltd.: Sydney, Australia, 2016; pp. 124–127. [Google Scholar]
- Randall, J.E.; Allen, G.R.; Steene, R.C. Dottybacks and Eel Blennies. In Fishes of the Great Barrier Reef and Coral Sea; University of Hawaii Press: Honolulu, HI, USA, 1997; pp. 124–132. [Google Scholar]
- Lubbock, R. Fishes of the Family Pseudochromidae (Perciformes) in the Northwest Indian Ocean and Red Sea. J. Zool. 1975, 176, 115–157. [Google Scholar] [CrossRef]
- Rhyne, A.L.; Tlusty, M.F.; Schofield, P.J.; Kaufman, L.; Morris, J.A.; Bruckner, A.W. Revealing the Appetite of the Marine Aquarium Fish Trade: The Volume and Biodiversity of Fish Imported into the United States. PLoS ONE 2012, 7, e35808. [Google Scholar] [CrossRef] [PubMed]
- Wittenrich, M.L. The Complete Illustrated Breeder’s Guide to Marine Aquarium Fishes; T.F.H. Publications: Neptune City, NJ, USA, 2007. [Google Scholar]
- Field, R.; Field, M. Dottybacks. In Reef Fish of the Red Sea; Routledge Ltd.: New York, NY, USA, 2016; pp. 76–78. [Google Scholar]
- Taquet, M.; Diringer, A. Dottybacks. In Fishes of the Indian Ocean and Red Sea; Quae: Versailles, France, 2012; pp. 233–236. ISBN 9782759218899. [Google Scholar]
- Jiang, J.; Nuez-Ortin, W.; Angell, A.; Zeng, C.; de Nys, R.; Vucko, M.J. Enhancing the Colouration of the Marine Ornamental Fish Pseudochromis fridmani Using Natural and Synthetic Sources of Astaxanthin. Algal Res. 2019, 42, 101596. [Google Scholar] [CrossRef]
- LiveAquaria ORA® Captive-Bred Orchid Dottyback. Available online: https://www.liveaquaria.com/product/prod_display.cfm?c=15+38+3465&pcatid=3465 (accessed on 10 April 2023).
- Palmtag, M.R. The Marine Ornamental Species Trade. In Marine Ornamental Species Aquaculture; Calado, R., Olivotto, I., Oliver, M.P., Holt, G.J., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2017; pp. 3–14. [Google Scholar]
- Wabnitz, C.; Taylor, M.; Green, E.; Razak, T. From Ocean to Aquarium: The Global Trade in Marine Ornamental Species; UNEP–WCMC: Cambridge, UK, 2003. [Google Scholar]
- Olivotto, I.; Chemello, G.; Vargas, A.; Randazzo, B.; Piccinetti, C.C.; Carnevali, O. Marine Ornamental Species Culture: From the Past to “Finding Dory”. Gen. Comp. Endocrinol. 2017, 245, 116–121. [Google Scholar] [CrossRef]
- Degidio, J.M.L.A.; Yanong, R.P.E.; Watson, C.A.; Ohs, C.L.; Cassiano, E.J.; Barden, K. Spawning, Embryology, and Larval Development of the Milletseed Butterflyfish Chaetodon miliaris in the Laboratory. N. Am. J. Aquac. 2017, 79, 205–215. [Google Scholar] [CrossRef]
- Majoris, J.E.; Francisco, F.A.; Atema, J.; Buston, P.M. Reproduction, Early Development, and Larval Rearing Strategies for Two Sponge-Dwelling Neon Gobies, Elacatinus lori and E. colini. Aquaculture 2018, 483, 286–295. [Google Scholar] [CrossRef]
- Mendoncą, R.C.; Chen, J.Y.; Zeng, C.; Tsuzuki, M.Y. Embryonic and Early Larval Development of Two Marine Angelfish, Centropyge bicolor and Centropyge bispinosa. Zygote 2020, 28, 196–202. [Google Scholar] [CrossRef]
- Moorhead, J.A. Broodstock Management, Development of Rearing Systems, and Feeding Regimes for Larvae of the Forktail Blenny, Meiacanthus atrodorsalis. Ph.D. Thesis, James Cook University, Townsville, Australia, 2017. [Google Scholar]
- Mies, M.; Güth, A.Z.; Scozzafave, M.S.; Sumida, P.Y.G. Spawning Behaviour and Activity in Seven Species of Ornamental Dottybacks. J. Zoo Aquar. Res. 2014, 2, 117–122. [Google Scholar]
- Shei, M.; Mies, M.; Olivotto, I. Other Demersal Spawners and Mouthbrooders. In Marine Ornamental Species Aquaculture; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 223–250. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C.; Cobcroft, J.M. Digestive System Ontogeny and the Effects of Weaning Time on Larval Survival, Growth and Pigmentation Development of Orchid Dottyback Pseudochromis fridmani. Aquaculture 2022, 549, 737737. [Google Scholar] [CrossRef]
- Wittenrich, M.L.; Turingan, R.G. Linking Functional Morphology and Feeding Performance in Larvae of Two Coral-Reef Fishes. Environ. Biol. Fishes 2011, 92, 295–312. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C. The Effects of Live Prey and Greenwater on the Early Larval Rearing of Orchid Dottyback Pseudochromis fridmani. Aquaculture 2021, 543, 737008. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C. Transition to Artemia Feeding Phase for Orchid Dottyback Pseudochromis fridmani Larvae: Establishing Suitable Prey Shift Time and Strategy. Aquaculture 2021, 545, 737180. [Google Scholar] [CrossRef]
- Moe, M.A. Breeding the Orchid Dottyback, Pseudochromis fridmani: An Aquarist’s Journal; Green Turtle Publications: Plantation, FL, USA, 1997; ISBN 9780939960095. [Google Scholar]
- Madhu, K.; Madhu, R.; Retheesh, T. Spawning, Embryonic Development and Larval Culture of Redhead Dottyback Pseudochromis dilectus Lubbock, 1976 under Captivity. Aquaculture 2016, 459, 73–83. [Google Scholar] [CrossRef]
- Madhu, K.; Madhu, R. Captive Spawning and Embryonic Development of Marine Ornamental Purple Firefish Nemateleotris decora (Randall & Allen, 1973). Aquaculture 2014, 424–425, 1–9. [Google Scholar] [CrossRef]
- Leu, M.Y.; Tai, K.; Meng, P.; Tang, C.; Wang, P. Embryonic, Larval and Juvenile Development of the Long Fin Batfish, Platax teira (Forsskål,1775) under Controlled Conditions with Special Regard to Mitigate Cannibalism for Larviculture. Aquaculture 2018, 493, 204–213. [Google Scholar] [CrossRef]
- Zeng, C.; Shao, L.; Ricketts, A.; Moorhead, J. The Importance of Copepods as Live Feed for Larval Rearing of the Green Mandarin Fish Synchiropus splendidus. Aquaculture 2018, 491, 65–71. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Olivotto, I.; Rollo, A.; Sulpizio, R.; Avella, M.; Tosti, L.; Carnevali, O. Breeding and Rearing the Sunrise Dottyback Pseudochromis flavivertex: The Importance of Live Prey Enrichment during Larval Development. Aquaculture 2006, 255, 480–487. [Google Scholar] [CrossRef]
- Olivotto, I.; Holt, S.A.; Carnevali, O.; Holt, G.J. Spawning, Early Development, and First Feeding in the Lemonpeel Angelfish Centropyge Flavissimus. Aquaculture 2006, 253, 270–278. [Google Scholar] [CrossRef]
- Baensch, F.U.; Tamaru, C.S. Spawning and Development of Larvae and Juveniles of the Rare Blue Mauritius Angelfish, Centropyge debelius (1988), in the Hatchery. J. World Aquac. Soc. 2009, 40, 425–439. [Google Scholar] [CrossRef]
- Manica, A. Parental Fish Change Their Cannibalistic Behaviour in Response to the Cost-to-Benefit Ratio of Parental Care. Anim. Behav. 2004, 67, 1015–1021. [Google Scholar] [CrossRef]
- Mehlis, M.; Bakker, T.C.M.; Frommen, J.G. Nutritional Benefits of Filial Cannibalism in Three-Spined Sticklebacks (Gasterosteus aculeatus). Naturwissenschaften 2009, 96, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Manica, A. Filial Cannibalism in Teleost Fish. Biol. Rev. 2002, 77, 261–277. [Google Scholar] [CrossRef]
- Rohwer, S. Parent Cannibalism of Offspring and Egg Raiding as a Courtship Strategy. Am. Nat. 1978, 112, 429–440. [Google Scholar] [CrossRef]
- Petersen, C.W.; Marchetti, K. Filial Cannibalism in the Cortez Damselfish Stegastes rectifraenum. Evolution 1989, 43, 158–168. [Google Scholar] [CrossRef]
- Vallon, M.; Heubel, K.U. Old but Gold: Males Preferentially Cannibalize Young Eggs. Behav. Ecol. Sociobiol. 2016, 70, 569–573. [Google Scholar] [CrossRef]
- Payne, A.G.; Smith, C.; Campbell, A.C. Filial Cannibalism Improves Survival and Development of Beaugregory Damselfish Embryos. Proc. R. Soc. B Biol. Sci. 2002, 269, 2095–2102. [Google Scholar] [CrossRef]
- Mrowka, W. Filial Cannibalism and Reproductive Success in the Maternal Mouthbrooding Cichlid Fish Pseudocrenilabrus multicolor. Behav. Ecol. Sociobiol. 1987, 21, 257–265. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Tateishi, T.; Terada, R.; Soyano, K.; Takegaki, T. Filial Cannibalism by Male Fish as an Infanticide to Restart Courtship by Self-Regulating Androgen Levels. Curr. Biol. 2018, 28, 2831–2836. [Google Scholar] [CrossRef] [PubMed]
- Stearns, D.E.; Holt, J.G.; Forward Jr, R.B.; Pickering, P.L. Ontogeny of Phototactic Behavior in Red Drum Larvae (Sciaenidae: Sciaenops ocellatus). Mar. Ecol. Prog. Ser. 1994, 104, 1–11. [Google Scholar] [CrossRef]
- Hilder, P.E.; Battaglene, S.C.; Hart, N.S.; Collin, S.P.; Cobcroft, J.M. Retinal Adaptations of Southern Bluefin Tuna Larvae: Implications for Culture. Aquaculture 2019, 507, 222–232. [Google Scholar] [CrossRef]
- Papadakis, I.E.; Kentouri, M.; Divanach, P.; Mylonas, C.C. Ontogeny of the Eye of Meagre (Argyrosomus regius) from Hatching to Juvenile and Implications to Commercial Larval Rearing. Aquaculture 2018, 484, 32–43. [Google Scholar] [CrossRef]
- Ina, Y.; Sakakura, Y.; Tanaka, Y.; Yamada, T.; Kumon, K.; Eba, T.; Hashimoto, H.; Konishi, J.; Takashi, T.; Gen, K. Development of Phototaxis in the Early Life Stages of Pacific Bluefin Tuna Thunnus orientalis. Fish. Sci. 2017, 83, 537–542. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Broodstock Pair | Monthly Spawning Events | Monthly Filial Cannibalism Rate (%) |
|---|---|---|
| Pair A | 4.1 ± 0.8 | 70 ± 41 |
| Pair B | 3.3 ± 1.3 | 37 ± 28 |
| Pair C | 3.9 ± 0.5 | 19 ± 30 |
| Pair D | 4.1 ± 1.1 | 64 ± 29 |
| Pair E | 3.9 ± 0.7 | 48 ± 34 |
| Pair F | 3.1 ± 1.6 | 54 ± 34 |
| Pair G | 3.3 ± 1.1 | 94 ± 12 |
| All Pairs Combined | 3.7 ± 1.1 | 55 ± 37 |
| Developmental Stage | Time | Main Characteristics | Corresponding Figure |
|---|---|---|---|
| Cleavage | |||
| 4-cell stage | 1 h | 2 × 2 array of blastomeres | Figure 3a |
| 16-cell stage | 1 h 30 min | 4 × 4 array of blastomeres | Figure 3b |
| 64-cell stage | 2 h | 3 layers of blastomeres | Figure 3c |
| Blastula | |||
| Between 128- and 256-cell stage | 2 h 30 min | 5–7 layers of blastomeres; enveloping layer forms | – |
| 512-cell stage | 3 h | 8–9 layers of blastomeres; yolk syncytial layer forms | Figure 3d |
| >1-k-cell stage | 3 h 30 min | Blastomeres become smaller | – |
| High stage | 4 h | Blastodisc starts to flatten | Figure 3e |
| Oblong stage | 5 h | Ellipsoidal shape; flattened blastodisc | Figure 3f |
| Between oblong and sphere stage | 7 h | Less distinguishable boundary between blastodisc and yolk | Figure 3g |
| Sphere stage | 8 h | Spherical shape | Figure 3h |
| Dome stage | 9 h | Forms a dome-like structure; beginning of epiboly | Figure 3i |
| 30% epiboly | 10 h | Blastoderm covers 30% of the yolk surface | Figure 3j |
| Gastrula | |||
| 60% epiboly | 12 h | Blastoderm covers 60% of the yolk surface | Figure 3k |
| 90% epiboly | 14 h | Blastoderm covers 90% of the yolk surface | Figure 3l |
| Bud | 16 h | Blastoderm completely covers the yolk; prospective head and tail | Figure 3m |
| Segmentation and Pharyngula | |||
| Optic primordium | 18 h | Optic primordium appears in the prospective head region | Figure 3n |
| 6-somite | 20 h | Kupffer’s vesicle appears near the tail region; | Figure 3o |
| 10-somite | 22 h | 10 somites in the embryo body | Figure 3p |
| Lens | 24 h | Rudiments of eyes and lens | Figure 3a |
| Heartbeat | 48 h | Visible heartbeat, blood circulation, and otic vesicles with two otoliths; myotomes and pigmentation develop on the body; head and tail resemble that of larvae | Figure 3b |
| Pigmented eyes | 72 h | Pigmented eyes, olfactory bulbs, and gall bladder appear; stronger heartbeats | Figure 4c,d |
| Brain | 96 h | Reflective eyes, pectoral fin buds, and operculum appear; distinguishable forebrain, midbrain, and hindbrain; frequent movements of the eyes and embryos | Figure 4e,f |
| Species | Incubation Temperature | Beginning of Epiboly | 75% Epiboly | Blastoderm Closure | Head Bud | 5-Somite | Metallic Eyes | Frequent Movements of Embryos | References |
|---|---|---|---|---|---|---|---|---|---|
| P. flavivertex | 27 °C | - | - | - | - | - | 96 h | 96 h | [34] |
| P. dilectus | 27 ± 1 °C | 7 h | 7 h 30 min | 8 h | 10 h | 13–18 h | 96 h | 96 h | [29] |
| P. fridmani | 27 ± 1 °C | 9 h | 12–14 h | 14–16 h | 16 h | 18–20 h | 72 h | 96 h | Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.Y.; Zeng, C.; Cobcroft, J.M. Year-Round Spawning, Filial Cannibalism, and Embryonic and Larval Development of the Coral Reef Fish Orchid Dottyback, Pseudochromis fridmani. Fishes 2023, 8, 451. https://doi.org/10.3390/fishes8090451
Chen JY, Zeng C, Cobcroft JM. Year-Round Spawning, Filial Cannibalism, and Embryonic and Larval Development of the Coral Reef Fish Orchid Dottyback, Pseudochromis fridmani. Fishes. 2023; 8(9):451. https://doi.org/10.3390/fishes8090451
Chicago/Turabian StyleChen, Jun Yu, Chaoshu Zeng, and Jennifer M. Cobcroft. 2023. "Year-Round Spawning, Filial Cannibalism, and Embryonic and Larval Development of the Coral Reef Fish Orchid Dottyback, Pseudochromis fridmani" Fishes 8, no. 9: 451. https://doi.org/10.3390/fishes8090451
APA StyleChen, J. Y., Zeng, C., & Cobcroft, J. M. (2023). Year-Round Spawning, Filial Cannibalism, and Embryonic and Larval Development of the Coral Reef Fish Orchid Dottyback, Pseudochromis fridmani. Fishes, 8(9), 451. https://doi.org/10.3390/fishes8090451