
Application of Fixed-Length Ultrasonic Interferometry to Determine the Kinetics of Light-/Heat-Induced Damage to Biological Membranes and Protein Complexes

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Setup
- Acceleration of the operation of the controller that tunes the DDS generator: an 8-bit ATMega family (Atmel, Microchip Technology Inc., Chandler, AZ, USA) with a clock frequency of 16 MHz was replaced with a 32-bit SAM3 controller (Atmel, Microchip Technology Inc., Chandler, AZ, USA), that is replacing the Arduino Uno controller with the Arduino DUE controller.
- Transition from a circuit with one oscillator and one oscilloscope serving two channels and a switch to a circuit with one 125 MHz master oscillator and two DDS generators.
- Replacement of the digital oscilloscope in the channel of the experimental cell with a high-speed recorder with 32 MB of RAM and a USB3.0 interface. The recorder is started by an external trigger. The trigger signal is generated by the Arduino DUE.
- The measurement procedure in the channel of the experimental cell was changed. The Arduino DUE controller independently forms a grid of 15 frequencies and the 16th interval with the generator turned off. The entire frequency grid is automatically fed to the input of the experimental cell without the participation of a personal computer. The measurement result (in the form of a continuous recording with a sampling frequency of 50 MHz and eight million samples long) is transferred to a computer and analyzed. Thus, the procedure for measuring the characteristics of one resonant peak takes 160 ms, rather than 4 s as in the previous version.
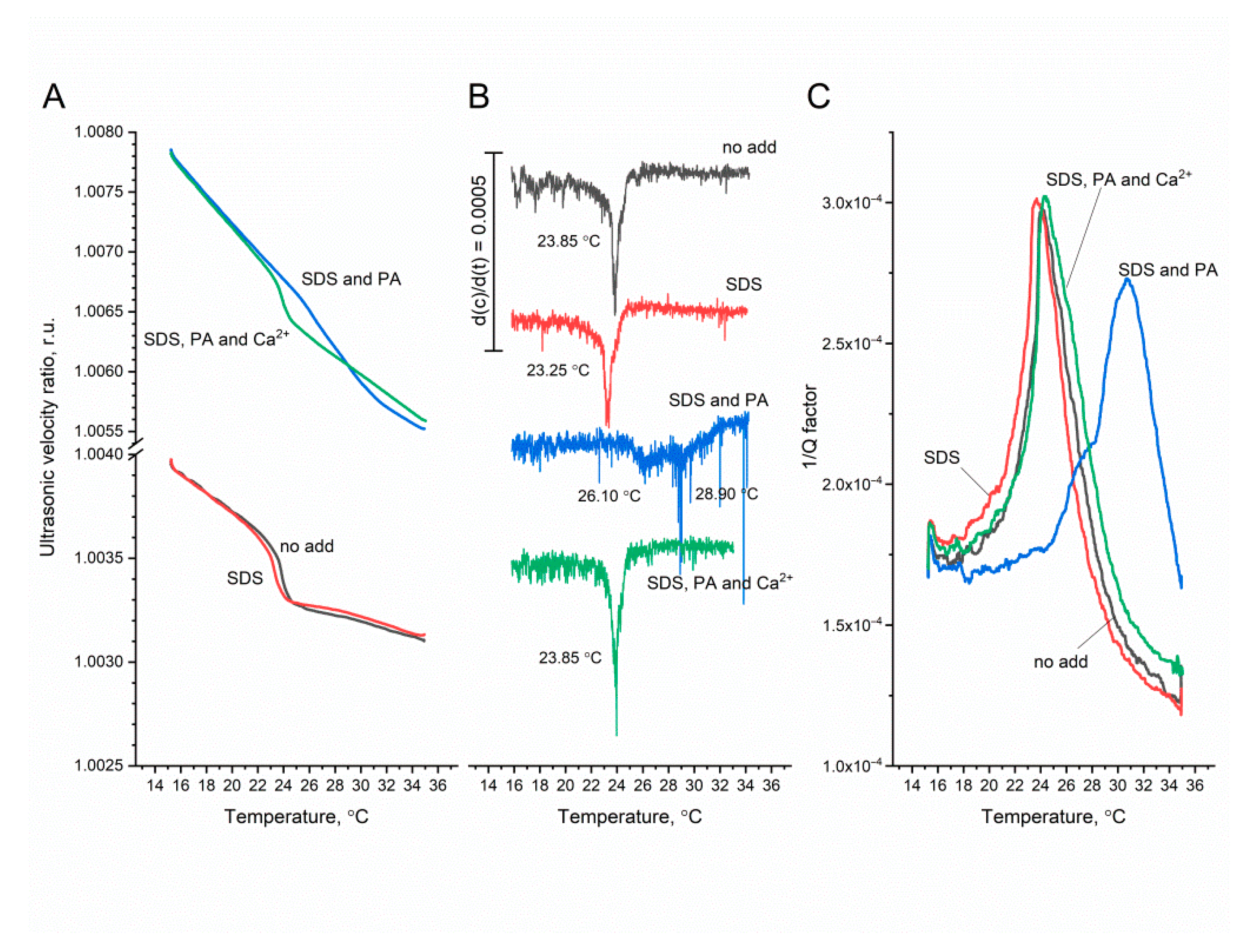
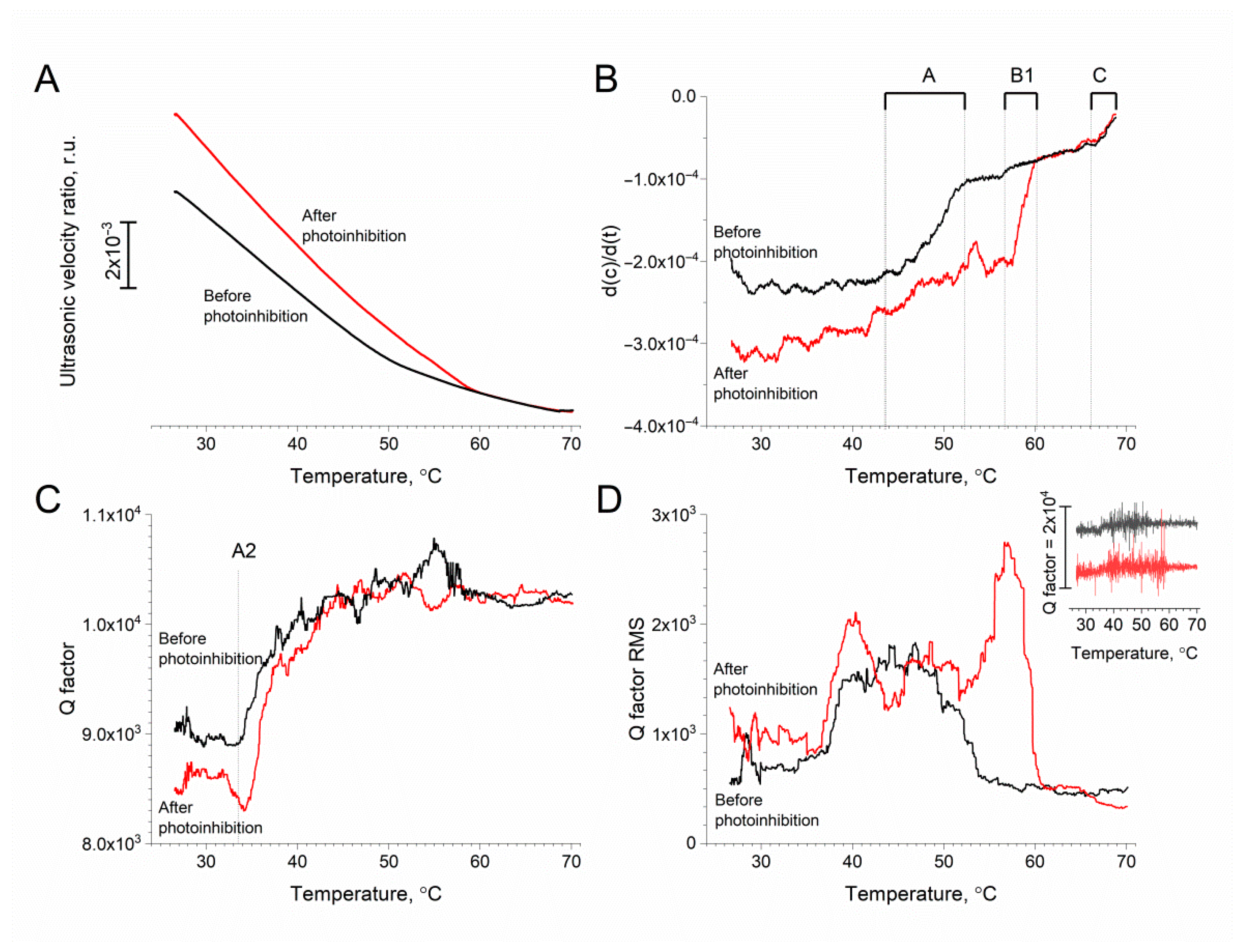
3.2. Measurements
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Losada-Pérez, P.; Jiménez-Monroy, K.; Van Grinsven, B.; Leys, J.; Janssens, S.; Peeters, M.; Glorieux, C.; Thoen, J.; Haenen, K.; De Ceuninck, W. Phase transitions in lipid vesicles detected by a complementary set of methods: Heat-transfer measurements, adiabatic scanning calorimetry, and dissipation-mode quartz crystal microbalance. Phys. Status Solidi 2014, 211, 1377–1388. [Google Scholar] [CrossRef]
- Hazel, J.R.; McKinley, S.J.; Gerrits, M.F. Thermal acclimation of phase behavior in plasma membrane lipids of rainbow trout hepatocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 275, R861–R869. [Google Scholar] [CrossRef] [PubMed]
- Hazel, J.R.; Williams, E.E. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef]
- Melchior, D.L.; Steim, J.M. Lipid-Associated Thermal Events in Biomembranes. In Progress in Surface and Membrane Science; Cadenhead, D.A., Danielli, J.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1979; Volume 13, pp. 211–296. [Google Scholar]
- Sandermann, H. Regulation of membrane enzymes by lipids. Biochim. Biophys. Acta (BBA)—Rev. Biomembr. 1978, 515, 209–237. [Google Scholar] [CrossRef]
- Leonard, J.; Lee, C.; Apperson, A.; Dills, S.; Saier, M. The role of membranes in the transport of small molecules. In Organization of Prokaryotic Cell Membranes; CRC Press: Boca Raton, FL, USA, 2018; pp. 1–52. [Google Scholar]
- Bondar, A.-N.; Lemieux, M.J. Reactions at Biomembrane Interfaces. Chem. Rev. 2019, 119, 6162–6183. [Google Scholar] [CrossRef]
- Finkelstein, A.V.; Ptitsyn, O. Protein Physics: A Course of Lectures; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Mohammadi, P.; Aranko, A.S.; Lemetti, L.; Cenev, Z.; Zhou, Q.; Virtanen, S.; Landowski, C.P.; Penttilä, M.; Fischer, W.J.; Wagermaier, W.; et al. Phase transitions as intermediate steps in the formation of molecularly engineered protein fibers. Commun. Biol. 2018, 1, 86. [Google Scholar] [CrossRef]
- Shin, Y.; Brangwynne, C.P. Liquid phase condensation in cell physiology and disease. Science 2017, 357, eaaf4382. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, S.; Huang, Y.; He, X.; Cui, H.; Zhu, X.; Zheng, Y. Phase Transition of Spindle-Associated Protein Regulate Spindle Apparatus Assembly. Cell 2015, 163, 108–122. [Google Scholar] [CrossRef]
- Zhang, H.; Elbaum-Garfinkle, S.; Langdon, E.M.; Taylor, N.; Occhipinti, P.; Bridges, A.A.; Brangwynne, C.P.; Gladfelter, A.S. RNA Controls PolyQ Protein Phase Transitions. Mol. Cell 2015, 60, 220–230. [Google Scholar] [CrossRef]
- Fersht, A.R. From the first protein structures to our current knowledge of protein folding: Delights and scepticisms. Nat. Rev. Mol. Cell Biol. 2008, 9, 650–654. [Google Scholar] [CrossRef]
- Boeynaems, S.; Alberti, S.; Fawzi, N.L.; Mittag, T.; Polymenidou, M.; Rousseau, F.; Schymkowitz, J.; Shorter, J.; Wolozin, B.; Van Den Bosch, L.; et al. Protein Phase Separation: A New Phase in Cell Biology. Trends Cell Biol. 2018, 28, 420–435. [Google Scholar] [CrossRef] [PubMed]
- Kharakoz, D.P. On the possible physiological role of the “liquid-solid” phase transition in biological membranes. Uspekhi Biol. Biol. Khimii 2001, 41, 333–364. [Google Scholar]
- Neupane, S.; Cordoyiannis, G.; Renner, F.U.; Losada-Pérez, P. Real-time monitoring of interactions between solid-supported lipid vesicle layers and short-and medium-chain length alcohols: Ethanol and 1-pentanol. Biomimetics 2019, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.T.; Kreutzberger, A.J.B.; Lee, J.; Kiessling, V.; Tamm, L.K. The role of cholesterol in membrane fusion. Chem. Phys. Lipids 2016, 199, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Widomska, J.; Mainali, L.; Raguz, M. High Cholesterol/Low Cholesterol: Effects in Biological Membranes: A Review. Cell Biochem. Biochem. Biochem. Biophys. 2017, 75, 369–385. [Google Scholar] [CrossRef]
- Privalov, P.L. Scanning microcalorimeters for studying macromolecules. Pure Appl. Chem. 1980, 52, 479–497. [Google Scholar] [CrossRef]
- Shi, H.; Xiong, L.; Yang, K.-y.; Tang, C.-q.; Kuang, T.-y.; Zhao, N.-m. Protein secondary structure and conformational changes of photosystem II during heat denaturation studied by Fourier transform-infrared spectroscopy. J. Mol. Struct. 1998, 446, 137–147. [Google Scholar] [CrossRef]
- Au-Durowoju, I.B.; Au-Bhandal, K.S.; Au-Hu, J.; Au-Carpick, B.; Au-Kirkitadze, M. Differential Scanning Calorimetry—A Method for Assessing the Thermal Stability and Conformation of Protein Antigen. JoVE 2017, e55262. [Google Scholar] [CrossRef]
- Privalov, P.L. Stability of proteins small globular proteins. Adv. Protein Chem. 1979, 33, 167–241. [Google Scholar]
- Privalov, P.L.; Gill, S.J. Stability of protein structure and hydrophobic interaction. Adv. Protein Chem. 1988, 39, 191–234. [Google Scholar]
- Donovan, J.W. Scanning calorimetry of complex biological structures. Trends Biochem. Sci. 1984, 9, 340–344. [Google Scholar] [CrossRef]
- Nolan, W.G.; Hopkins, H.P.; Kalini, S.A.M. Differential scanning calorimetric investigation of pea chloroplast thylakoids and thylakoid fractions. Arch. Biochem. Biophys. 1992, 297, 19–27. [Google Scholar] [CrossRef]
- Krumova, S.B.; Todinova, S.J.; Dobrikova, A.G.; Taneva, S.G. Differential scanning calorimetry of photosynthetic membranes: Resolving contributions of the major photosynthetic complexes to the sequential thermal transitions. Trends Photochem. Photobiol. 2011, 12, 37–51. [Google Scholar]
- Thompson, L.K.; Sturtevant, J.M.; Brudvig, G.W. Differential scanning calorimetric studies of photosystem II: Evidence for a structural role for cytochrome b559 in the oxygen-evolving complex. Biochemistry 1986, 25, 6161–6169. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.K.; Blaylock, R.; Sturtevant, J.M.; Brudvig, G.W. Molecular basis of the heat denaturation of photosystem II. Biochemistry 1989, 28, 6686–6695. [Google Scholar] [CrossRef] [PubMed]
- Blume, A. Biological calorimetry: Membranes. Thermochim. Acta 1991, 193, 299–347. [Google Scholar] [CrossRef]
- Mineev, K.S. Development of NMR Spectroscopy Methods and Their Application to Investigate Oligomerization of Membrane Proteins; Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry: Moscow, Russia, 2019. [Google Scholar]
- Krivanek, R.; Jeworrek, C.; Czeslik, C.; Winter, R. Composition Fluctuations in Phospholipid-Sterol Vesicles—A Small-angle Neutron Scattering Study. Z. Phys. Chem. 2008, 222, 1679–1692. [Google Scholar] [CrossRef]
- Krivanek, R.; Dau, H.; Haumann, M. Enthalpy Changes during Photosynthetic Water Oxidation Tracked by Time-Resolved Calorimetry Using a Photothermal Beam Deflection Technique. Biophys. J. 2008, 94, 1890–1903. [Google Scholar] [CrossRef]
- Neupane, S.; De Smet, Y.; Renner, F.U.; Losada-Pérez, P. Quartz crystal microbalance with dissipation monitoring: A versatile tool to monitor phase transitions in biomimetic membranes. Front. Mater. 2018, 5, 46. [Google Scholar] [CrossRef]
- Bryant, C.M.; McClements, D.J. Ultrasonic spectroscopy study of relaxation and scattering in whey protein solutions. J. Sci. Food Agric. 1999, 79, 1754–1760. [Google Scholar] [CrossRef]
- McClements, D.J. Ultrasonic measurements in particle size analysis. Encycl. Anal. Chem. 2000, 5581–5588. [Google Scholar]
- Corredig, M.; Alexander, M.; Dalgleish, D.G. The application of ultrasonic spectroscopy to the study of the gelation of milk components. Food Res. Int. 2004, 37, 557–565. [Google Scholar] [CrossRef]
- Taylor, T.M.; Davidson, P.M.; Bruce, B.D.; Weiss, J. Ultrasonic spectroscopy and differential scanning calorimetry of liposomal-encapsulated nisin. J. Agric. Food Chem. 2005, 53, 8722–8728. [Google Scholar] [CrossRef] [PubMed]
- Kertész, I.; Nagy, D.; Baranyai, L.; Pásztor-Huszár, K.; Varsányi, K.; Le Phuong Nguyen, L.; Felföldi, J. Development of a Novel Ultrasonic Spectroscopy Method for Estimation of Viscosity Change during Milk Clotting. Molecules 2021, 26, 5906. [Google Scholar] [CrossRef]
- Sarvazyan, A.P. Ultrasonic velocimetry of biological compounds. Annu. Rev. Biophys. Biophys. Chem. 1991, 20, 321–342. [Google Scholar] [CrossRef]
- Eggers, F.; Funck, T. Ultrasonic measurements with milliliter liquid samples in the 0.5–100 MHz range. Rev. Sci. Instrum. 1973, 44, 969–977. [Google Scholar] [CrossRef]
- Sarvazyan, A.P.; Kharakoz, D.P. Small Volume Differential Interferometer for Velocity and Absorption Measurements of Ultrasound. Prib. Tekhn. Eksper. 1981, 203–206. [Google Scholar]
- Kharakoz, D.P. Investigation of the State and Phase Transitions of Protein Molecules and Lipid Membranes by Measuring the Volume and Compressibility of Solutions; Institute of Theoretical and Experimental Biophysics of Russian Academy of Sciences: Moscow, Russia, 1995. [Google Scholar]
- Blumenfeld, L.A. Problems of Biological Physics; Nauka: Moscow, Russia, 1974; p. 335. [Google Scholar]
- Astashev, M.; Belosludtsev, K.; Kharakoz, D. Method for digital measurement of phase-frequency characteristics for a fixed-length ultrasonic spectrometer. Acoust. Phys. 2014, 60, 335–341. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Astashev, M.E.; Penkov, N.V.; Gudkov, S.V.; Dyachenko, I.A.; Samartsev, V.N.; Belosludtsev, K.N. Effects of Phospholipase A2 Inhibitors on Bilayer Lipid Membranes. J. Membr. Biol. 2016, 249, 339–347. [Google Scholar] [CrossRef]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Agafonov, A.V.; Astashev, M.E.; Kazakov, A.S.; Saris, N.-E.L.; Mironova, G.D. Ca2+-dependent permeabilization of mitochondria and liposomes by palmitic and oleic acids: A comparative study. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 2600–2606. [Google Scholar] [CrossRef]
- MacDonald, R.C.; MacDonald, R.I.; Menco, B.P.M.; Takeshita, K.; Subbarao, N.K.; Hu, L.-r. Small-volume extrusion apparatus for preparation of large, unilamellar vesicles. Biochim. Biophys. Acta (BBA) Biomembr. 1991, 1061, 297–303. [Google Scholar] [CrossRef]
- Preparing Large, Unilamellar Vesicles by Extrusion (LUVET). Available online: https://avantilipids.com/tech-support/liposome-preparation/luvet (accessed on 13 September 2022).
- Drazenovic, J.; Wang, H.; Roth, K.; Zhang, J.; Ahmed, S.; Chen, Y.; Bothun, G.; Wunder, S.L. Effect of lamellarity and size on calorimetric phase transitions in single component phosphatidylcholine vesicles. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 532–543. [Google Scholar] [CrossRef] [PubMed]
- van Leeuwen, P.J.; Nieveen, M.C.; van de Meent, E.J.; Dekker, J.P.; van Gorkom, H.J. Rapid and simple isolation of pure photosystem II core and reaction center particles from spinach. Photosynth. Res. 1991, 28, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Khorobrykh, A. Hydrogen Peroxide and Superoxide Anion Radical Photoproduction in PSII Preparations at Various Modifications of the Water-Oxidizing Complex. Plants 2019, 8, 329. [Google Scholar] [CrossRef]
- Ford, R.; Evans, M. Isolation of a photosystem 2 preparation from higher plants with highly enriched oxygen evolution activity. FEBS Lett. 1983, 160, 159–164. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Meister, A.; Kerth, A.; Blume, A. Interaction of sodium dodecyl sulfate with dimyristoyl-sn-glycero-3-phosphocholine monolayers studied by infrared reflection absorption spectroscopy. A new method for the determination of surface partition coefficients. J. Phys. Chem. B 2004, 108, 8371–8378. [Google Scholar] [CrossRef]
- López, O.; Cócera, M.; Pons, R.; Azemar, N.; De la Maza, A. Kinetic studies of liposome solubilization by sodium dodecyl sulfate based on a dynamic light scattering technique. Langmuir 1998, 14, 4671–4674. [Google Scholar] [CrossRef]
- Hildebrand, A.; Beyer, K.; Neubert, R.; Garidel, P.; Blume, A. Temperature dependence of the interaction of cholate and deoxycholate with fluid model membranes and their solubilization into mixed micelles. Colloids Surf. B Biointerfaces 2003, 32, 335–351. [Google Scholar] [CrossRef]
- Juan-Colás, J.; Dresser, L.; Morris, K.; Lagadou, H.; Ward, R.H.; Burns, A.; Tear, S.; Johnson, S.; Leake, M.C.; Quinn, S.D. The Mechanism of Vesicle Solubilization by the Detergent Sodium Dodecyl Sulfate. Langmuir 2020, 36, 11499–11507. [Google Scholar] [CrossRef]
- Deo, N.; Somasundaran, P. Effects of sodium dodecyl sulfate on mixed liposome solubilization. Langmuir 2003, 19, 7271–7275. [Google Scholar] [CrossRef]
- Jafari, M.; Mehrnejad, F.; Rahimi, F.; Asghari, S.M. The molecular basis of the sodium dodecyl sulfate effect on human ubiquitin structure: A molecular dynamics simulation study. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lillie, M.; Gosline, J. Effects of lipids on elastin’s viscoelastic properties. Biopolym. Orig. Res. Biomol. 2002, 64, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Anosov, A.A.; Smirnova, E.Y.; Korepanova, E.A.; Shogenov, I.M. The effects of SDS at subsolubilizing concentrations on the planar lipid bilayer permeability: Two kinds of current fluctuations. Chem. Phys. Lipids 2019, 218, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Yanagihara, S.-i.; Misono, Y.; Suzuki, M. Effect of fatty acids on phase behavior of hydrated dipalmitoylphosphatidylcholine bilayer: Saturated versus unsaturated fatty acids. Chem. Phys. Lipids 2001, 109, 117–133. [Google Scholar] [CrossRef]
- Ortiz, A.; Gómez-Fernández, J.C. A differential scanning calorimetry study of the interaction of free fatty acids with phospholipid membranes. Chem. Phys. Lipids 1987, 45, 75–91. [Google Scholar] [CrossRef]
- Eynaudi, A.; Díaz-Castro, F.; Bórquez, J.C.; Bravo-Sagua, R.; Parra, V.; Troncoso, R. Differential effects of Oleic and Palmitic Acids on Lipid Droplet-Mitochondria interaction in the hepatic cell line HepG2. Front. Nutr. 2021, 8, 775382. [Google Scholar] [CrossRef]
- Saitta, F.; Motta, P.; Barbiroli, A.; Signorelli, M.; La Rosa, C.; Janaszewska, A.; Klajnert-Maculewicz, B.; Fessas, D. Influence of Free Fatty Acids on Lipid Membrane–Nisin Interaction. Langmuir 2020, 36, 13535–13544. [Google Scholar] [CrossRef]
- Saitta, F.; Signorelli, M.; Fessas, D. Dissecting the effects of free fatty acids on the thermodynamic stability of complex model membranes mimicking insulin secretory granules. Colloids Surf. B Biointerfaces 2019, 176, 167–175. [Google Scholar] [CrossRef]
- Halstenberg, S.; Heimburg, T.; Hianik, T.; Kaatze, U.; Krivanek, R. Cholesterol-induced variations in the volume and enthalpy fluctuations of lipid bilayers. Biophys. J. 1998, 75, 264–271. [Google Scholar] [CrossRef]
- Umena, Y.; Kawakami, K.; Shen, J.-R.; Kamiya, N. Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 Å. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef]
- Loll, B.; Kern, J.; Saenger, W.; Zouni, A.; Biesiadka, J. Lipids in photosystem II: Interactions with protein and cofactors. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1767, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, K.N.; Iverson, T.M.; Maghlaoui, K.; Barber, J.; Iwata, S. Architecture of the photosynthetic oxygen-evolving center. Science 2004, 303, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Zouni, A.; Witt, H.-T.; Kern, J.; Fromme, P.; Krauss, N.; Saenger, W.; Orth, P. Crystal structure of photosystem II from Synechococcus elongatus at 3.8 Å resolution. Nature 2001, 409, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Klimov, V.; Shafiev, M.; Allakhverdiev, S. Photoinactivation of the reactivation capacity of photosystem II in pea subchloroplast particles after a complete removal of manganese. Photosynth. Res. 1990, 23, 59–65. [Google Scholar] [CrossRef]
- Jegerschoeld, C.; Virgin, I.; Styring, S. Light-dependent degradation of the D1 protein in photosystem II is accelerated after inhibition of the water splitting reaction. Biochemistry 1990, 29, 6179–6186. [Google Scholar] [CrossRef]
- Telfer, A.; De Las Rivas, J.; Barber, J. β-Carotene within the isolated Photosystem II reaction centre: Photooxidation and irreversible bleaching of this chromophore by oxidised P680. Biochim. Biophys. Acta (BBA) Bioenerg. 1991, 1060, 106–114. [Google Scholar] [CrossRef]
- Yanykin, D.; Khorobrykh, A.; Zastrizhnaya, O.; Klimov, V. Interaction of molecular oxygen with the donor side of photosystem II after destruction of the water-oxidizing complex. Biochemistry 2014, 79, 205–212. [Google Scholar] [CrossRef]
- Theg, S.M.; Filar, L.J.; Dilley, R.A. Photoinactivation of chloroplasts already inhibited on the oxidizing side of Photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 1986, 849, 104–111. [Google Scholar] [CrossRef]
- Rao, N.; Mehra, S.; Bridges, J.; Venkatraman, S. Experimental point spread function of FM pulse imaging scheme. Ultrason. Imaging 1995, 17, 114–141. [Google Scholar] [CrossRef]
- Tobocman, W.; Driscoll, D.; Shokrollahi, N.; Izatt, J. Free of speckle ultrasound images of small tissue structures. Ultrasonics 2002, 40, 983–996. [Google Scholar] [CrossRef]
- Amirmazlaghani, M.; Amindavar, H. Wavelet domain Bayesian processor for speckle removal in medical ultrasound images. IET Image Process. 2012, 6, 580–588. [Google Scholar] [CrossRef]
- Gan, T.; Hutchins, D.; Billson, D.; Schindel, D. The use of broadband acoustic transducers and pulse-compression techniques for air-coupled ultrasonic imaging. Ultrasonics 2001, 39, 181–194. [Google Scholar] [CrossRef]
- Iizuka, Y. High signal-to-noise ratio ultrasonic testing system using chirp pulse compression. Insight 1998, 40, 282–285. [Google Scholar]
- Moreau, A.; Lévesque, D.; Lord, M.; Dubois, M.; Monchalin, J.-P.; Padioleau, C.; Bussière, J. On-line measurement of texture, thickness and plastic strain ratio using laser-ultrasound resonance spectroscopy. Ultrasonics 2002, 40, 1047–1056. [Google Scholar] [CrossRef]
- Potter, M.; Dixon, S.; Morrison, J.; Suliamann, A. Development of an advanced multimode automatic ultrasonic texture measurement system for laboratory and production line application. Ultrasonics 2006, 44, e813–e817. [Google Scholar] [CrossRef]
- Leemans, V.; Destain, M.-F. Ultrasonic internal defect detection in cheese. J. Food Eng. 2009, 90, 333–340. [Google Scholar] [CrossRef]
- Hæggström, E.; Luukkala, M. Ultrasound detection and identification of foreign bodies in food products. Food Control 2001, 12, 37–45. [Google Scholar] [CrossRef]
- Létang, C.; Piau, M.; Verdier, C.; Lefebvre, L. Characterization of wheat-flour–water doughs: A new method using ultrasound. Ultrasonics 2001, 39, 133–141. [Google Scholar] [CrossRef]
- Cho, B.K.; Irudayaraj, J. A noncontact ultrasound approach for mechanical property determination of cheeses. J. Food Sci. 2003, 68, 2243–2247. [Google Scholar] [CrossRef]
- Rao, N. Investigation of a pulse compression technique for medical ultrasound: A simulation study. Med. Biol. Eng. Comput. 1994, 32, 181–188. [Google Scholar] [CrossRef]
- Gan, T.; Hutchins, D.; Billson, D. Preliminary studies of a novel air-coupled ultrasonic inspection system for food containers. J. Food Eng. 2002, 53, 315–323. [Google Scholar] [CrossRef]
- Benedito, J.; Carcel, J.; Clemente, G.; Mulet, A. Cheese maturity assessment using ultrasonics. J. Dairy Sci. 2000, 83, 248–254. [Google Scholar] [CrossRef]
- Benedito, J.; Simal, S.; Clemente, G.; Mulet, A. Manchego cheese texture evaluation by ultrasonics and surface probes. Int. Dairy J. 2006, 16, 431–438. [Google Scholar] [CrossRef]
- El Kadi, Y.A.; Moudden, A.; Faiz, B.; Maze, G.; Decultot, D. Ultrasonic monitoring of fish thawing process optimal time of thawing and effect of freezing/thawing. Acta Sci. Pol. Technol. Aliment. 2013, 12, 273–281. [Google Scholar]
- Kargerová, A.; Pekař, M. High-Resolution Ultrasonic Spectroscopy Study of Interactions between Hyaluronan and Cationic Surfactants. Langmuir 2014, 30, 11866–11872. [Google Scholar] [CrossRef]
- Cardona, T.; Sedoud, A.; Cox, N.; Rutherford, A.W. Charge separation in Photosystem II: A comparative and evolutionary overview. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1817, 26–43. [Google Scholar] [CrossRef]
- Guskov, A.; Kern, J.; Gabdulkhakov, A.; Broser, M.; Zouni, A.; Saenger, W. Cyanobacterial photosystem II at 2.9-Å resolution and the role of quinones, lipids, channels and chloride. Nat. Struct. Mol. Biol. 2009, 16, 334–342. [Google Scholar] [CrossRef]
- Mizusawa, N.; Wada, H. The role of lipids in photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1817, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Shutilova, N.; Semenova, G.; Klimov, V.; Shnyrov, V. Temperature-induced functional and structural transformations of the photosystem II oxygen-evolving complex in spinach subchloroplast preparations. Biochem. Mol. Biol. Int. 1995, 35, 1233–1243. [Google Scholar]
- Lípová, L.; Krchňák, P.; Komenda, J.; Ilík, P. Heat-induced disassembly and degradation of chlorophyll-containing protein complexes in vivo. Biochim. Biophys. Acta (BBA) Bioenerg. 2010, 1797, 63–70. [Google Scholar] [CrossRef]
- Kirchhoff, H.; Borinski, M.; Lenhert, S.; Chi, L.; Büchel, C. Transversal and lateral exciton energy transfer in grana thylakoids of spinach. Biochemistry 2004, 43, 14508–14516. [Google Scholar] [CrossRef] [PubMed]
- Vass, I. Molecular mechanisms of photodamage in the Photosystem II complex. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1817, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Tyystjärvi, E. Photoinhibition of photosystem II. Int. Rev. Cell Mol. Biol. 2013, 300, 243–303. [Google Scholar]
- De Las Rivas, J.; Barber, J. Structure and thermal stability of photosystem II reaction centers studied by infrared spectroscopy. Biochemistry 1997, 36, 8897–8903. [Google Scholar] [CrossRef] [PubMed]
- Okamura, M.; Satoh, K.; Isaacson, R.; Feher, G. Evidence of the primary charge separation in the D1/D2 complex of Photosystem II from spinach: EPR of the triplet state. Prog. Photosynth. Res. 1987, 1, 379–381. [Google Scholar]
- Telfer, A.; Barber, J.; Evans, M. Oxidation-reduction potential dependence of reaction centre triplet formation in the isolated D1/D2/cytochrome b-559 photosystem II complex. FEBS Lett. 1988, 232, 209–213. [Google Scholar] [CrossRef]
- Telfer, A.; Bishop, S.M.; Phillips, D.; Barber, J. Isolated photosynthetic reaction center of photosystem II as a sensitizer for the formation of singlet oxygen. Detection and quantum yield determination using a chemical trapping technique. J. Biol. Chem. 1994, 269, 13244–13253. [Google Scholar] [CrossRef]
- Ananyev, G.; Klimov, V. Photoproduction bound hydrogen peroxide in subchloroplast preparations of photosystem II. Dokl AN SSSR 1988, 298, 1007–1011. [Google Scholar]
- Ananyev, G.; Wydrzynski, T.; Renger, G.; Klimov, V. Transient peroxide formation by the manganese-containing, redox-active donor side of photosystem II upon inhibition of O2 evolution with lauroylcholine chloride. Biochim. Biophys. Acta (BBA) Bioenerg. 1992, 1100, 303–311. [Google Scholar] [CrossRef]
- Ananyev, G.; Renger, G.; Wacker, U.; Klimov, V. The photoproduction of superoxide radicals and the superoxide dismutase activity of Photosystem II. The possible involvement of cytochrome b559. Photosynth. Res. 1994, 41, 327–338. [Google Scholar] [CrossRef]
- Wydrzynski, T.; Vänngård, T. H2O2 formation by photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 1989, 973, 23–28. [Google Scholar] [CrossRef]
- Klimov, V.; Ananyev, G.; Zastryzhnaya, O.; Wydrzynski, T.; Renger, G. Photoproduction of hydrogen peroxide in Photosystem II membrane fragments: A comparison of four signals. Photosynth. Res. 1993, 38, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Strzałka, K. Dark reoxidation of the plastoquinone-pool is mediated by the low-potential form of cytochrome b-559 in spinach thylakoids. Photosynth. Res. 1999, 62, 273–279. [Google Scholar] [CrossRef]
- Khorobrykh, S.; Ivanov, B. Oxygen reduction in a plastoquinone pool of isolated pea thylakoids. Photosynth. Res. 2002, 71, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Khorobrykh, S.A.; Khorobrykh, A.A.; Yanykin, D.V.; Ivanov, B.N.; Klimov, V.V.; Mano, J.i. Photoproduction of catalase-insensitive peroxides on the donor side of manganese-depleted photosystem II: Evidence with a specific fluorescent probe. Biochemistry 2011, 50, 10658–10665. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P.; Šnyrychová, I.; Kruk, J.; Strzałka, K.; Nauš, J. Evidence that cytochrome b 559 is involved in superoxide production in photosystem II: Effect of synthetic short-chain plastoquinones in a cytochrome b 559 tobacco mutant. Biochem. J. 2006, 397, 321–327. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of reactive oxygen species by photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 2009, 1787, 1151–1160. [Google Scholar] [CrossRef]
- Pospíšil, P. Molecular mechanisms of production and scavenging of reactive oxygen species by photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1817, 218–231. [Google Scholar] [CrossRef]
- Yanykin, D.; Khorobrykh, A.; Terentyev, V.; Klimov, V. Two pathways of photoproduction of organic hydroperoxides on the donor side of photosystem 2 in subchloroplast membrane fragments. Photosynth. Res. 2017, 133, 129–138. [Google Scholar] [CrossRef]
- Barbato, R.; Friso, G.; Ponticos, M.; Barber, J. Characterization of the Light-induced Cross-linking of the α-Subunit of Cytochrome b559 and the D1 Protein in Isolated Photosystem II Reaction Centers (∗). J. Biol. Chem. 1995, 270, 24032–24037. [Google Scholar] [CrossRef]
- Henmi, T.; Yamasaki, H.; Sakuma, S.; Tomokawa, Y.; Tamura, N.; Shen, J.-R.; Yamamoto, Y. Dynamic Interaction between the D1 protein, CP43 and OEC33 at the lumenal side of photosystem II in spinach chloroplasts: Evidence from light-induced cross-Linking of the proteins in the donor-side photoinhibition. Plant Cell Physiol. 2003, 44, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Nakatani, E.; Henmi, T.; Ferjani, A.; Harada, Y.; Tamura, N.; Yamamoto, Y. Turnover of the aggregates and cross-linked products of the D1 protein generated by acceptor-side photoinhibition of photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 1999, 1413, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Salter, A.; De Las Rivas, J.; Barber, J.; Andersson, B. On the molecular mechanisms of light induced D1 protein degradation. Res. Photosynth. 1992, 4, 395–402. [Google Scholar]
- Singh, A.K.; Singhal, G. Formation of cross-linking between photosystem 2 proteins during irradiation of thylakoid membranes at high temperature. Photosynthetica 1999, 36, 213–223. [Google Scholar] [CrossRef]
- Seidler, A. Intermolecular and Intramolecular Interactions of the 33-kDa Protein in Photosystem II. Eur. J. Biochem. 1996, 242, 485–490. [Google Scholar] [CrossRef]
- Shutova, T.; Nikitina, J.; Deikus, G.; Andersson, B.; Klimov, V.; Samuelsson, G. Structural dynamics of the manganese-stabilizing protein effect of pH, calcium, and manganese. Biochemistry 2005, 44, 15182–15192. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanykin, D.V.; Astashev, M.E.; Khorobrykh, A.A.; Paskhin, M.O.; Serov, D.A.; Gudkov, S.V. Application of Fixed-Length Ultrasonic Interferometry to Determine the Kinetics of Light-/Heat-Induced Damage to Biological Membranes and Protein Complexes. Inventions 2022, 7, 87. https://doi.org/10.3390/inventions7040087
Yanykin DV, Astashev ME, Khorobrykh AA, Paskhin MO, Serov DA, Gudkov SV. Application of Fixed-Length Ultrasonic Interferometry to Determine the Kinetics of Light-/Heat-Induced Damage to Biological Membranes and Protein Complexes. Inventions. 2022; 7(4):87. https://doi.org/10.3390/inventions7040087
Chicago/Turabian StyleYanykin, Denis V., Maxim E. Astashev, Andrey A. Khorobrykh, Mark O. Paskhin, Dmitriy A. Serov, and Sergey V. Gudkov. 2022. "Application of Fixed-Length Ultrasonic Interferometry to Determine the Kinetics of Light-/Heat-Induced Damage to Biological Membranes and Protein Complexes" Inventions 7, no. 4: 87. https://doi.org/10.3390/inventions7040087
APA StyleYanykin, D. V., Astashev, M. E., Khorobrykh, A. A., Paskhin, M. O., Serov, D. A., & Gudkov, S. V. (2022). Application of Fixed-Length Ultrasonic Interferometry to Determine the Kinetics of Light-/Heat-Induced Damage to Biological Membranes and Protein Complexes. Inventions, 7(4), 87. https://doi.org/10.3390/inventions7040087