
Modelling of Climate Change’s Impact on Prunus armeniaca L.’s Flowering Time

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
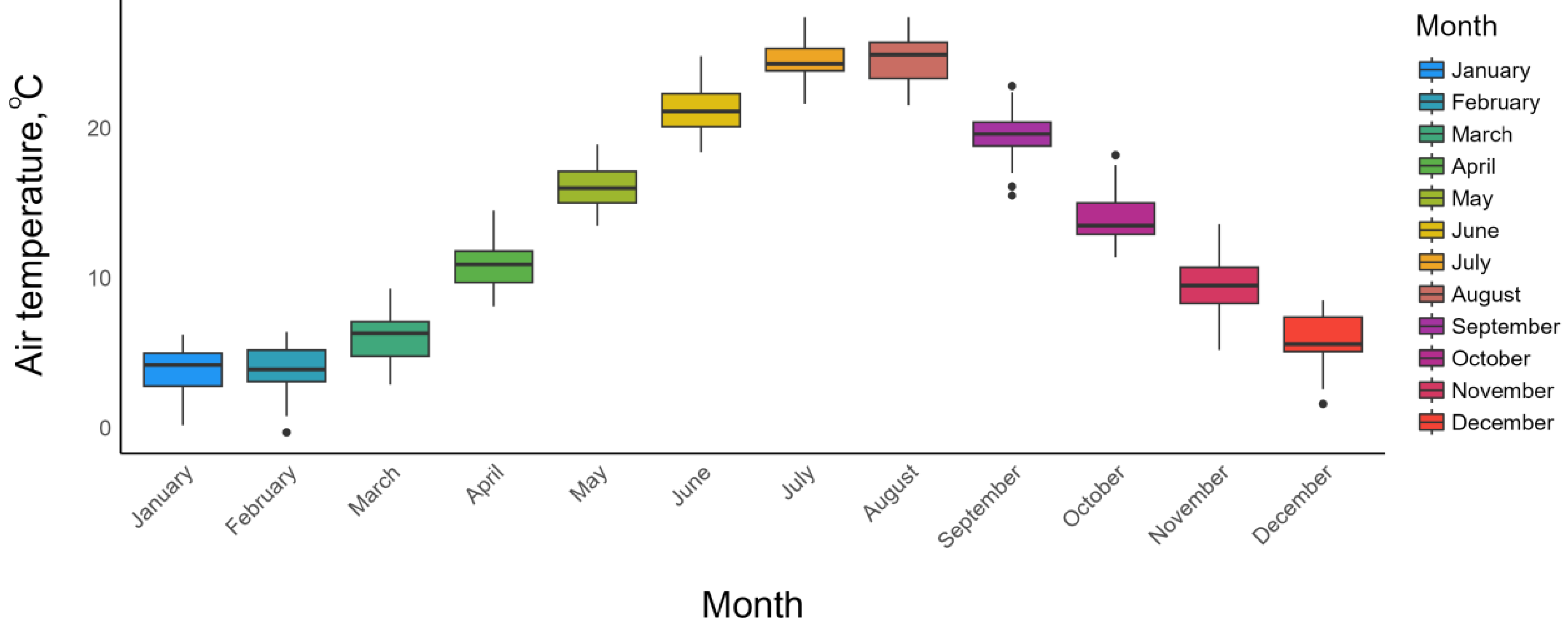
2.1. Phenological, Meteorological Data and Research Site of Prunus armeniaca L.
2.2. Study of the Morphogenesis of Flowering Kidneys
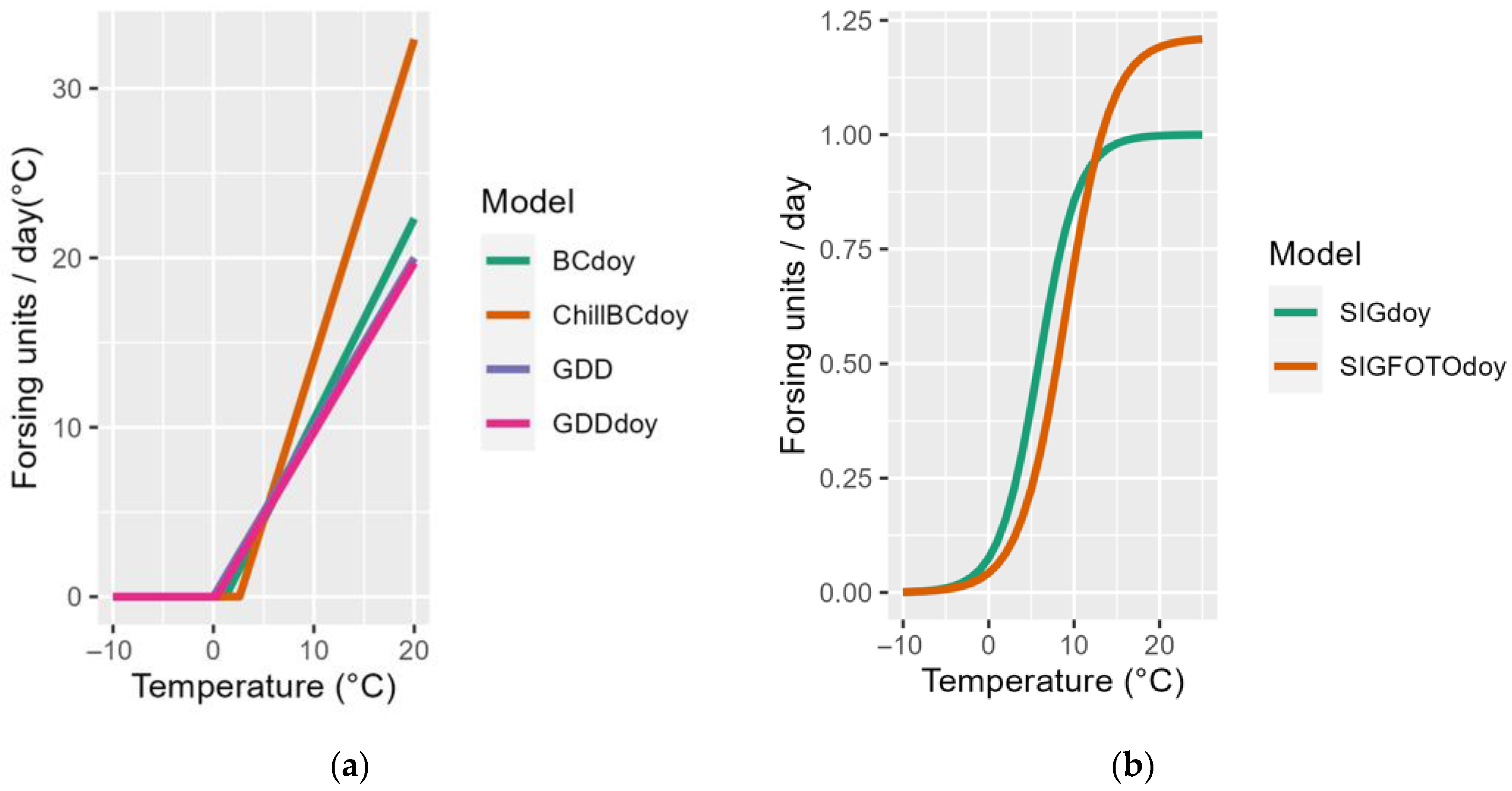
2.3. Phenolic Model
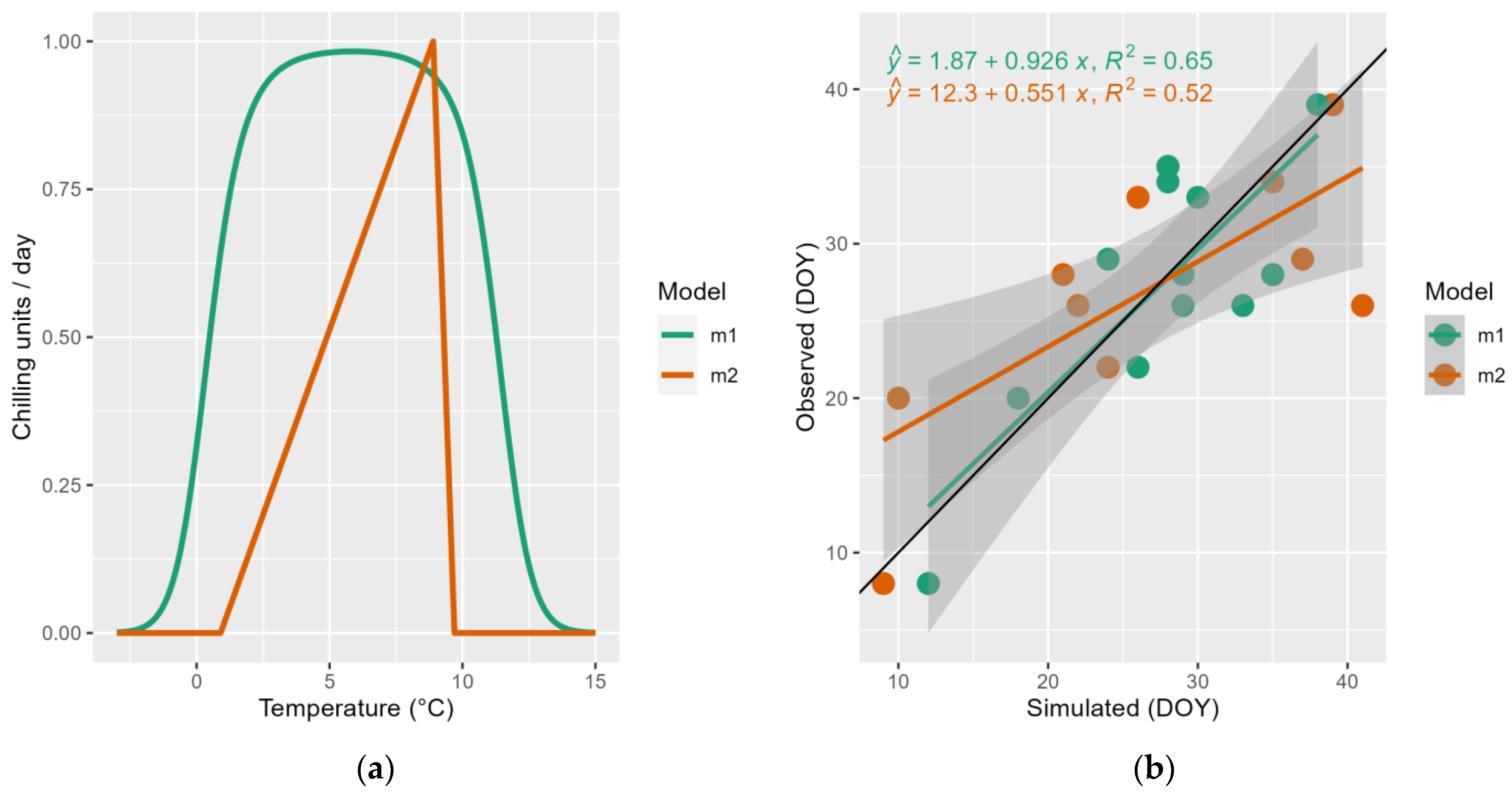
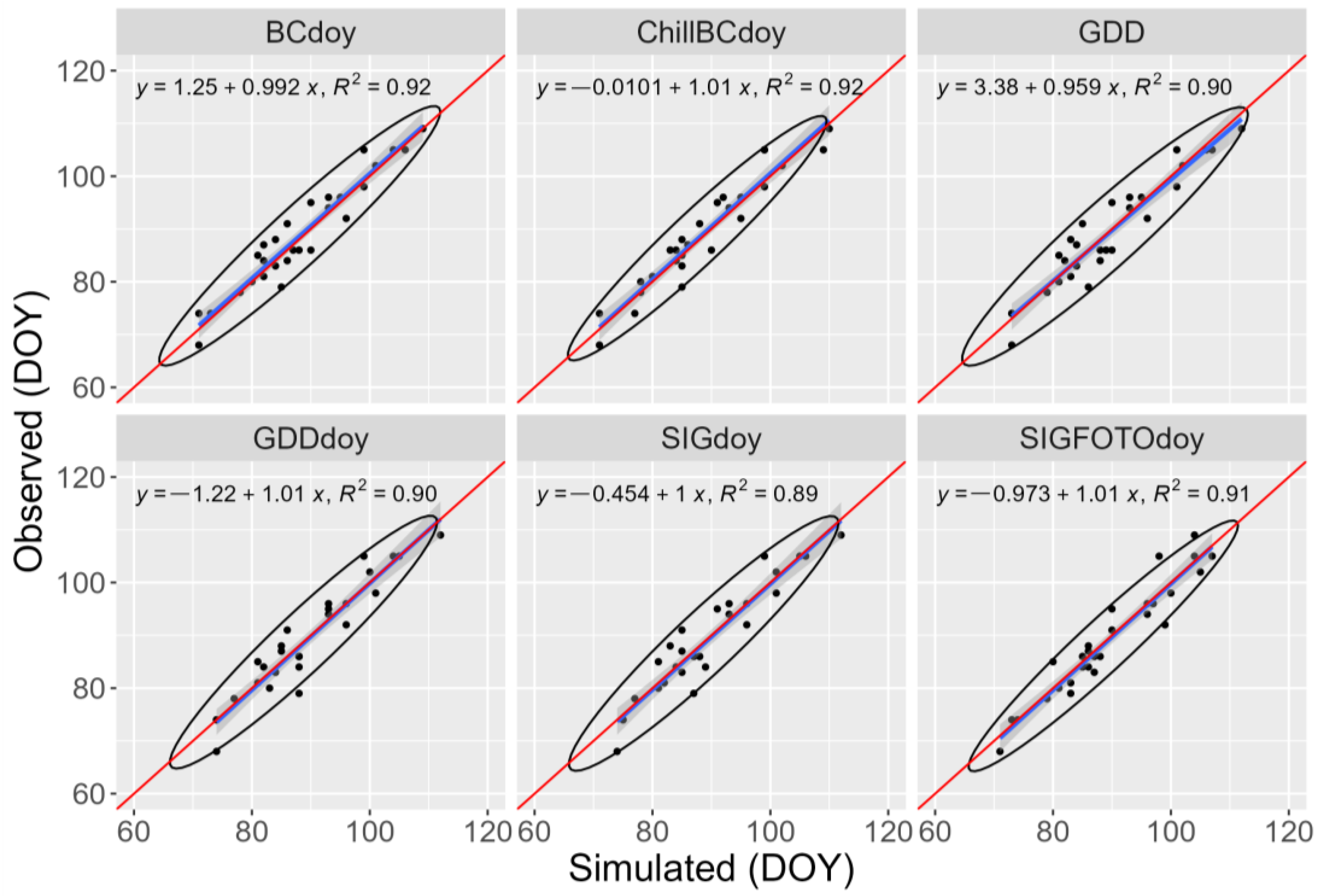
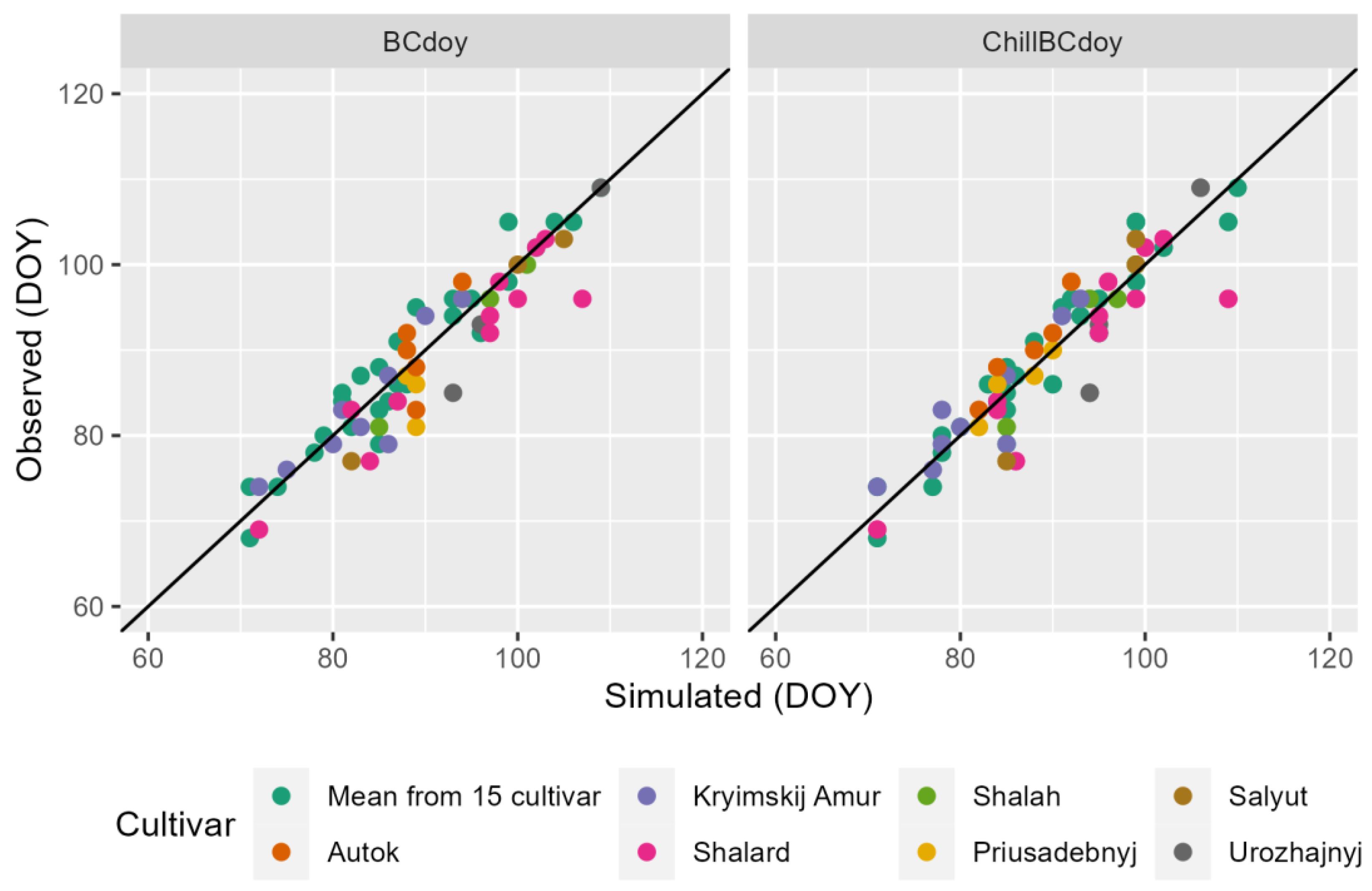
2.4. Model Evaluation
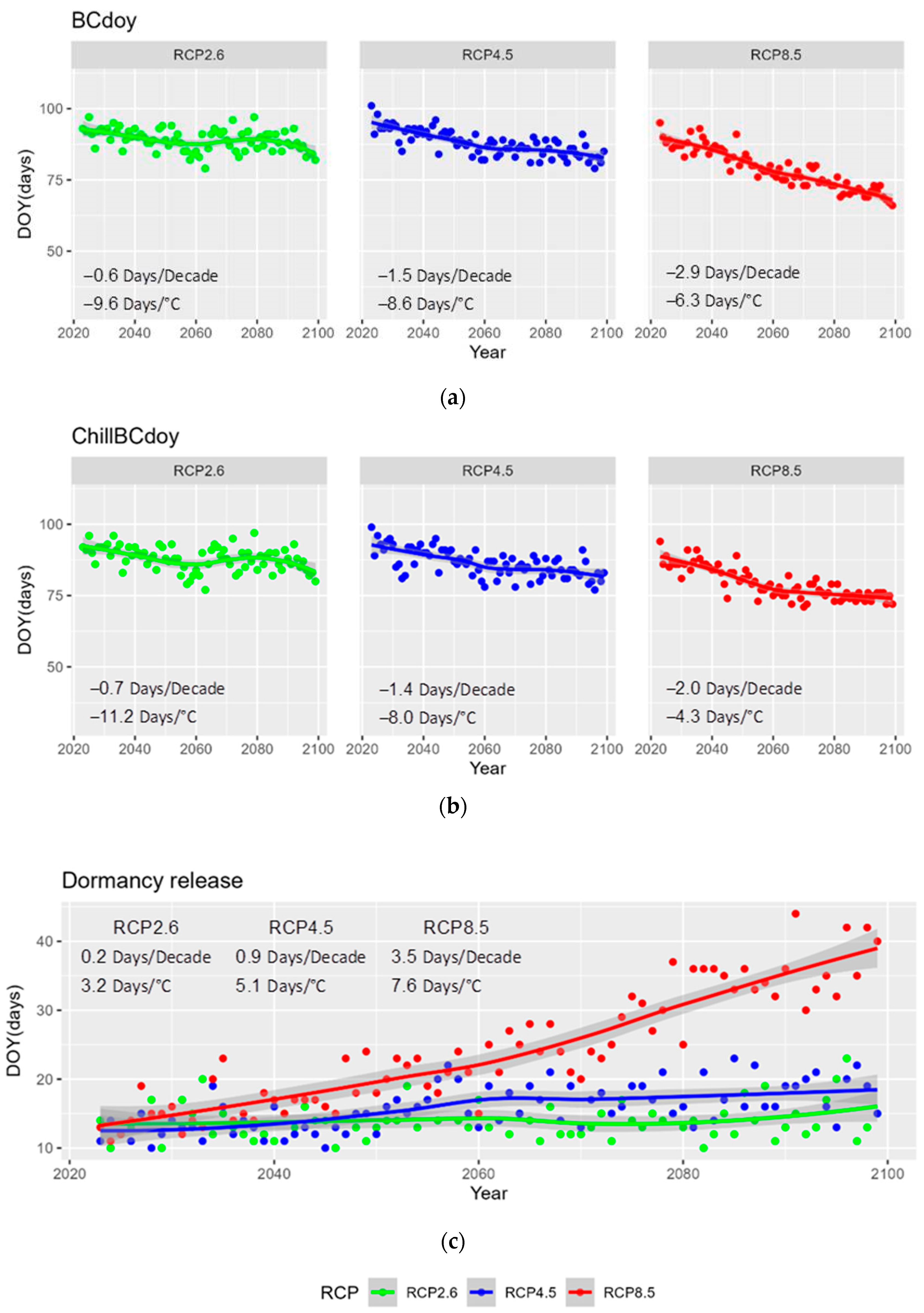
Assessment of Shifts in Spring Phenology Using Climate Scenarios
2.5. Statistical Analysis
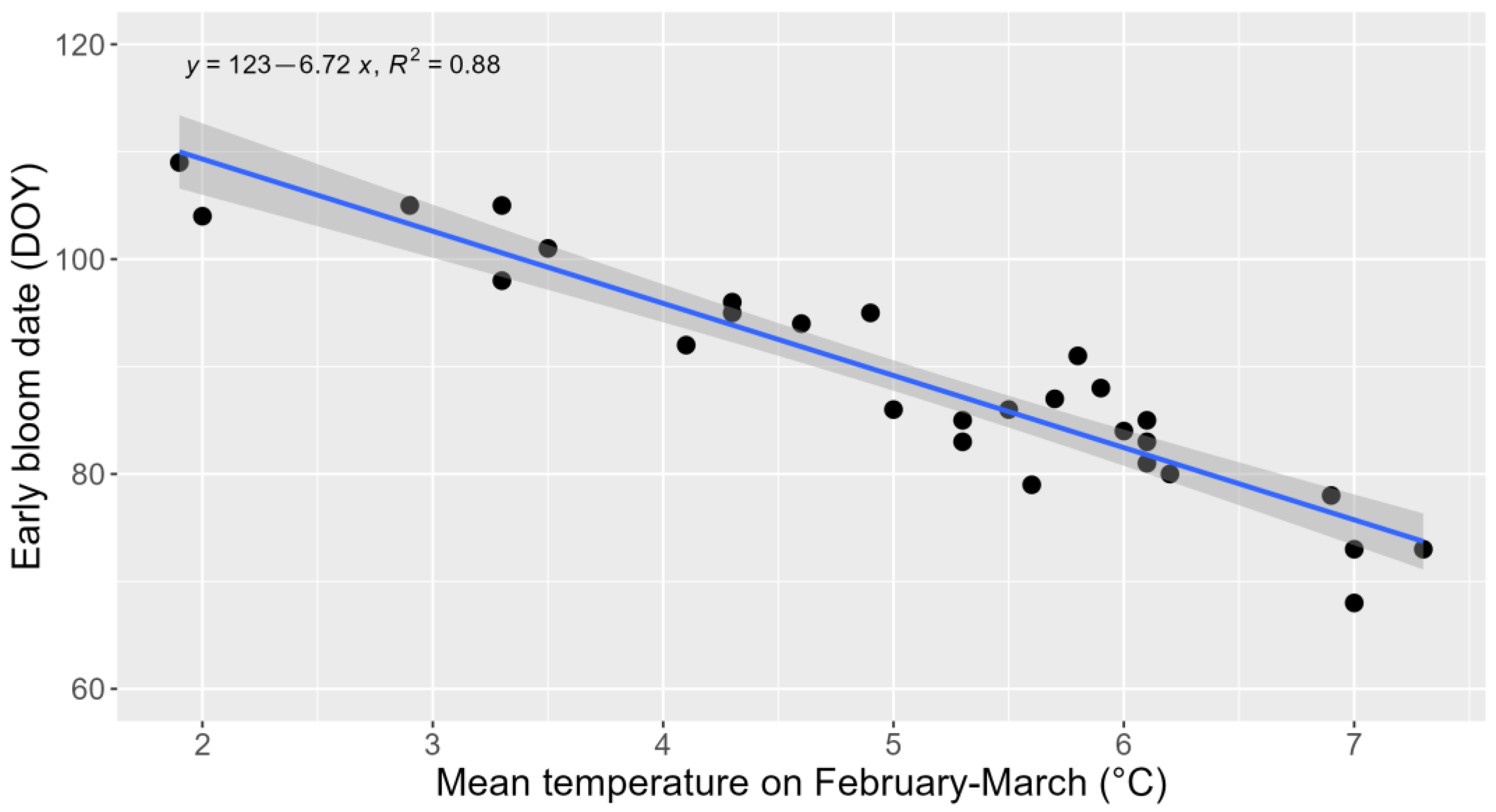
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vujadinović Mandić, M.; Vuković Vimić, A.; Ranković-Vasić, Z.; Đurović, D.; Ćosić, M.; Sotonica, D.; Nikolić, D.; Đurđević, V. Observed Changes in Climate Conditions and Weather-Related Risks in Fruit and Grape Production in Serbia. Atmosphere 2022, 13, 948. [Google Scholar] [CrossRef]
- Parmesan, C.; Hanley, M.E. Plants and climate change: Complexities and surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Grandis, A.; Arenque, B.C.; Buckeridge, M.S. Impacts of climate changes on crop physiology and food quality. Food Res. Int. 2010, 43, 1814–1823. [Google Scholar] [CrossRef]
- Omidifard, Z.; Gharaghani, A. Chilling and heat requirements and bloom frost tolerance of Prunus scoparia (Spach) C. K. Schneid and P. elaeagnifolia Spach. Trees 2022, 36, 1559–1572. [Google Scholar] [CrossRef]
- Herrera, S.; Lora, J.; Fadón, E.; Hedhly, A.; Alonso, J.M.; Hormaza, J.I.; Rodrigo, J. Male Meiosis as a Biomarker for Endoto Ecodormancy Transition in Apricot. Front. Plant. Sci. 2022, 13, 842333. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.; Mojahid, H.; Fadón, E.; Rodrigo, J.; Ruiz, D.; Egea, J.A.; Ben Mimoun, M.; Kodad, O.; El Yaacoubi, A.; Ghrab, M.; et al. Climate change impacts on winter chill in Mediterranean temperate fruit orchards. Reg. Environ. Change 2023, 23, 7. [Google Scholar] [CrossRef]
- Korsakova, S.; Korsakov, P. Features development of olive trees during the growing season in the Southern coast of the Crimea. BIO Web Conf. 2021, 38, 00060. [Google Scholar] [CrossRef]
- Atagul, O.; Calle, A.; Demirel, G.; Lawton, J.M.; Bridges, W.C.; Gasic, K. Estimating Heat Requirement for Flowering in Peach Germplasm. Agronomy 2022, 12, 1002. [Google Scholar] [CrossRef]
- Korzin, V.; Gorina, V.; Saplev, N. The effect of weather conditions in southern Russia on the frost resistance of apricot generative buds. Hortic. Sci. 2021, 48, 158–165. [Google Scholar] [CrossRef]
- Gorina, V.; Korzin, V.; Mesyats, N. Apricot breeding for late flowering in Nikita Botanical Gardens. Acta Hortic. 2020, 1282, 25–30. [Google Scholar] [CrossRef]
- Gorina, V.; Korzin, V.; Mesyats, N.; Pilkevich, R. Drought tolerance in apricot cultivars and breeding forms under the conditions of the southern coast of Crimea. Acta Hortic. 2021, 1324, 413–420. [Google Scholar] [CrossRef]
- Dragavtseva, I.; Kuznetsova, A.P.; Mozhar, N.V.; Klyukina, A.V. Influence of temperature stresses of winter-spring period the fruiting of fruit crops (apricot, pear) in the Krasnodar territory under changing climate conditions. Sci. Work. North. Cauc. Fed. Sci. Cent. Hortic. Vitic. Wine-Mak. 2019, 23, 109–116. (in Russian). [Google Scholar] [CrossRef]
- Dragavceva, I.A.; Kuzneczova, A.P.; Klyukina, A.V.; Kovalenko, S.P. The reaction of apricot tree for stressful situation of winter period taking into account the climate change. Fruit. Grow. Vitic. South. Russ. 2020, 65, 141–150. (in Russian). [Google Scholar] [CrossRef]
- Korzin, V.V. Analysis of the development and contemporary state of apricot culture in the world and Russian Federation. Hortic. Vitic. 2019, 6, 35–41. (in Russian). [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Data. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 4 February 2022).
- Razavi, F.; Hajilou, J.; Tabatabaei, S.J.; Dadpour, M.R. Comparison of chilling and heat requirement in some peach and apricot cultivars. Res. Plant Biol. 2011, 1, 40–47. Available online: https://www.updatepublishing.com/journal/index.php/ripb/article/view/2571 (accessed on 25 January 2023).
- Liu, W.; Liu, N.; Zhang, Y.; Yu, X.; Sun, M.; Xu, M.; Zhang, Q.; Liu, S. Apricot cultivar evolution and breeding program in China. Acta Hortic. 2012, 966, 223–228. [Google Scholar] [CrossRef]
- Gorina, V.M.; Korzin, V.V. Long-term Investigations Results of Low Temperatures Influence on Apricot Generative Buds. Acta Hortic. 2016, 1139, 363–368. [Google Scholar] [CrossRef]
- Mesterházy, I.; Raffai, P.; Szalay, L.; Bozó, L.; Ladányi, M. Estimation of Blooming Start with the Adaptation of the Unified Model for Three Apricot Cultivars (Prunus armeniaca L.) Based on Long-Term Observations in Hungary (1994–2020). Diversity 2022, 14, 560. [Google Scholar] [CrossRef]
- Korzin, V.V. Influence of weather conditions on flowering and productivity of different apricot (Armeniaca vulgaris Lam.) varieties under conditions on the south coast of the Crimea. Tavricheskij Vestn. Agrar. Nauk. 2018, 3, 65–71. (in Russian). Available online: https://tvan.niishk.site/data/documents/7_3.pdf (accessed on 25 January 2023).
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef]
- Korsakova, S.P. Fenologicheskaya reakciya Quercus pubescens Willd. na klimaticheskie izmeneniya v usloviyax suxix subtropikov. Tr. Karadag. Nauchnoj Stn. Im. T.I. Vyazemskogo Prir. Zapov. RAN 2018, 2, 30–46. (in Russian). [Google Scholar] [CrossRef]
- Baumgarten, F.; Zohner, C.M.; Gessler, A.; Vitasse, Y. Chilled to be forced: The 376 best dose to wake up buds from winter dormancy. New. Phytol. 2021, 230, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Zohner, C.; Benito, B.; Svenning, J.-C.; Renner, S.S. Day length unlikely to constrain climate-driven shifts in leaf-out times of northern woody plants. Nat. Clim. Change 2016, 6, 1120–1123. [Google Scholar] [CrossRef]
- Gauzere, J.; Lucas, C.; Ronce, O.; Davi, H.; Chuine, I. Sensitivity analysis of tree phenology models reveals increasing sensitivity of their predictions to winter chilling temperature and photoperiod with warming climate. Ecol. Model. 2019, 411, 108805. [Google Scholar] [CrossRef]
- Fila, G.; Di Lena, B.; Gardiman, M.; Storchi, P.; Tomasi, D.; Silvestroni, O.; Pitacco, A. Calibration and validation of grapevine budburst models using growth-room experiments as data source. Agric. For. Meteorol. 2012, 160, 69–79. [Google Scholar] [CrossRef]
- Hlaszny, E. A Szolo (Vitis vinifera L.) Korai Fenológiai Válaszadásának Modellezése a Kunsági Borvidéken Növényfelvételezések, Idojárási Megfigyelések és Regionális Klímamodell Alapján. Ph.D. Thesis, Corvinus University of Budapest, Budapest, Hungary, 2012; 163p. (in Hungarian). Available online: http://phd.lib.uni-corvinus.hu/669/3/Hlaszny_Edit_thu.pdf (accessed on 25 January 2023).
- Williams, D.W.; Andris, H.L.; Beede, R.H.; Luvisi, D.A.; Norton, M.V.K.; Williams, L.E. Validation of a Model for the Growth and Development of the Thompson Seedless Grapevine. II. Phenology. Am. J. Enol. Vitic. 1985, 36, 283–289. [Google Scholar] [CrossRef]
- Cesaraccio, C.; Spano, D.; Snuder, R.L.; Duce, P. Chilling and forcing model to predict bud-burst of crop and forest species. Agric. For. Meteorol. 2004, 126, 1–13. [Google Scholar] [CrossRef]
- Drogoudi, P.; Cantín, C.M.; Brandi, F.; Butcaru, A.; Cos-Terrer, J.; Cutuli, M.; Foschi, S.; Galindo, A.; García-Brunton, J.; Luedeling, E.; et al. Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology. Plants 2023, 12, 584. [Google Scholar] [CrossRef]
- Samigulina, N.S. Practical Work Breeding and Cultivating Fruit and Berry Crops; MichGAU: Michurinsk, Russia, 2006; 197p, Available online: http://window.edu.ru/resource/493/64493/files/0242.pdf (accessed on 25 January 2023).
- Forsythe, W.C.; Rykiel, E.J., Jr.; Stahl, R.S.; Wu, H.-I.; Schoolfield, R.M. A model comparison for daylength as a function of latitude and day of year. Ecol. Model. 1995, 80, 87–95. [Google Scholar] [CrossRef]
- Chuine, I.; de Cortazar-Atauri, I.G.; Kramer, K.; Hänninen, H. Plant development models. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2003; pp. 217–235. [Google Scholar] [CrossRef]
- Chuine, I. A unified model for budburst of trees. J. Theor. Biol. 2000, 207, 337–347. [Google Scholar] [CrossRef]
- Hänninen, H. Modelling bud dormancy release in trees from cool and temperate regions. Acta For. Fenn. 1990, 213, 1–47. [Google Scholar] [CrossRef]
- Blümel, K.; Chmielewski, F.-M. Shortcomings of classical phenological forcing models and a way to overcome them. Agric. For. Meteorol. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Migliavacca, M.; Sonnentag, O.; Keenan, T.F.; Cescatti, A.; O′Keefe, J.; Richardson, A.D. On the uncertainty of phenological responses to climate change, and implications for a terrestrial biosphere model. Biogeosciences 2012, 9, 2063–2083. [Google Scholar] [CrossRef]
- Korsakova, S.P.; Plugatar, Y.V.; Sharmagiy, A.K. Predictive simulation of spring development of Cydalima perspectalis Walker in the south of Russia. IOP Conf. Ser. Earth Environ. Sci. 2022, 949, 012004. [Google Scholar] [CrossRef]
- Korsakova, S.P. The evaluation of future climate change in the Southern coast of the Crimea. Ekosistemy 2018, 15, 151–165. Available online: http://ekosystems.cfuv.ru/wp-content/uploads/2018/12/018korsakova.pdf (accessed on 25 January 2023). (in Russian).
- Gauzere, J.; Delzon, S.; Davi, H.; Bonhomme, M.; de Cortazar-Atauri, I.G.; Chuine, I. Integrating interactive effects of chilling and photoperiod in phenological process-based models. A case study with two European tree species: Fagus sylvatica and Quercus petraea. Agric. For. Meteorol. 2017, 244–245, 9–20. [Google Scholar] [CrossRef]
- The Federal State Budgetary Institution “Voeikov Main Geophysical Observatory” (FGBI “MGO”). Chapter «Climate Change in Russia in 21 Century (Modeli CMIP 5)». Available online: http://voeikovmgo.ru/index.php/izmenenie-klimata-v-rossii-v-xxi-veke (accessed on 15 February 2023). (in Russian).
- The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 15 February 2023).
- Julian, C.; Herrero, M.; Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environ. Exp. Bot. 2014, 100, 20–25. [Google Scholar] [CrossRef]
- Hillmann, L.; Elsysy, M.; Goeckeritz, C.; Hollender, C.; Rothwell, N.; Blanke, M.; Einhorn, T. Preanthesis changes in freeze resistance, relative water content, and ovary growth preempt bud phenology and signify dormancy release of sour cherry floral buds. Planta 2021, 254, 74. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for “Redhaven” and “Elberta” peach trees. Hortic. Sci. 1974, 9, 331–332. [Google Scholar] [CrossRef]
- Andreini, L.; Cortázar-Atauri, I.G.; Chuine, I.; Viti, R.; Bartolini, S.; Ruiz, D.; Campoy, J.A.; Legave, J.M.; Audergon, J.-M.; Bertuzzi, P. Understanding dormancy release in apricot flower buds (Prunus armeniaca L.) using several process-based phenological models. Agric. For. Meteorol. 2014, 184, 210–219. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Gasic, K. Peach [Prunus persica (L.) Batsch] cultivars differ in apparent base temperature and growing degree hour requirement for floral bud break. Front. Plant Sci. 2022, 13, 801606. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, J.H. Chilling requirements of peach varieties. Proc. Am. Soc. Horticul. Sci. 1950, 56, 122–128. Available online: https://www.scirp.org/reference/ReferencesPapers.aspx?ReferenceID=433046 (accessed on 15 February 2023).
- Cantin, C.M.; Wang, X.W.; Almira, M.; Arús, P.; Eduardo, I. Inheritance and QTL analysis of chilling and heat requirements for flowering in an interspecific almond × peach (Texas × Earlygold) F2 population. Euphytica 2020, 216, 51. [Google Scholar] [CrossRef]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and Heat Requirements of Temperate Stone Fruit Trees (Prunus sp.). Agronomy 2020, 10, 409. [Google Scholar] [CrossRef]
- Hackett, W.P.; Hartmann, H.T. The Influence of Temperature on Floral Initiation in the Olive. Physiol. Plant. 1967, 20, 430–436. [Google Scholar] [CrossRef]
- Stadnik, S.A.; Falkova, T.V.; Yablonskij, E.A.; Chernyak, V.V. Influence of temperature on bioelectrical activity and physiological processes in plant in winter. Tr. GNBS 1974, 64, 99–105. (in Russian). [Google Scholar]
- Sawamura, Y.; Suesada, Y.; Sugiura, T.; Yaegaki, H. Chilling Requirements and Blooming Dates of Leading Peach Cultivars and a Promising Early Maturing Peach Selection, Momo Tsukuba 127. Hortic. J. 2017, 86, 426–436. [Google Scholar] [CrossRef]
- Legave, J.M.; Christen, D.; Giovannini, D.; Oger, R. Global warming in Europe and its impact on floral bud phenology in fruit species. Acta Hortic. 2009, 838, 21–26. [Google Scholar] [CrossRef]
- Kwon, J.; Nam, E.; Yun, S.; Kim, S.; Song, S.; Lee, J.; Hwang, K. Chilling and heat requirement of peach cultivars and changes in chilling accumulation spectrums based on 100-year records in Republic of Korea. Agric. For. Meteorol. 2020, 288–289, 108009. [Google Scholar] [CrossRef]
- Vuković, A.J.; Vujadinović, M.P.; Rendulić, S.M.; Đurđević, V.S.; Ruml, M.M.; Babić, V.P.; Popović, D.P. Global warming Impact on Climate Change in Serbia. Therm. Sci. 2018, 22, 2267–2280. [Google Scholar] [CrossRef]
- Alonso, J.M.; Ansón, J.M.; Espiau, M.T. R. Socias i Company. Determination of endodormancy break in almond flower buds by a correlation model using the average temperature of different day intervals and its application to the estimation of chill and heat requirements and blooming date. J. Am. Soc. Hortic. Sci. 2005, 130, 308–318. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Ben Mimoun, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J.; Luedeling, E. Cultivar-specific responses of sweet cherry flowering to rising temperatures during dormancy. Agric. For. Meteorol. 2021, 307, 108486. [Google Scholar] [CrossRef]
- Kitamura, Y.; Habu, T.; Yamane, H.; Nishiyama, S.; Kajita, K.; Sobue, T.; Kawai, T.; Numaguchi, K.; Nakazaki, T.; Kitajima, A.; et al. Identification of QTLs controlling chilling and heat requirements for dormancy release and bud break in Japanese apricot (Prunus mume). Tree Genet. Genomes 2018, 14, 33. [Google Scholar] [CrossRef]
- Abou-Saaid, O.; El Yaacoubi, A.; Moukhli, A.; El Bakkali, A.; Oulbi, S.; Delalande, M.; Farrera, I.; Kelner, J.-J.; Lochon-Menseau, S.; El Modafar, C.; et al. Statistical Approach to Assess Chill and Heat Requirements of Olive Tree Based on Flowering Date and Temperatures Data: Towards Selection of Adapted Cultivars to Global Warming. Agronomy 2022, 12, 2975. [Google Scholar] [CrossRef]
- Wypych, A.; Ustrnul, Z.; Sulikowska, A.; Chmielewski, F.-M.; Bochenek, B. Spatial and temporal variability of the frost-free season in Central Europe and its circulation background. Int. J. Climatol. 2017, 37, 3340–3352. [Google Scholar] [CrossRef]
- Chervenkov, H.; Slavov, K. Assessment of agrometeorological indices over Southeast Europe in the context of climate change (1961–2018). Idojaras 2021, 125, 255–269. [Google Scholar] [CrossRef]
- Cherif, S.; Doblas-Miranda, E.; Lionello, P.; Borrego, C.; Giorgi, F.; Iglesias, A.; Jebari, S.; Mahmoudi, E.; Moriondo, M.; Pringault, O.; et al. Drivers of change. In Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future. First Mediterranean Assessment Report; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UMEP/MAP: Marseille, France, 2020; pp. 59–180. [Google Scholar] [CrossRef]
- Meng, L.; Zhou, Y.; Gu, L.; Richardson, A.D.; Peñuelas, J.; Fu, Y.; Wang, Y.; Asrar, G.R.; De Boeck, H.J.; Mao, J.; et al. Photoperiod decelerates the advance of spring phenology of six deciduous tree species under climate warming. Glob. Change Biol. 2021, 27, 2914–2927. [Google Scholar] [CrossRef]
- Benmoussa, H.; Luedeling, E.; Ghrab, M.; Ben Mimoun, M. Severe winter chill decline impacts Tunisian fruit and nut orchards. Clim. Change 2020, 162, 1249–1267. [Google Scholar] [CrossRef]
- Rodrнguez, A.; Pйrez-Lypez, D.; Sбnchez, E.; Centeno, A.; Gymara, I.; Dosio, A.; Ruiz-Ramos, M. Chilling accumulation in fruit trees in Spain under climate change. Nat. Hazards Earth Syst. Sci. 2019, 19, 1087–1103. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Formula | Parameters to Be Defined |
|---|---|---|
| m1 | a, b, c,
| |
| m2 | , C*; | |
| GDD | ||
| GDDdoy | ||
| BCdoy | ||
| SIGdoy | ||
| SIGFOTOdoy | ||
| ChillBC doy (m1 + BCdoy) | | a, b, c, C*; |
| Phenological Model | |||||
|---|---|---|---|---|---|
| GDD | GDDdoy | BCdoy | SIGFOTOdoy | SIGdoy | ChillBCdoy |
| Parameters | |||||
| t1 = 31.Jan | t1 = 04.Feb | t1 = 27.Jan | t1 = 15.Jan | t1 = 02.Feb | t0 = 01.Nov |
| Tb = 0.0 | Tb = 0.3 | Tb = 1.2 | bf = −0.3685 | bf =−0.4291 | a =0.1394 |
| F* = 308.6 | F* = 277.1 | EXPO =1.578 | cf =8.9921 | cf =5.8505 | b =1.5034 |
| F* = 310.7 | EXPO =1.7541 | F* = 25.5 | c =11.2765 | ||
| F* = 21.1 | C* = 68.1 | ||||
| Tb = 2.6 | |||||
| EXPO =2.8603 | |||||
| F* = 261.1 | |||||
| Calibration data | |||||
| n = 20 | n = 20 | n = 20 | n = 20 | n = 20 | n = 20 |
| R2 = 0.91 | R2 = 0.91 | R2 = 0.92 | R2 = 0.91 | R2 = 0.90 | R2 = 0.92 |
| RMSE = 3.2 | RMSE = 3.2 | RMSE = 3.0 | RMSE = 3.3 | RMSE = 3.3 | RMSE = 3.0 |
| AICc = 48.78 | AICc = 48.47 | AICc = 46.61 | AICc = 51.64 | AICc = 50.74 | AICc = 52.24 |
| θ = −0.5 | θ = −0.1 | θ = 0.5 | θ = −0.4 | θ = 0.3 | θ = −0.4 |
| Validation data | |||||
| n = 8 | n = 8 | n = 8 | n = 8 | n = 8 | n = 8 |
| R2 = 0.78 | R2 = 0.79 | R2 = 0.83 | R2 = 0.82 | R2 = 0.78 | R2 = 0.83 |
| RMSE = 3.9 | RMSE = 3.8 | RMSE = 3.4 | RMSE = 3.5 | RMSE = 3.7 | RMSE = 3.4 |
| θ = 0.4 | θ = −0.8 | θ = 0.9 | θ = −0.6 | θ = 0.1 | θ = −0.6 |
| Model | Shapiro–Wilk Test | Breusch–Pagan Test | ||
|---|---|---|---|---|
| W | p-Value | BP | p-Value | |
| GDD | 0.97146 | 0.6201 | 0.8750 | 0.3496 |
| GDDdoy | 0.97562 | 0.7359 | 0.0226 | 0.8804 |
| BCdoy | 0.97095 | 0.6063 | 0.1388 | 0.7094 |
| SIGdoy | 0.97169 | 0.6266 | 1.8782 | 0.1763 |
| SIGFOTOdoy | 0.94023 | 0.1120 | 0.2212 | 0.6382 |
| ChillBCdoy | 0.97537 | 0.7289 | 1.1889 | 0.2756 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korsakova, S.; Korzin, V.; Plugatar, Y.; Kazak, A.; Gorina, V.; Korzina, N.; Khokhlov, S.; Makoveichuk, K. Modelling of Climate Change’s Impact on Prunus armeniaca L.’s Flowering Time. Inventions 2023, 8, 65. https://doi.org/10.3390/inventions8030065
Korsakova S, Korzin V, Plugatar Y, Kazak A, Gorina V, Korzina N, Khokhlov S, Makoveichuk K. Modelling of Climate Change’s Impact on Prunus armeniaca L.’s Flowering Time. Inventions. 2023; 8(3):65. https://doi.org/10.3390/inventions8030065
Chicago/Turabian StyleKorsakova, Svetlana, Vadim Korzin, Yuri Plugatar, Anatoliy Kazak, Valentina Gorina, Natalia Korzina, Sergey Khokhlov, and Krystina Makoveichuk. 2023. "Modelling of Climate Change’s Impact on Prunus armeniaca L.’s Flowering Time" Inventions 8, no. 3: 65. https://doi.org/10.3390/inventions8030065
APA StyleKorsakova, S., Korzin, V., Plugatar, Y., Kazak, A., Gorina, V., Korzina, N., Khokhlov, S., & Makoveichuk, K. (2023). Modelling of Climate Change’s Impact on Prunus armeniaca L.’s Flowering Time. Inventions, 8(3), 65. https://doi.org/10.3390/inventions8030065