
The Relationship between Training Cycle-Dependent Fluctuations in Resting Blood Lactate Levels and Exercise Performance in College-Aged Rugby Players


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Design
2.3. Exercise Performance Measurement
2.4. Body Composition
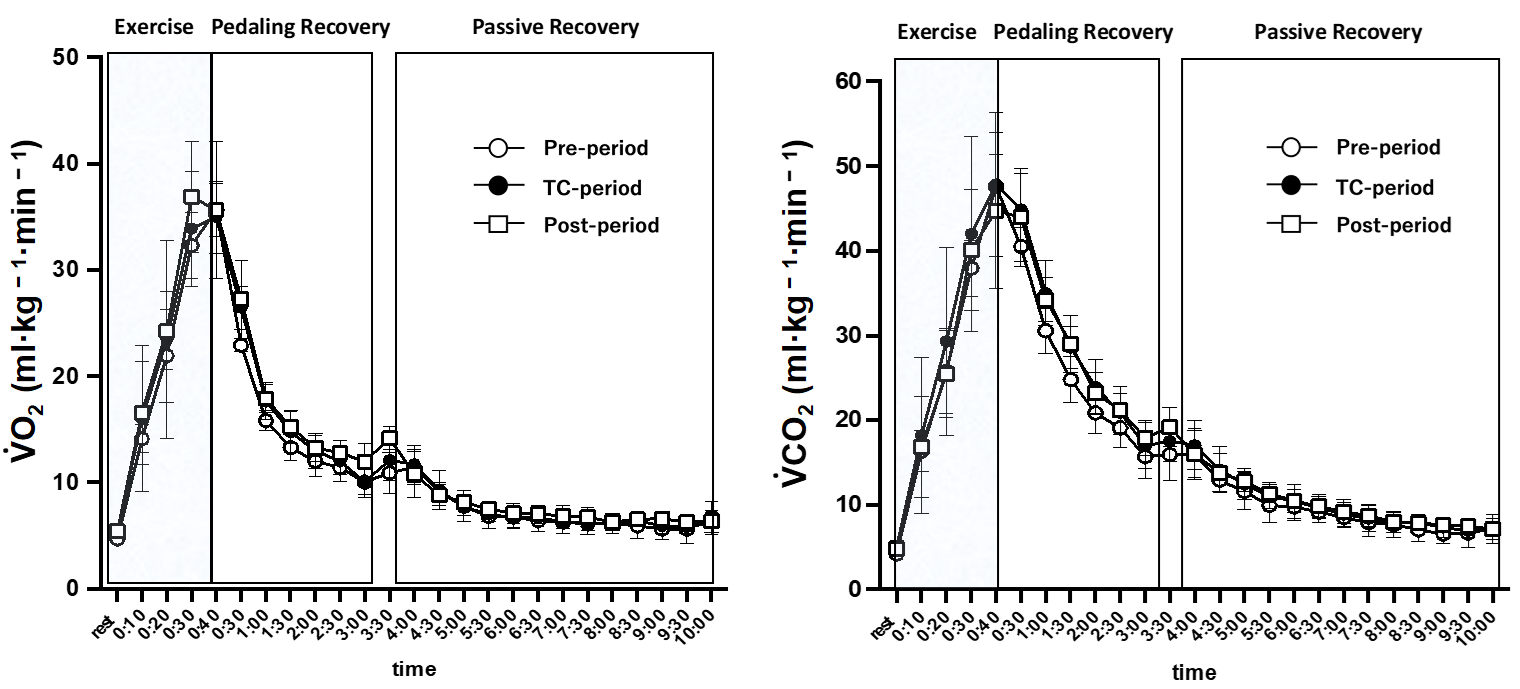
2.5. Respiratory Measurements
2.6. Calorie Calculation
2.7. Statistical Analysis
3. Results
3.1. Whole Body Composition
3.2. Resting Blood Lactate and Glucose Levels in the Morning (after Waking Up)
3.3. Analysis of the 40 s Wingate Anaerobic Test (40-s WT)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Broskey, N.T.; Zou, K.; Dohm, G.L.; Houmard, J.A. Plasma Lactate as a Marker for Metabolic Health. Exerc. Sport Sci. Rev. 2020, 48, 119–124. [Google Scholar] [CrossRef]
- Barres, R.; Osler, M.E.; Yan, J.; Rune, A.; Fritz, T.; Caidahl, K.; Krook, A.; Zierath, J.R. Non-CpG methylation of the PGC-1alpha promoter through DNMT3B controls mitochondrial density. Cell Metab. 2009, 10, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Avogaro, A.; Toffolo, G.; Miola, M.; Valerio, A.; Tiengo, A.; Cobelli, C.; Del Prato, S. Intracellular lactate- and pyruvate-interconversion rates are increased in muscle tissue of non-insulin-dependent diabetic individuals. J. Clin. Investig. 1996, 98, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Juraschek, S.P.; Selvin, E.; Miller, E.R.; Brancati, F.L.; Young, J.H. Plasma lactate and diabetes risk in 8045 participants of the atherosclerosis risk in communities study. Ann. Epidemiol. 2013, 23, 791–796. [Google Scholar] [CrossRef] [Green Version]
- Gollnick, P.D.; Bayly, W.M.; Hodgson, D.R. Exercise intensity, training, diet, and lactate concentration in muscle and blood. Med. Sci. Sports Exerc. 1986, 18, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Beneke, R.; Leithauser, R.M.; Ochentel, O. Blood lactate diagnostics in exercise testing and training. Int. J. Sports Physiol. Perform. 2011, 6, 8–24. [Google Scholar] [CrossRef] [Green Version]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [Green Version]
- Dubouchaud, H.; Butterfield, G.E.; Wolfel, E.E.; Bergman, B.C.; Brooks, G.A. Endurance training, expression, and physiology of LDH, MCT1, and MCT4 in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E571–E579. [Google Scholar] [CrossRef] [Green Version]
- Bergman, Bduring rest and exercise in men. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E244–E251. [CrossRef] [Green Version]
- Burgomaster, K.A.; Cermak, N.M.; Phillips, S.M.; Benton, C.R.; Bonen, A.; Gibala, M.J. Divergent response of metabolite transport proteins in human skeletal muscle after sprint interval training and detraining. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1970–R1976. [Google Scholar] [CrossRef]
- Fiorenza, M.; Gunnarsson, T.P.; Hostrup, M.; Iaia, F.M.; Schena, F.; Pilegaard, H.; Bangsbo, J. Metabolic stress-dependent regulation of the mitochondrial biogenic molecular response to high-intensity exercise in human skeletal muscle. J. Physiol. 2018, 596, 2823–2840. [Google Scholar] [CrossRef] [Green Version]
- Burgomaster, K.A.; Hughes, S.C.; Heigenhauser, G.J.; Bradwell, S.N.; Gibala, M.J. Six sessions of sprint interval training increases muscle oxidative potential and cycle endurance capacity in humans. J. Appl. Physiol. 2005, 98, 1985–1990. [Google Scholar] [CrossRef]
- Hashimoto, T.; Hussien, R.; Oommen, S.; Gohil, K.; Brooks, G.A. Lactate sensitive transcription factor network in L6 cells: Activation of MCT1 and mitochondrial biogenesis. FASEB J. 2007, 21, 2602–2612. [Google Scholar] [CrossRef]
- Kano, R.; Sato, K. A Competitive sprinter’s resting blood lactate levels fluctuate with a one-year training cycle: Case Reports. J. Funct. Morphol. Kinesiol. 2021, 6, 95. [Google Scholar] [CrossRef]
- Thomas, C.; Perrey, S.; Lambert, K.; Hugon, G.; Mornet, D.; Mercier, J. Monocarboxylate transporters, blood lactate removal after supramaximal exercise, and fatigue indexes in humans. J. Appl. Physiol. 2005, 98, 804–809. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Bishop, D.J.; Lambert, K.; Mercier, J.; Brooks, G.A. Effects of acute and chronic exercise on sarcolemmal MCT1 and MCT4 contents in human skeletal muscles: Current status. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R1–R14. [Google Scholar] [CrossRef] [Green Version]
- Messonnier, L.A.; Emhoff, C.A.; Fattor, J.A.; Horning, M.A.; Carlson, T.J.; Brooks, G.A. Lactate kinetics at the lactate threshold in trained and untrained men. J. Appl. Physiol. 2013, 114, 1593–1602. [Google Scholar] [CrossRef]
- Poole, D.C.; Rossiter, H.B.; Brooks, G.A.; Gladden, L.B. The anaerobic threshold: 50+ years of controversy. J. Physiol. 2021, 599, 737–767. [Google Scholar] [CrossRef]
- Bergman, B.C.; Wolfel, E.E.; Butterfield, G.E.; Lopaschuk, G.D.; Casazza, G.A.; Horning, M.A.; Brooks, G.A. Active muscle and whole body lactate kinetics after endurance training in men. J. Appl. Physiol. 1999, 87, 1684–1696. [Google Scholar] [CrossRef] [Green Version]
- San-Millan, I.; Brooks, G.A. Assessment of metabolic flexibility by means of measuring blood lactate, fat, and carbohydrate oxidation responses to exercise in professional endurance athletes and less-fit individuals. Sports Med. 2018, 48, 467–479. [Google Scholar] [CrossRef]
- Mujika, I.; Padilla, S. Detraining: Loss of training-induced physiological and performance adaptations. Part II: Long term insufficient training stimulus. Sports Med. 2000, 30, 145–154. [Google Scholar] [CrossRef]
- Wibom, R.; Hultman, E.; Johansson, M.; Matherei, K.; Constantin-Teodosiu, D.; Schantz, P.G. Adaptation of mitochondrial ATP production in human skeletal muscle to endurance training and detraining. J. Appl. Physiol. 1992, 73, 2004–2010. [Google Scholar] [CrossRef]
- Coyle, E.F.; Martin, W.H., 3rd; Sinacore, D.R.; Joyner, M.J.; Hagberg, J.M.; Holloszy, J.O. Time course of loss of adaptations after stopping prolonged intense endurance training. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1984, 57, 1857–1864. [Google Scholar] [CrossRef]
- Coyle, E.F.; Martin, W.H., 3rd; Bloomfield, S.A.; Lowry, O.H.; Holloszy, J.O. Effects of detraining on responses to submaximal exercise. J. Appl. Physiol. 1985, 59, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.; Reidy, P.T.; Bhattarai, N.; Sidossis, L.S.; Rasmussen, B.B. Resistance exercise training alters mitochondrial function in human skeletal muscle. Med. Sci. Sports Exerc. 2015, 47, 1922–1931. [Google Scholar] [CrossRef] [Green Version]
- Evertsen, F.; Medbo, J.I.; Bonen, A. Effect of training intensity on muscle lactate transporters and lactate threshold of cross-country skiers. Acta Physiol. Scand. 2001, 173, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.W.; Mackenhauer, J.; Roberts, J.C.; Berg, K.M.; Cocchi, M.N.; Donnino, M.W. Etiology and therapeutic approach to elevated lactate levels. Mayo Clin. Proc. 2013, 88, 1127–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzeri, C.; Valente, S.; Chiostri, M.; Gensini, G.F. Clinical significance of lactate in acute cardiac patients. World J. Cardiol. 2015, 7, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.E.; Pories, W.J.; Houmard, J.A.; Tanner, C.J.; Zheng, D.; Zou, K.; Coen, P.M.; Goodpaster, B.H.; Kraus, W.E.; Dohm, G.L. Plasma lactate as a marker of metabolic health: Implications of elevated lactate for impairment of aerobic metabolism in the metabolic syndrome. Surgery 2019, 166, 861–866. [Google Scholar] [CrossRef]
- Broskey, N.T.; Pories, W.J.; Jones, T.E.; Tanner, C.J.; Zheng, D.; Cortright, R.N.; Yang, Z.W.; Khang, N.; Yang, J.; Houmard, J.A.; et al. The association between lactate and muscle aerobic substrate oxidation: Is lactate an early marker for metabolic disease in healthy subjects? Physiol. Rep. 2021, 9, e14729. [Google Scholar] [CrossRef]
- McKenna, M.J.; Heigenhauser, G.J.; McKelvie, R.S.; Obminski, G.; MacDougall, J.D.; Jones, N.L. Enhanced pulmonary and active skeletal muscle gas exchange during intense exercise after sprint training in men. J. Physiol. 1997, 501 Pt 3, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Souissi, N.; Bessot, N.; Chamari, K.; Gauthier, A.; Sesboue, B.; Davenne, D. Effect of time of day on aerobic contribution to the 30-s Wingate test performance. Chronobiol. Int. 2007, 24, 739–748. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Period | TC-Period | Post-Period | |
|---|---|---|---|
| Body Weight (kg) | 77.8 ± 8.9 | 78.6 ± 10.0 | 77.8 ± 10.9 |
| Muscle Mass (kg) | 34.4 ± 4.2 | 35.2 ± 4.4 * | 34.2 ± 4.3 |
| Body Fat (%) | 22.3 ± 3.6 | 21.5 ± 4.1 * | 22.7 ± 4.4 |
| Calorie Intake (kcal/day) | 2426.3 ± 443.8 | 2543.1 ± 431.4 | 2657.1 ± 454.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kano, R.; Sato, K. The Relationship between Training Cycle-Dependent Fluctuations in Resting Blood Lactate Levels and Exercise Performance in College-Aged Rugby Players. J. Funct. Morphol. Kinesiol. 2022, 7, 93. https://doi.org/10.3390/jfmk7040093
Kano R, Sato K. The Relationship between Training Cycle-Dependent Fluctuations in Resting Blood Lactate Levels and Exercise Performance in College-Aged Rugby Players. Journal of Functional Morphology and Kinesiology. 2022; 7(4):93. https://doi.org/10.3390/jfmk7040093
Chicago/Turabian StyleKano, Ryotaro, and Kohei Sato. 2022. "The Relationship between Training Cycle-Dependent Fluctuations in Resting Blood Lactate Levels and Exercise Performance in College-Aged Rugby Players" Journal of Functional Morphology and Kinesiology 7, no. 4: 93. https://doi.org/10.3390/jfmk7040093
APA StyleKano, R., & Sato, K. (2022). The Relationship between Training Cycle-Dependent Fluctuations in Resting Blood Lactate Levels and Exercise Performance in College-Aged Rugby Players. Journal of Functional Morphology and Kinesiology, 7(4), 93. https://doi.org/10.3390/jfmk7040093