
Epidemiology of Rabies in Lesotho: The Importance of Routine Surveillance and Virus Characterization

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diagnostic Training and Sample Cohort
2.2. Direct, Rapid Immunohistochemical Test (DRIT)
2.3. Direct Fluorescent Antibody (DFA) Test
2.4. Resolving Diagnostic Incongruities
2.5. Data Analysis
2.6. Molecular Epidemiology of Rabies in LESOTHO
2.6.1. Sample Cohort
2.6.2. Viral RNA Extractions, PCR, and Sequencing
2.6.3. Phylogenetic Analysis
3. Results
3.1. Statistical Analysis of DRIT Diagnostic Efficacy
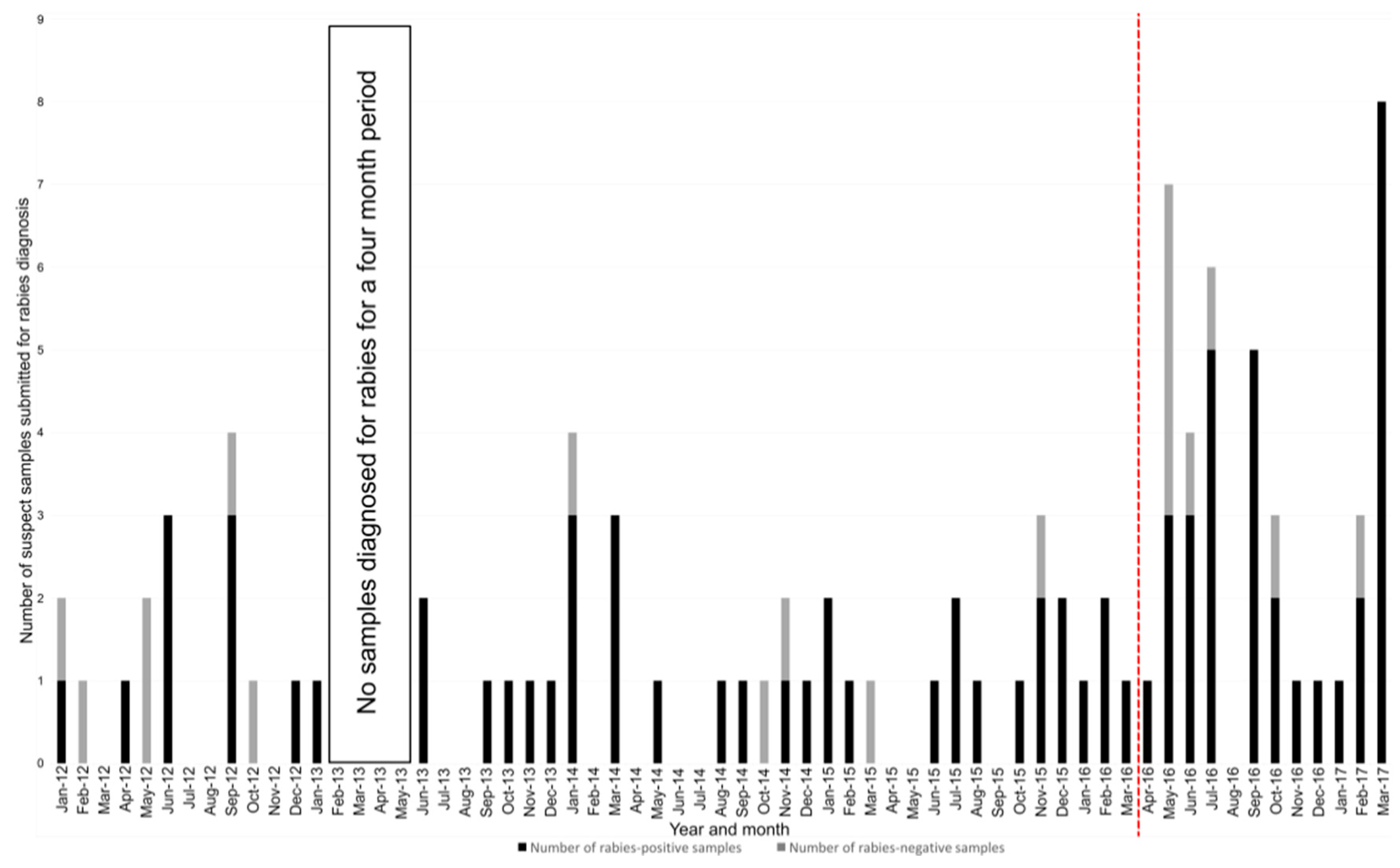
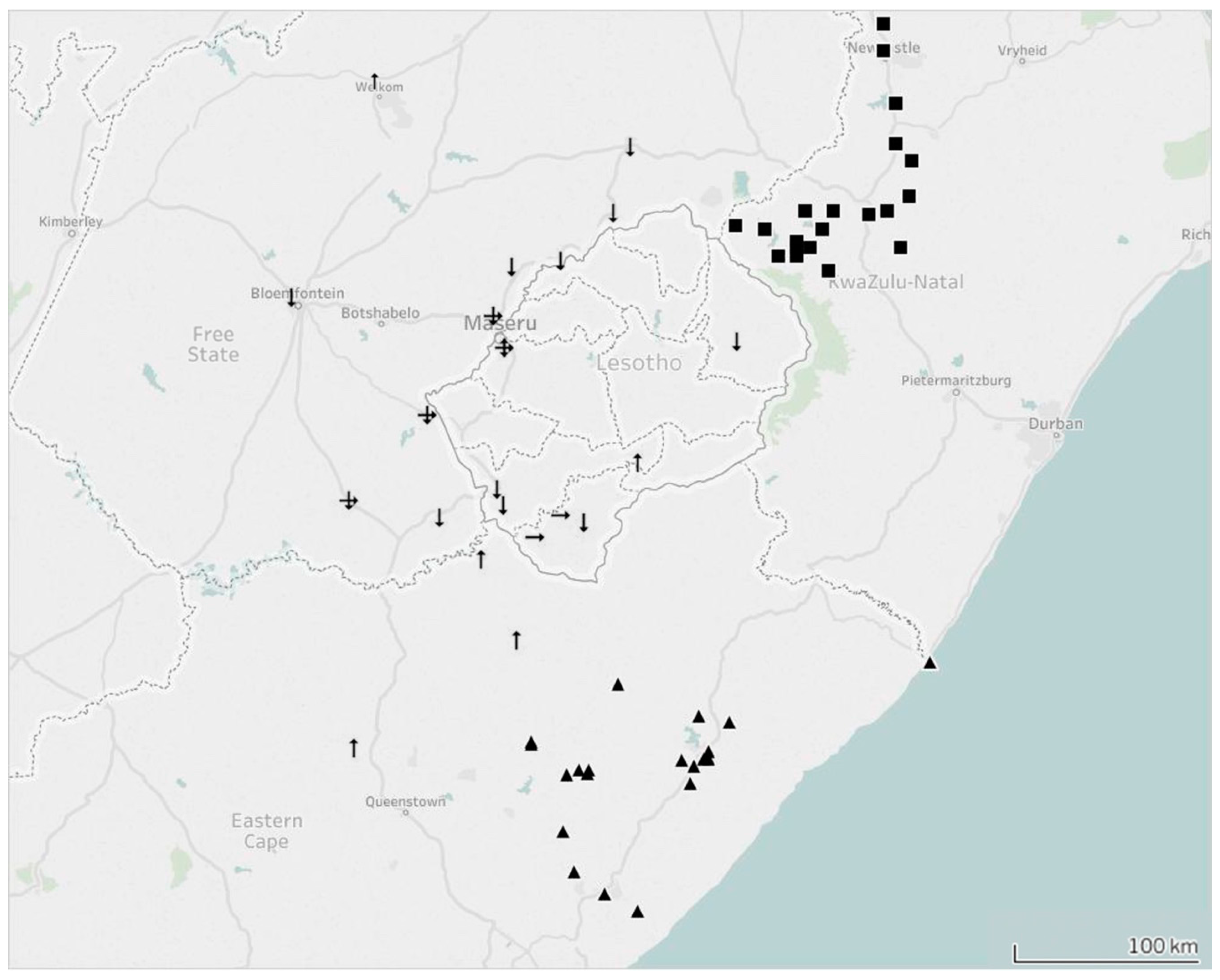
3.2. Increased Sample Submission and Distribution Analysis
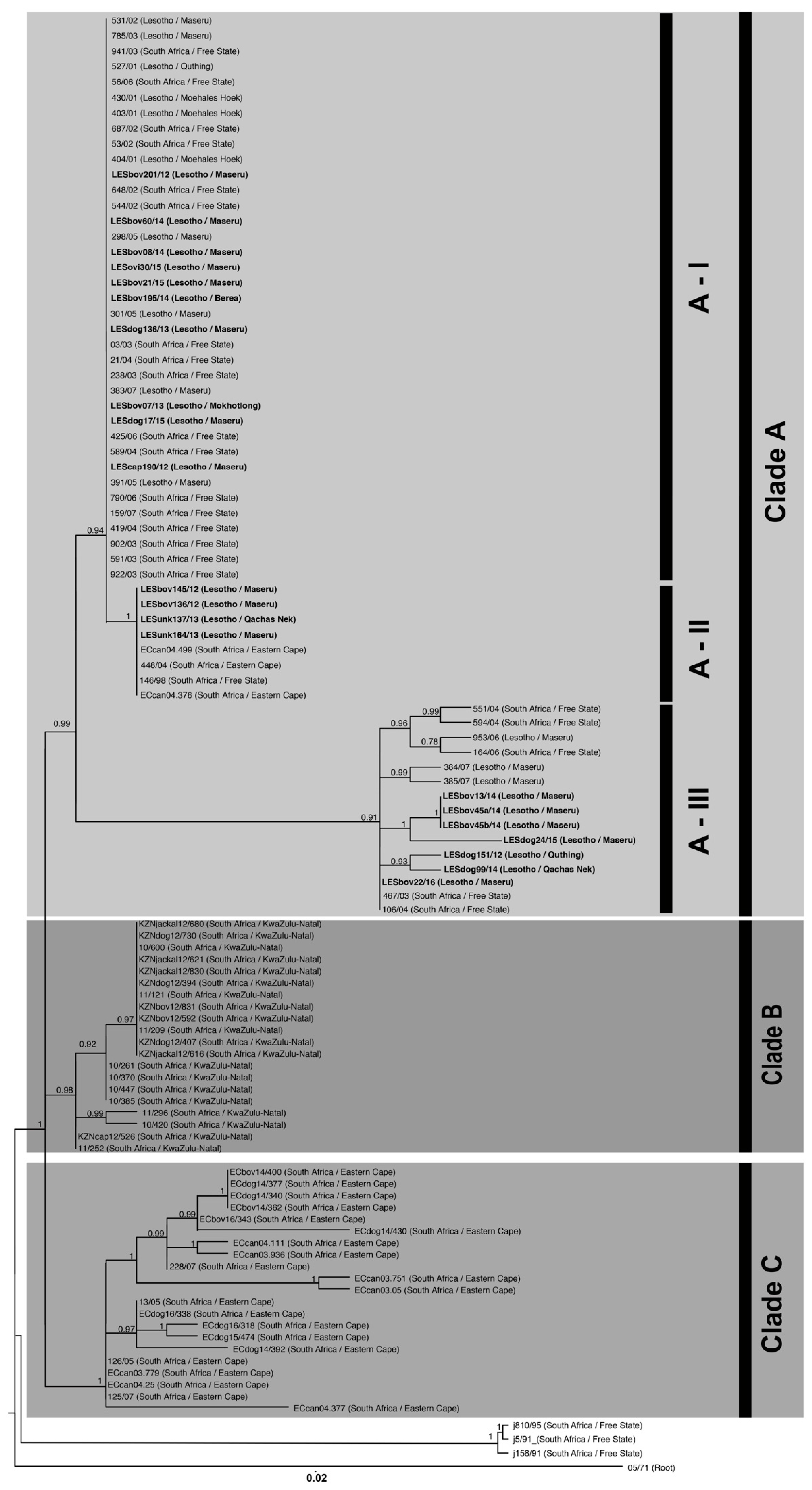
3.3. Molecular Epidemiology
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the global burden of endemic canine rabies. PLoS Negl. Trop. Dis. 2015, 9. [Google Scholar] [CrossRef] [Green Version]
- Coleman, P.G.; Fevre, E.M.; Cleaveland, S. Estimating the public health impact of rabies. Emerg. Infect. Dis. 2004, 10, 140–142. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). WHO Expert Consultation on Rabies: Second Report; World Health Organization: Geneva, Switzerland, 2013; pp. 1–141. [Google Scholar]
- Taylor, L.H.; Hampson, K.; Fahrion, A.; Abela-Ridder, B.; Nel, L.H. Difficulties in estimating the human burden of canine rabies. Acta Trop. 2015, 165, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R.; Barnard, B.J.H.; Meredith, C.D.; Bishop, G.C.; Bruckner, G.K.; Foggin, C.M.; Hübschle, O.J. Rabies in southern Africa. Onderstepoort J. Vet. Res. 1993, 60, 325–346. [Google Scholar] [PubMed]
- Khomari, L. Rabies in Lesotho. In Proceedings of the international conference on epidemiology, control and prevention of rabies in eastern and southern Africa, Lusaka, Zambia, 2–5 June 1992; pp. 45–46. [Google Scholar]
- Scott, C. Rabies in Lesotho. Vet. Rec. 1988, 122. [Google Scholar] [CrossRef]
- Makalo, M.J.R.; Khanyane, M.; Maseko, L. PARACON Country Report: Lesotho (Available on request from the PARACON Steering Committee). Johannesburg, South Africa, 2015. Available online: https://rabiesalliance.org/contact-us?src=PARACON (accessed on July 2015).
- Ngoepe, C.E.; Sabeta, C.; Nel, L. The spread of canine rabies into Free State Province of South Africa: A molecular epidemiological characterization. Virus Res. 2009, 142, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Bishop, G.C.; Durrheim, D.N.; Kloeck, P.E.; Godlonton, J.D.; Bingham, J.; Speare, R.; Rabies Advisory Group 2000. Rabies: Guide for the Medical, Veterinary and allied Professions, 2nd ed.; Department of Agriculture and Department of Health: Pretoria, South Africa, 2010.
- Davis, P.L.; Rambaut, A.; Bourhy, H.; Holmes, E.C. The evolutionary dynamics of canid and mongoose rabies virus in southern Africa. Arch. Virol. 2007, 152, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Coetzer, A.; Anahory, I.; Dias, P.T.; Sabeta, C.T.; Scott, T.P.; Markotter, W.; Nel, L.H. Enhanced diagnosis of rabies and molecular evidence for the transboundary spread of the disease in Mozambique. J. S. Afr. Vet. Assoc. 2017, 88. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.J.; Abelseth, M.K.; Atanasiu, P. The Fluorescent Antibody Test. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.-X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 88–89. [Google Scholar]
- Barrat, J. Simple technique for the collection and shipment of brain specimens for rabies diagnosis. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.-X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 425–432. [Google Scholar]
- Coetzer, A. Comparison of Biotinylated Monoclonal and Polyclonal Antibodies in an Evaluation of a Direct Rapid Immunohistochemical Test for the Routine Diagnosis of Rabies in Southern Africa. Master’s Thesis, University of Pretoria, Pretoria, South Africa, June 2013. [Google Scholar]
- Coetzer, A.; Nel, L.H.; Rupprecht, C.E. Demonstration of Lyssavirus antigens by a direct rapid immunohistochemical test. In Current Laboratory Techniques in Rabies Diagnosis, Research and Prevention, 1st ed.; Rupprecht, C.E., Nagarajan, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 27–36. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Protocol for Postmortem Diagnosis of Rabies in Animals by Direct Fluorescent Antibody Testing. 2003. Available online: https://www.cdc.gov/rabies/pdf/rabiesdfaspv2.pdf (accessed on 27 September 2016).
- Coertse, J.; Weyer, J.; Nel, L.H.; Markotter, W. Improved PCR methods for detection of African rabies and rabies-related lyssaviruses. J. Clin. Microbiol. 2010, 48, 3949–3955. [Google Scholar] [CrossRef] [PubMed]
- Sacramento, D.; Bourhy, H.; Tordo, N. PCR technique as an alternative method for diagnosis and molecular epidemiology of rabies virus. Mol. Cell. Probes 1991, 5, 229–240. [Google Scholar] [CrossRef]
- Von Teichman, B.F.; Thomson, G.R.; Meredith, C.D.; Nel, L.H. Molecular epidemiology of rabies virus in South Africa: Evidence for two distinct virus groups. J. Gen. Virol. 1995, 76, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symp. 1999, 41, 95–98. [Google Scholar]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- KwaZulu-Natal Department of the Agriculture Rural Development (KZNDARD). Jackal Rabies in the KwaZulu-Natal Midlands; 2015. Available online: http://www.kzndard.gov.za/images/Documents/AnimalHealth/AnimalDiseases/Jackal-Rabies-in-the-KwaZulu-Natal-Midlands.pdf (accessed on 18 July 2017).
- Durr, S.; Naissengar, S.; Mindekem, R.; Diguimbye, C.; Niezgoda, M.; Kuzmin, I.; Rupprecht, C.E.; Zinsstag, J. Rabies diagnosis for developing countries. PLoS Negl. Trop. Dis. 2008, 2. [Google Scholar] [CrossRef] [PubMed]
- Weyer, J.; Blumberg, L. Rabies: Challenge of diagnosis in resource poor countries. Infect Dis. J. Pak. 2007, 16, 86–88. [Google Scholar]
- McElhinney, L.M.; Marston, D.A.; Brookes, S.M.; Fooks, A.R. Effects of carcase decomposition on rabies virus infectivity and detection. J. Virol. Methods 2014, 207, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Mani, R.S.; Madhusudana, S.N. Laboratory diagnosis of human rabies: Recent advances. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Rojas Anaya, E.; Loza-Rubio, E.; Banda Ruiz, V.M.; Hernández Baumgarten, E. Use of reverse transcription-polymerase chain reaction to determine the stability of rabies virus genome in brains kept at room temperature. J. Vet. Diagn. Investig. 2006, 18, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.C.; Venditti, L.L.R.; Queiroz, L.H. Comparison between RT-PCR and the mouse inoculation test for detection of rabies virus in samples kept for long periods under different conditions. J. Virol. Methods 2010, 164, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, T.D.; Teixeira, M.F.; Costa, E.C.; Vitaliano, A.B.; Teles, C.H.; Barroso, I.C.; Dias, R.P.; Moraes, N.B. Medium-term cryopreservation of rabies virus samples. Rev. Soc. Bras. Med. Trop. 2013, 46, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Lembo, T.; Niezgoda, M.; Velasco-Villa, A.; Cleaveland, S.; Ernest, E.; Rupprecht, C.E. Evaluation of a direct, rapid immunohistochemical test for rabies diagnosis. Emerg. Infect. Dis. 2006, 12, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.E.; Cliquet, F.; Fehlner-Gardiner, C.; Fooks, A.R.; Mueller, T.; Sabeta, C.; Slate, D. Progress in the development of a direct rapid immunohistochemical test for diagnosing rabies. News Colleagues 2014, 3, 87–95. [Google Scholar]
- Coetzer, A.; Sebeta, C.T.; Markotter, W.; Rupprecht, C.E.; Nel, L.H. Comparison of biotinylated monoclonal and polyclonal antibodies in an evaluation of a direct rapid immunohistochemical test for the routine diagnosis of rabies in southern Africa. PLoS Negl. Trop. Dis. 2014, 9. [Google Scholar] [CrossRef]
- Mkhize, G.C.; Ngoepe, E.C.; Du Plessis, B.J.; Reininghaus, B.; Sabeta, C.T. Re-emergence of dog rabies in Mpumalanga Province, South Africa. Vector-Borne Zoonotic Dis. 2010, 10, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C. The mammals of Lesotho. Res. Natl. Museum. Bloemfontein. 1994, 10, 171–241. [Google Scholar]
- Rowe-Rowe, D.T. Black-backed jackal diet in relation to food availability in the Natal Drakensberg. S. Afr. J. Wildl. Res. 1983, 13, 17–23. [Google Scholar]
- Lesotho; Ministry of Local Government and Chieftainship. Quthing District Council: Local Government District Information Handbook: A Compilation of Crucial Information for the District, 1st ed.; June 2008. Available online: https://www.giz.de/en/downloads/en-lesotho-quthing-information-handbook.pdf (accessed on June 2008).
- Day, M.J. One health: The importance of companion animal vector-borne diseases. Parasites Vectors 2011, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; Banyard, A.C.; Horton, D.L.; Johnson, N.; McElhinney, L.M.; Jackson, A.C. Current status of rabies and prospects for elimination. Lancet 2014, 384, 1389–1399. [Google Scholar] [CrossRef]
- Russel, C.A.; Real, L.A.; Smith, D.L. Spatial control of rabies on heterogeneous landscapes. PLoS ONE 2006, 1, e27. [Google Scholar] [CrossRef] [PubMed]
- United Nations Department of Economic and Social Affairs; United Nations Department of Economic and Social Affairs: New York, NY, USA, 2014.




{kind=link}
{kind=link}
{kind=link}
| Sample Number | Species | Town | DRIT Result | DFA Result | Real-Time PCR Result | |
|---|---|---|---|---|---|---|
| 1 * | 15/09/11 | Bovine | Maseru | Positive | Positive | --- |
| 2 * | 254/09/11 | Bovine | Mokhotlong | Negative | Negative | --- |
| 3 * | 11/01/12 | Bovine | Berea | Positive | Positive | --- |
| 4 * | 24/01/12 | Bovine | Maseru | Negative | Negative | --- |
| 5 * | 04/02/12 | Bovine | Maseru | Negative | Negative | --- |
| 6 * | 106/04/12 | Bovine | Maseru | Negative | Negative | --- |
| 7 * | 113/05/12 | Bovine | Maseru | Negative | Negative | --- |
| 8 * | 123/05/12 | Canine | Maseru | Negative | Negative | --- |
| 9 * | 14/06/12 | Canine | Maseru | Positive | Positive | --- |
| 10 *,# | 145/06/12 | Bovine | Maseru | Positive | Positive | --- |
| 11 *,# | 136/06/12 | Canine | Maseru | Positive | Positive | --- |
| 12 * | 12/09/12 | Bovine | Maseru | Negative | Negative | --- |
| 13 *,# | 151/09/12 | Bovine | Quthing | Positive | Positive | --- |
| 14 *,# | 190/09/12 | Canine | Maseru | Positive | Positive | --- |
| 15 *,# | 201/09/12 | Caprine | Maseru | Positive | Positive | --- |
| 16 * | 211/10/12 | Canine | Maseru | Positive | Positive | --- |
| 17 * | 276/12/12 | Bovine | Maseru | Positive | Positive | --- |
| 18 *,# | 07/01/13 | Bovine | Mokhotlong | Positive | Positive | --- |
| 19 *,# | 136/06/13 | Bovine | Maseru | Positive | Positive | --- |
| 20 *,# | 137/06/13 | Canine | Qacha’s Nek | Positive | Positive | --- |
| 21 *,# | 164/09/13 | Canine | Maseru | Positive | Positive | --- |
| 22 * | 10/10/13 | Bovine | Berea | Positive | Positive | --- |
| 23 * | 05/11/13 | Bovine | Berea | Positive | Positive | --- |
| 24 *,# | 194/12/13 | Canine | Maseru | Positive | Positive | --- |
| 25 *,# | 08/01/14 | Bovine | Maseru | Positive | Positive | --- |
| 26 * | 10/01/14 | Bovine | Maseru | Negative | Negative | --- |
| 27 *,# | 13/01/14 | Bovine | Maseru | Positive | Positive | --- |
| 28 * | 23/01/14 | Canine | Maseru | Positive | Positive | --- |
| 29 * | 20/03/14 | Bovine | Maseru | Positive | Positive | --- |
| 30 *,# | 45a/03/14 | Bovine | Maseru | Positive | Positive | --- |
| 31 *,# | 45b/03/14 | Bovine | Maseru | Positive | Positive | --- |
| 32 *,# | 60/05/14 | Canine | Qacha’s Nek | Positive | Positive | --- |
| 33 * | 26/08/14 | Bovine | Maseru | Positive | Positive | --- |
| 34 *,# | 99/09/14 | Bovine | Maseru | Positive | Positive | --- |
| 35 * | 21/10/14 | Canine | Maseru | Negative | Negative | --- |
| 36 * | 192/11/14 | Bovine | Berea | Negative | Negative | --- |
| 37 * | 193/11/14 | Canine | Qacha’s Nek | Positive | Positive | --- |
| 38 * | 10/12/14 | Equine | Berea | Negative ° | Positive ° | Negative |
| 39 * | 07/01/15 | Canine | Maseru | Positive | Positive | --- |
| 40 *,# | 30/01/15 | Ovine | Maseru | Positive | Positive | --- |
| 41 *,# | 17/02/15 | Canine | Maseru | Positive | Positive | --- |
| 42 * | 18/03/15 | Feline | Maseru | Negative | Negative | --- |
| 43 * | 18/06/15 | Ovine | Qacha’s Nek | Positive | Positive | --- |
| 44 * | 150/11/15 | Canine | Maseru | Positive | Positive | --- |
| 45 *,# | 21/07/15 | Bovine | Maseru | Positive | Positive | --- |
| 46 * | 23/07/15 | Bovine | Maseru | Positive | Positive | --- |
| 47 *,# | 24/08/15 | Canine | Maseru | Positive | Positive | --- |
| 48 * | 06/10/15 | Canine | Maseru | Positive | Positive | --- |
| 49 * | 10/11/15 | Caprine | Maseru | Positive | Positive | --- |
| 50 * | 26/11/15 | Bovine | Mohales Hoek | Negative ° | Positive ° | Negative |
| 51 * | 149/11/15 | Caprine | Maseru | Positive | Positive | --- |
| 52 * | 29/12/15 | Canine | Maseru | Negative ° | Positive ° | Negative |
| 53 * | 161/12/15 | Canine | Berea | Positive | Positive | --- |
| 54 * | 14/01/16 | Canine | Maseru | Negative ° | Positive ° | Negative |
| 55 *# | 22/02/16 | Bovine | Maseru | Positive | Positive | --- |
| 56 * | 26/02/16 | Bovine | Berea | Negative ° | Positive ° | Negative |
| 57 * | 03/03/16 | Canine | Maseru | Negative ° | Positive ° | Negative |
| 58 + | 95/04/16 | Bovine | Berea | Positive | Positive | --- |
| 59 + | 100/05/2016 | Canine | Maseru | Negative | Negative | --- |
| 60 + | 105/05/2016 | Canine | Berea | Negative | Negative | --- |
| 61 + | 109/05/2016 | Canine | Maseru | Positive | Positive | --- |
| 62 + | 110/05/2016 | Canine | Berea | Positive | Positive | --- |
| 63 + | 113/05/2016 | Ovine | Berea | Negative | Negative | --- |
| 64 + | 115/05/2016 | Bovine | Berea | Positive | Positive | --- |
| 65 + | 123/6/2016 | Canine | Maseru | Positive | Positive | --- |
| 66 + | 125/6/2016 | Ovine | Maseru | Positive | Positive | --- |
| 67 + | 127/06/2016 | Porcine | Mohales Hoek | Negative | Negative | --- |
| 68 + | 128/06/2016 | Canine | Mohales Hoek | Positive | Positive | --- |
| 69 + | 131/07/2016 | Canine | Berea | Positive | Positive | --- |
| 70 + | 132/07/2016 | Canine | Maseru | Positive | Positive | --- |
| 71 + | 133/07/2016 | Canine | Maseru | Positive | Positive | --- |
| 72 + | 136/07/2016 | equine | Maseru | Negative | Negative | --- |
| 73 + | 137/07/2016 | Canine | Mohales Hoek | Positive | Positive | --- |
| 74 + | 138/07/2016 | Bovine | Mohales Hoek | Positive | Positive | --- |
| 75 + | 163/09/2016 | Ovine | Maseru | Positive | Positive | --- |
| 76 + | 179/09/2016 | Bovine | Maseru | Positive | Positive | --- |
| 77 + | 182/09/2016 | Canine | Mohales Hoek | Positive | Positive | --- |
| 78 + | 184/09/2016 | Canine | Maseru | Positive | Positive | --- |
| 79 + | 185/09/2016 | Canine | Maseru | Positive | Positive | --- |
| 80 + | 196/10/2016 | Canine | Berea | Positive | Positive | --- |
| 81 + | 198/10/2016 | Ovine | Maseru | Negative | Negative | --- |
| 82 + | 199/10/2016 | Bovine | Maseru | Positive | Positive | --- |
| 83 + | 210/11/2016 | Canine | Berea | Positive | Positive | --- |
| 84 + | 234/12/2016 | Canine | Maseru | Positive | Positive | --- |
| 85 + | 30/01/2017 | Bovine | Mafeteng | Positive | Positive | --- |
| 86 + | 62/02/2017 | Bovine | Maseru | Positive | Positive | --- |
| 87 + | 63/02/2017 | Canine | Maseru | Positive | Positive | --- |
| 88 + | 71/02/2017 | Bovine | Quthing | Negative | Negative | --- |
| 89 + | 63/03/2017 | Ovine | Maseru | Positive | Positive | --- |
| 90 + | 69/03/2017 | Bovine | Maseru | Positive | Positive | --- |
| 91 + | 70/03/2017 | Equine | Maseru | Positive | Positive | --- |
| 92 + | 72/03/2017 | Canine | Maseru | Positive | Positive | --- |
| 93 + | 88/03/2017 | Canine | Mokhotlong | Positive | Positive | --- |
| 94 + | 89/03/2017 | Bovine | Mokhotlong | Positive | Positive | --- |
| 95 + | 90/03/2017 | Canine | Mokhotlong | Positive | Positive | --- |
| 96 + | 91/03/2017 | Ovine | Mokhotlong | Positive | Positive | --- |
| True Positive | False Positive | True Negative | False Negative | Diagnostic Sensitivity * | Diagnostic Specificity * | |
|---|---|---|---|---|---|---|
| DFA | 72 | 6 | 18 | 0 | 100% (95.01–100%) | 75.00% (53.29–90.23%) |
| DRIT | 72 | 0 | 24 | 0 | 100% (95.01–100%) | 100% (85.75–100%) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coetzer, A.; Coertse, J.; Makalo, M.J.; Molomo, M.; Markotter, W.; Nel, L.H. Epidemiology of Rabies in Lesotho: The Importance of Routine Surveillance and Virus Characterization. Trop. Med. Infect. Dis. 2017, 2, 30. https://doi.org/10.3390/tropicalmed2030030
Coetzer A, Coertse J, Makalo MJ, Molomo M, Markotter W, Nel LH. Epidemiology of Rabies in Lesotho: The Importance of Routine Surveillance and Virus Characterization. Tropical Medicine and Infectious Disease. 2017; 2(3):30. https://doi.org/10.3390/tropicalmed2030030
Chicago/Turabian StyleCoetzer, Andre, Jessica Coertse, Mabusetsa Joseph Makalo, Marosi Molomo, Wanda Markotter, and Louis Hendrik Nel. 2017. "Epidemiology of Rabies in Lesotho: The Importance of Routine Surveillance and Virus Characterization" Tropical Medicine and Infectious Disease 2, no. 3: 30. https://doi.org/10.3390/tropicalmed2030030
APA StyleCoetzer, A., Coertse, J., Makalo, M. J., Molomo, M., Markotter, W., & Nel, L. H. (2017). Epidemiology of Rabies in Lesotho: The Importance of Routine Surveillance and Virus Characterization. Tropical Medicine and Infectious Disease, 2(3), 30. https://doi.org/10.3390/tropicalmed2030030