Confronting the Emerging Threat to Public Health in Northern Australia of Neglected Indigenous Arboviruses



{kind=link}
Abstract
:1. Introduction
2. Arbovirus Ecology and Epidemiology
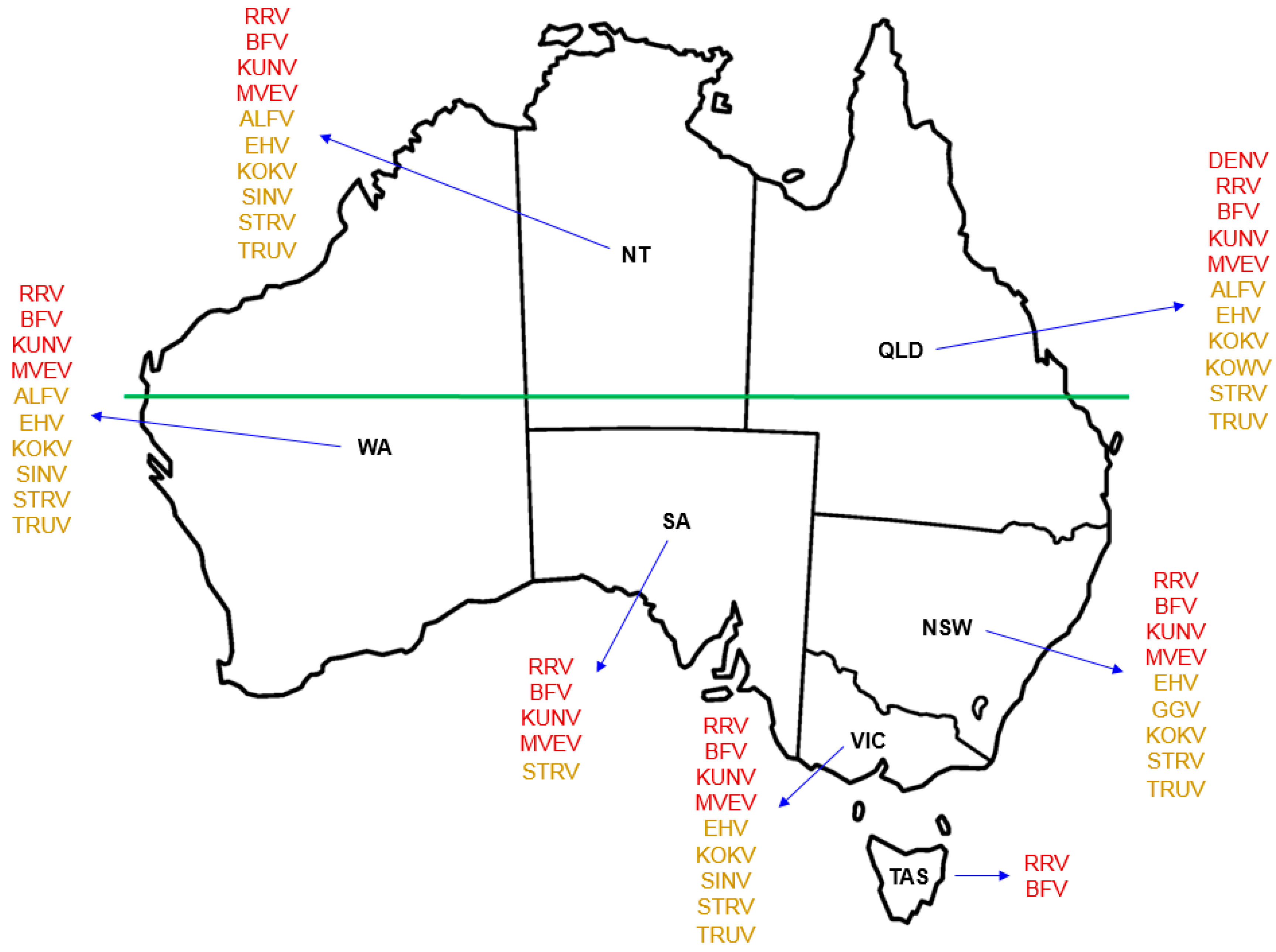
3. Arboviruses in Australia
4. Undifferentiated Febrile Illness and Pyrexia of Unknown Origin
5. UFI/PUO as a Health Problem
6. Diagnosis of Australian Arboviral Infection
7. A Causal Link between Neglected Arboviral Infections and UFI/PUO?
8. Transmission Cycles of Australian Arboviruses
9. Northern Australia’s Climate Favours Arboviruses
10. Potential Public Health Threat
11. A Call to Arms for Novel Diagnostic Tests and Therapy Targets
12. One Component of a ‘One Health’ Approach to Combating Arboviruses
13. Conclusions and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Arboviruses and Human Disease: Report of a WHO Scientific Group; Technical Report Series No. 369; WHO: Geneva, Switzerland, 1967. [Google Scholar]
- Centers for Disease Control and Prevention. Arbovirus Catalog. Available online: https://wwwn.cdc.gov/arbocat/ (accessed on 11 September 2017).
- Centers for Disease Control and Prevention. National Notifiable Diseases Surveillance System (NNDSS)—Arboviral Diseases, Neuroinvasive and Non-neuroinvasive 2015 Case Definition. Available online: https://wwwn.cdc.gov/nndss/conditions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive/case-definition/2015/ (accessed on 11 September 2017).
- Beckham, J.D.; Tyler, K.L. Arbovirus infections. Continuum (Minneapolis, Minn) 2015, 21, 1599–1611. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, N.; Bradbury, R.S.; Aaskov, J.G.; Taylor-Robinson, A.W. Neglected Australian arboviruses: Quam gravis? Microbes Infect. 2017, 19, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Australian Government Department of Health. National Notifiable Diseases Surveillance System. Notifications for All Diseases by State & Territory and Year. Available online: http://www9.health.gov.au/cda/source/rpt_2_sel.cfm (accessed on 11 September 2017).
- Weaver, S.C.; Barrett, A.D. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Young, P.R.; Ng, L.F.P.; Hall, R.A.; Smith, D.W.; Johansen, C.A. Arbovirus infection. In Manson’s Tropical Diseases, 23rd ed.; Farrar, J., Hotez, P.J., Junghanss, T., Kang, G., Lalloo, D., White, N.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 129–161. [Google Scholar]
- Morens, D.M.; Fauci, A.S. Emerging infectious diseases: Threats to human health and global stability. PLoS Pathog. 2013, 9, e1003467. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.-M.; Musso, D. Emerging arboviruses in the Pacific. Lancet 2014, 384, 1571–1572. [Google Scholar] [CrossRef]
- Gubler, D.J. Dengue viruses: Their evolution, history and emergence as a global public health problem. In Dengue and Dengue Hemorrhagic Fever; Gubler, D.J., Ooi, E., Vasudevan, S., Farrar, J., Eds.; CAB International: Wallingford, UK, 2014; pp. 1–29. [Google Scholar]
- Mackenzie, J.S.; Gubler, D.J.; Peterson, L.R. Emerging flaviviruses: The spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Murray, K.O. Dengue, West Nile virus, chikungunya, Zika and now Mayaro? PLoS Negl. Trop. Dis. 2017, 11, e0005462. [Google Scholar] [CrossRef] [PubMed]
- Sutherst, R.W. Global change and human vulnerability to vector-borne diseases. Clin. Microbiol. Rev. 2004, 17, 136–173. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C. Mosquito-borne disease and climate change in Australia: Time for a reality check. Aust. J. Entomol. 2009, 48, 1–7. [Google Scholar] [CrossRef]
- Fraser, J.R. Epidemic polyarthritis and Ross River virus disease. Clin. Rheum. Dis. 1986, 12, 369–388. [Google Scholar] [PubMed]
- Vale, T.G.; Carter, I.W.; McPhie, K.A.; James, G.; Cloonan, M.J. Human arbovirus infections along the south coast of New South Wales. Aust. J. Exp. Biol. Med. Sci. 1986, 64, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C. Arboviruses and their vectors in Australia: An update on the ecology and epidemiology of some mosquito-borne arboviruses. Rev. Med. Vet. Entomol. 1995, 83, 141–158. [Google Scholar]
- Gyawali, N.; Bradbury, R.S.; Taylor-Robinson, A.W. The epidemiology of dengue infection: Harnessing past experience and current knowledge to support implementation of future control strategies. J. Vector Borne Dis. 2016, 53, 293–304. [Google Scholar] [PubMed]
- Naish, S.; Tong, S. Hot spot detection and spatio-temporal dynamics of dengue in Queensland, Australia. In Proceedings of the ISPRS Technical Commission VIII Symposium, Hyderabad, India, 9–12 December 2014; Dadhwal, V.K., Diwakar, P.G., Seshasai, M.V.R., Raju, P.L.N., Hakeem, A., Eds.; International Society of Photogrammetry and Remote Sensing. pp. 197–204. [Google Scholar]
- Gyawali, N.; Bradbury, R.S.; Taylor-Robinson, A.W. Knowledge, attitude and recommendations for practice regarding dengue among the resident population of Queensland, Australia. Asian Pac. J. Trop. Biomed. 2016, 6, 360–366. [Google Scholar] [CrossRef]
- Hawkes, R.A.; Boughton, C.R.; Naim, H.M.; Wild, J.; Chapman, B. Arbovirus infections of humans in New South Wales. Seroepidemiology of the flavivirus group of togaviruses. Med. J. Aust. 1985, 143, 555–561. [Google Scholar] [PubMed]
- Mackenzie, J.; Lindsay, M.; Coelen, R.; Broom, A.; Hall, R.; Smith, D. Arboviruses causing human disease in the Australasian zoogeographic region. Arch. Virol. 1994, 136, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Guard, R.W.; McAuliffe, M.; Stallman, N.; Bramston, B. Haemorrhagic manifestations with Sindbis infection. Case report. Pathology 1982, 14, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Aaskov, J.G.; Phillips, D.A.; Wiemers, M.A. Possible clinical infection with Edge Hill virus. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 452–453. [Google Scholar] [CrossRef]
- Boughton, C.R.; Hawkes, R.A.; Naim, H.M. Illness caused by a Kokobera-like virus in south-eastern Australia. Med. J. Aust. 1986, 145, 90–92. [Google Scholar] [PubMed]
- Australian Government Department of Health. Australian National Notifiable Diseases by Disease Type. Available online: http://www.health.gov.au/internet/main/publishing.nsf/Content/cda-surveil-nndss-casedefs-distype.htm (accessed on 11 September 2017).
- Colmant, A.M.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Suen, W.W.; O’Brien, C.A.; van den Hurk, A.F.; Hall, R.A. A newly discovered flavivirus in the yellow fever virus group displays restricted replication in vertebrates. J. Gen. Virol. 2016, 97, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.A.; Williams, S.H.; Melville, L.F.; Nicholson, J.; Hall, R.A.; Bielefeldt-Ohmann, H.; Prow, N.A.; Chidlow, G.R.; Wong, S.; Sinha, R.; et al. Characterization of Fitzroy River virus and serologic evidence of human and animal infection. Emerg. Infect. Dis. 2017, 23, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Petersdorf, R.G.; Beeson, P.B. Fever of unexplained origin: Report on 100 cases. Medicine 1961, 40, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Durack, D.T.; Street, A.C. Fever of unknown origin—Reexamined and redefined. Curr. Clin. Top. Infect. Dis. 1991, 11, 35–51. [Google Scholar] [PubMed]
- Mourad, O.; Palda, V.; Detsky, A.S. A comprehensive evidence-based approach to fever of unknown origin. Arch. Intern. Med. 2003, 163, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A.; Swartz, M.N. Fever of undetermined origin. N. Engl. J. Med. 1973, 289, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.B.; Featherstone, H.J.; Petersdorf, R.G. Fever of undetermined origin: diagnosis and follow-up of 105 cases, 1970–1980. Medicine 1982, 61, 269–292. [Google Scholar] [CrossRef] [PubMed]
- De Kleijn, E.M.; Vandenbroucke, J.P.; van der Meer, J.W.; Group, N.F.S. Fever of unknown origin (FUO): I. A prospective multicenter study of 167 patients with FUO, using fixed epidemiologic entry criteria. Medicine 1997, 76, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Boivin, G.; Hardy, I.; Tellier, G.; Maziade, J. Predicting influenza infections during epidemics with use of a clinical case definition. Clin. Infect. Dis. 2000, 31, 1166–1169. [Google Scholar] [CrossRef] [PubMed]
- Rongrungruang, Y.; Leelarasamee, A. Characteristics and outcomes of adult patients with symptomatic dengue virus infections. J. Infect. Dis. Antimicrob. Agents 2001, 18, 19–23. [Google Scholar]
- Efstathiou, S.P.; Pefanis, A.V.; Tsiakou, A.G.; Skeva, I.I.; Tsioulos, D.I.; Achimastos, A.D.; Mountokalakis, T.D. Fever of unknown origin: Discrimination between infectious and non-infectious causes. Eur. J. Intern. Med. 2010, 21, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Vanderschueren, S.; Knockaert, D.; Adriaenssens, T.; Demey, W.; Durnez, A.; Blockmans, D.; Bobbaers, H. From prolonged febrile illness to fever of unknown origin: The challenge continues. Arch. Intern. Med. 2003, 163, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Buysschaert, I.; Vanderschueren, S.; Blockmans, D.; Mortelmans, L.; Knockaert, D. Contribution of 18fluoro-deoxyglucose positron emission tomography to the work-up of patients with fever of unknown origin. Eur. J. Intern. Med. 2004, 15, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Bleeker-Rovers, C.P.; Vos, F.J.; de Kleijn, E.M.; Mudde, A.H.; Dofferhoff, T.S.; Richter, C.; Smilde, T.J.; Krabbe, P.F.; Oyen, W.J.; van der Meer, J.W. A prospective multicenter study on fever of unknown origin: The yield of a structured diagnostic protocol. Medicine 2007, 86, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Robine, A.; Hot, A.; Maucort-Boulch, D.; Iwaz, J.; Broussolle, C.; Sève, P. Fever of unknown origin in the 2000s: Evaluation of 103 cases over eleven years. Presse Méd. 2014, 43, e233–e240. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, M.; Uehara, Y.; Yokokawa, H.; Hosoda, T.; Watanabe, Y.; Shiga, T.; Inui, A.; Otsuki, Y.; Fujibayashi, K.; Isonuma, H.; Naito, T. Analysis of 256 cases of classic fever of unknown origin. Intern. Med. 2013, 53, 2471–2475. [Google Scholar] [CrossRef]
- Doherty, R.L.; Whitehead, R.H.; Gorman, B.M.; O’Gower, A.K. The isolation of a third group A arbovirus in Australia, with preliminary observations on its relationship to epidemic polyarthritis. Aust. J. Sci. 1963, 26, 183–184. [Google Scholar]
- Oseni, R.A.; Donaldson, M.D.; Dalglish, D.A.; Aaskov, J.G. Detection by ELISA of IgM antibodies to Ross River virus in serum from patients with suspected epidemic polyarthritis. Bull. World Health Organ. 1983, 61, 703–708. [Google Scholar] [PubMed]
- Hargreaves, J.; Longbottom, H.; Myint, H.; Herceg, A.; Oliver, G.; Curran, M.; Evans, D. Annual report of the National Notifiable Diseases Surveillance System, 1994. Commun. Dis. Intell. 1995, 19, 542–574. [Google Scholar]
- Tong, S.; Bi, P.; Hayes, J.; Donald, K.; Mackenzie, J. Geographic variation of notified Ross River virus infections in Queensland, Australia, 1985–1996. Am. J. Trop. Med. Hyg. 2001, 65, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Marshall, I.D.; Woodroofe, G.M.; Hirsch, S. Viruses recovered from mosquitoes and wildlife serum collected in the Murray Valley of south-eastern Australia, February 1974, during an epidemic of encephalitis. Aust. J. Exp. Biol. Med. Sci. 1982, 60, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Jacups, S.P.; Whelan, P.I.; Currie, B.J. Ross River virus and Barmah Forest virus infections: A review of history, ecology, and predictive models, with implications for tropical northern Australia. Vector Borne Zoonotic Dis. 2008, 8, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Australian Government Department of Health. National Notifiable Diseases: Australia’s Notifiable Diseases Status. Annual Reports of the National Notifiable Diseases Surveillance System, 1994–2014. Available online: http://www.health.gov.au/internet/main/publishing.nsf/Content/cda-pubs-annlrpt-nndssar.htm (accessed on 11 September 2017).
- Doherty, R.L.; Carley, J.G.; Filippich, C.; White, J.; Gust, I.D. Murray Valley encephalitis in Australia, 1974: Antibody response in cases and community. Aust. N.Z. J. Med. 1976, 6, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.A.; Aaskov, J.G.; Atkin, C.; Wiemers, M.A. Isolation of Kunjin virus from a patient with a naturally acquired infection. Med. J. Aust. 1992, 157, 190–191. [Google Scholar] [PubMed]
- Mackenzie, J.; Smith, D.; Broom, A.; Bucens, M. Australian encephalitis in Western Australia, 1978–1991. Med. J. Aust. 1993, 158, 591–595. [Google Scholar] [PubMed]
- Broom, A.; Whelan, P.; Smith, D.; Lindsay, M.; Melville, L.; Bolisetty, S.; Wheaton, G.; Brown, A.; Higgins, G. An outbreak of Australian encephalitis in Western Australia and Central Australia (Northern Territory and South Australia) during the 2000 wet season. Arbovirus Res. Aust. 2001, 8, 37–42. [Google Scholar]
- Doherty, R.L. Arthropod-borne viruses in Australia and their relation to infection and disease. Prog. Med. Virol. 1974, 17, 136–192. [Google Scholar] [PubMed]
- Susilawati, T.N.; McBride, W.J.H. Undiagnosed undifferentiated fever in far north Queensland, Australia: A retrospective study. Int. J. Infect. Dis. 2014, 27, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Frost, M.J.; Zhang, J.; Edmonds, J.H.; Prow, N.A.; Gu, X.; Davis, R.; Hornitzky, C.; Arzey, K.E.; Finlaison, D.; Hick, P.; et al. Characterization of virulent West Nile virus Kunjin strain, Australia, 2011. Emerg. Infect. Dis. 2012, 18, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Doherty, R.L.; Gorman, B.M.; Whitehead, R.H.; Carley, J.G. Studies of arthropod-borne virus infections in Queensland. V. Survey of antibodies to group A arboviruses in man and other animals. Aust. J. Exp. Biol. Med. Sci. 1966, 44, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Inglis, T.J.; Bradbury, R.S.; McInnes, R.L.; Frances, S.P.; Merritt, A.J.; Levy, A.; Nicholson, J.; Neville, P.J.; Lindsay, M.; Smith, D.W. Deployable molecular detection of arboviruses in the Australian Outback. Am. J. Trop. Med. Hyg. 2016, 95, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Vale, T.G.; Spratt, D.M.; Cloonan, M.J. Serological evidence of arbovirus infection in native and domesticated mammals on the south coast of New South Wales. Aust. J. Zool. 1991, 39, 1–7. [Google Scholar] [CrossRef]
- Anderson, S.G. Murray Valley encephalitis and Australian X disease. Epidemiol. Infect. 1954, 52, 447–468. [Google Scholar] [CrossRef]
- Doherty, R. Arboviruses of Australia. Aust. Vet. J. 1972, 48, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Australian Government. Northern Australia Infrastructure Facility Act 2016. Available online: https://www.legislation.gov.au/Details/C2016A00041 (accessed on 11 September 2017).
- Australian Government. Australian Weather and Seasons—A Variety of Climates. Available online: http://www.australia.gov.au/about-australia/australian-story/austn-weather-and-the-seasons (accessed on 11 September 2017).
- Parham, P.E.; Waldock, J.; Christophides, G.K.; Hemming, D.; Agusto, F.; Evans, K.J.; Fefferman, N.; Gaff, H.; Gumel, A.; LaDeau, S.; et al. Climate, environmental and socio-economic change: Weighing up the balance in vector-borne disease transmission. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20130551. [Google Scholar] [CrossRef] [PubMed]
- Van den Hurk, A.F.; Craig, S.B.; Tulsiani, S.M.; Jansen, C.C. Emerging tropical diseases in Australia. Part 4. Mosquito-borne diseases. Ann. Trop. Med. Parasitol. 2010, 104, 623–640. [Google Scholar] [CrossRef] [PubMed]
- Australian Government. Our North, Our Future: White Paper on Developing Northern Australia. Available online: http://northernaustralia.gov.au/sites/prod.office-northern-australia.gov.au/files/files/NAWP-FullReport.pdf (accessed on 11 September 2017).
- Cummings Economics, March 2015. Long-Term Population Growth in Northern Australia. Available online: http://www.cummings.net.au/pdf/recent/J2806NthnAusLongTermPopulationGrowth.pdf (accessed on 11 September 2017).
- Inglis, T.J. Climate change and infectious diseases in Australia. Aust. Prescr. 2009, 32, 58–59. [Google Scholar] [CrossRef]
- Selvey, L.A.; Johansen, C.A.; Broom, A.K.; Antão, C.; Lindsay, M.D.; Mackenzie, J.S.; Smith, D.W. Rainfall and sentinel chicken seroconversions predict human cases of Murray Valley encephalitis in the north of Western Australia. BMC Infect. Dis. 2014, 14, 672. [Google Scholar] [CrossRef] [PubMed]
- Tall, J.A.; Gatton, M.L.; Tong, S. Ross River virus disease activity associated with naturally occurring nontidal flood events in Australia: A systematic review. J. Med. Entomol. 2014, 51, 1097–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, T.R.; McBride, J.L.; Chan, J.; Emanuel, K.; Holland, G.; Landsea, C.; Held, I.; Kossin, J.P.; Srivastava, A.K.; Sugi, M. Tropical cyclones and climate change. Nat. Geosci. 2010, 3, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Mayer, S.V.; Tesh, R.B.; Vasilakis, N. The emergence of arthropod-borne viral diseases: A global prospective on dengue, chikungunya and Zika fevers. Acta Trop. 2017, 166, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Quan, P.L.; Jabado, O.J.; Conlan, S.; Hirschberg, D.L.; Liu, Y.; Zhai, J.; Renwick, N.; Hui, J.; Hegyi, H.; et al. Panmicrobial oligonucleotide array for diagnosis of infectious diseases. Emerg. Infect. Dis. 2007, 13, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Giry, C.; Roquebert, B.; Li-Pat-Yuen, G.; Gasque, P.; Jaffar-Bandjee, M.C. Improved detection of genus-specific Alphavirus using a generic TaqMan® assay. BMC Microbiol. 2017, 17, 164. [Google Scholar] [CrossRef] [PubMed]
- Vina-Rodriguez, A.; Sachse, K.; Ziegler, U.; Chaintoutis, S.C.; Keller, M.; Groschup, M.H.; Eiden, M. A novel pan-Flavivirus detection and identification assay based on RT-qPCR and microarray. Biomed. Res. Int. 2017, 4248756. [Google Scholar] [CrossRef] [PubMed]
- Rosani, U.; Gerdol, M. A bioinformatics approach reveals seven nearly-complete RNA-virus genomes in bivalve RNA-seq data. Virus Res. 2017, 239, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Khou, C.; Pardigon, N. Identifying attenuating mutations: Tools for a new vaccine design against flaviviruses. Intervirology 2017, 60, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Mishra, S.; Milhotra, A.; Nagpal, R.; Mohan, M.; Singhal, A.; Kumari, P. Challenges with mosquito-borne viral diseases: Outbreak of the monsters. Curr. Top. Med. Chem. 2017, 17, 2199–2214. [Google Scholar] [CrossRef] [PubMed]
- Sun Star Manila, 25 April 2016. DOH Records 362 Adverse Reactions of Dengue Vaccine. Available online: http://www.sunstar.com.ph/manila/local-news/2016/04/25/doh-records-362-adverse-effects-dengue-vaccine-469921 (accessed on 11 September 2017).
- Malay Mail Online, 7 April 2017. Controversial Dengue Vaccine Approved for Further Trials in Malaysia. Available online: http://www.themalaymailonline.com/malaysia/article/controversial-dengue-vaccine-approved-for-further-trials-in-malaysia#sErjCc1APGerWKmy.99 (accessed on 11 September 2017).
- Pierson, T.C.; Fremont, D.H.; Kuhn, R.J.; Diamond, M.S. Structural insights into the mechanisms of antibody-mediated neutralization of flavivirus infection: Implications for vaccine development. Cell Host Microbe 2008, 4, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Subedi, D.; Taylor-Robinson, A.W. Anti-idiotype antibody against pre-membrane-specific antibody as an adjunct to current dengue vaccination strategy. Immun. Dis. 2015, 3, 1–7. [Google Scholar]
- Fauci, A.S.; Morens, D.M. Zika virus in the Americas—Yet another arbovirus threat. N. Engl. J. Med. 2016, 374, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Stringer, E.M.; de Silva, A.M. The emerging Zika virus epidemic in the Americas: Research priorities. JAMA 2016, 315, 1945–1946. [Google Scholar] [CrossRef] [PubMed]
- Rüegg, S.R.; McMahon, B.J.; Häsler, B.; Esposito, R.; Nielsen, L.R.; Ifejika Speranza, C.; Ehlinger, T.; Peyre, M.; Aragrande, M.; Zinsstag, J.; et al. A blueprint to evaluate One Health. Front. Public Health 2017, 5, 20. [Google Scholar]
- Gyawali, N.; Bradbury, R.S.; Taylor-Robinson, A.W. Do neglected Australian arboviruses pose a global epidemic threat? Aust. N. Z. J. Public Health 2016, 40, 596. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.C.; Barrington, D.J. A transdisciplinary approach to managing emerging and resurging mosquito-borne diseases in the Western Pacific Region. Trop. Med. Infect. Dis. 2017, 2, 1. [Google Scholar] [CrossRef]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gyawali, N.; Taylor-Robinson, A.W. Confronting the Emerging Threat to Public Health in Northern Australia of Neglected Indigenous Arboviruses. Trop. Med. Infect. Dis. 2017, 2, 55. https://doi.org/10.3390/tropicalmed2040055
Gyawali N, Taylor-Robinson AW. Confronting the Emerging Threat to Public Health in Northern Australia of Neglected Indigenous Arboviruses. Tropical Medicine and Infectious Disease. 2017; 2(4):55. https://doi.org/10.3390/tropicalmed2040055
Chicago/Turabian StyleGyawali, Narayan, and Andrew W. Taylor-Robinson. 2017. "Confronting the Emerging Threat to Public Health in Northern Australia of Neglected Indigenous Arboviruses" Tropical Medicine and Infectious Disease 2, no. 4: 55. https://doi.org/10.3390/tropicalmed2040055
APA StyleGyawali, N., & Taylor-Robinson, A. W. (2017). Confronting the Emerging Threat to Public Health in Northern Australia of Neglected Indigenous Arboviruses. Tropical Medicine and Infectious Disease, 2(4), 55. https://doi.org/10.3390/tropicalmed2040055