Outer Membrane Protein A Conservation among Orientia tsutsugamushi Isolates Suggests Its Potential as a Protective Antigen and Diagnostic Target



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. O. tsutsugamushi DNA Samples Examined in This Study
2.2. PCR, Cloning, and DNA Sequence Analyses
2.3. In Silico Analyses and GenBank Accession Numbers of O. tsutsugamushi Strain ompA Sequences
2.4. Confirmation of ompA PCR Primer Specificity
2.5. Mice
2.6. Quantitative Real-Time PCR (qPCR)
3. Results
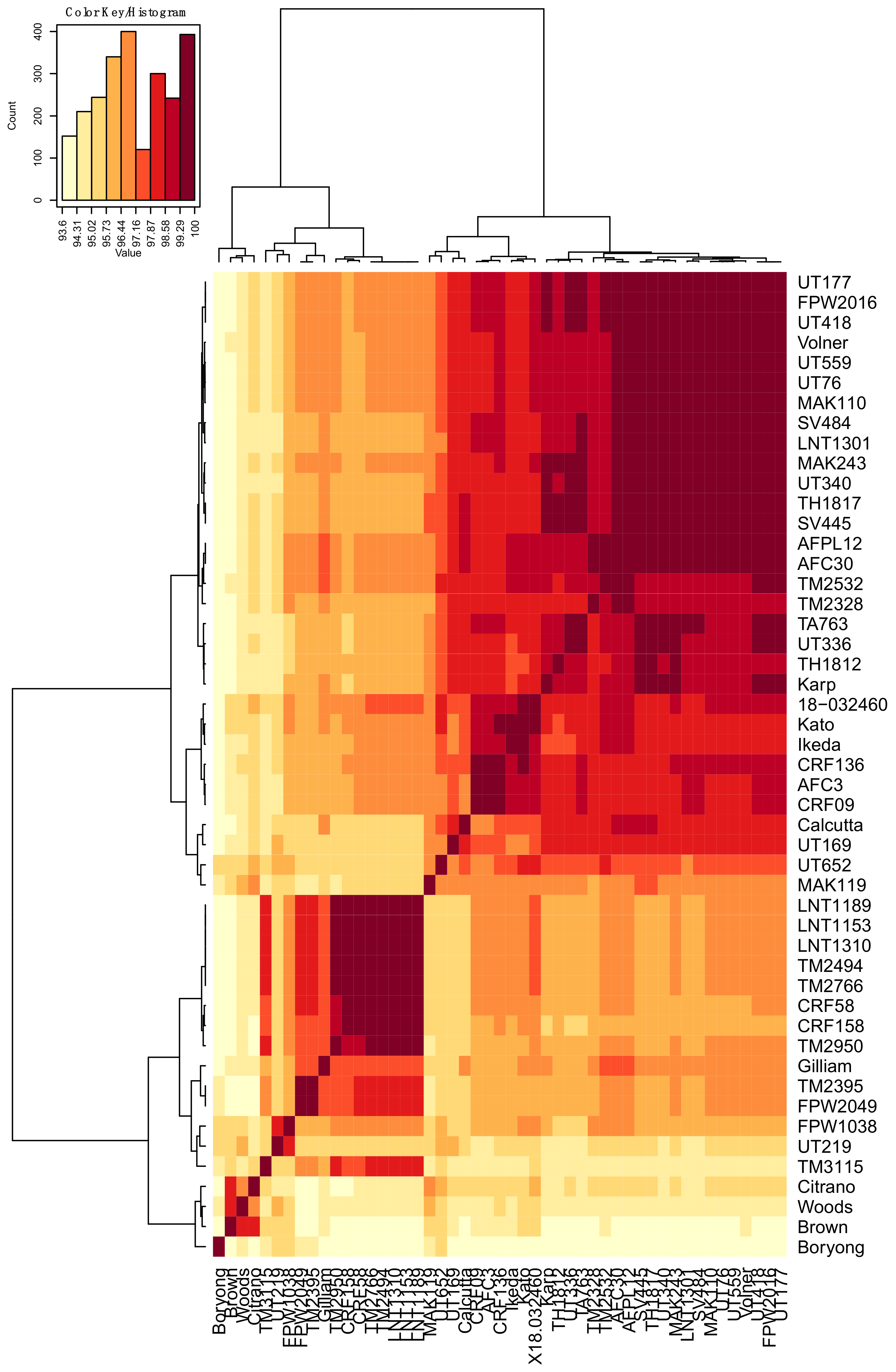
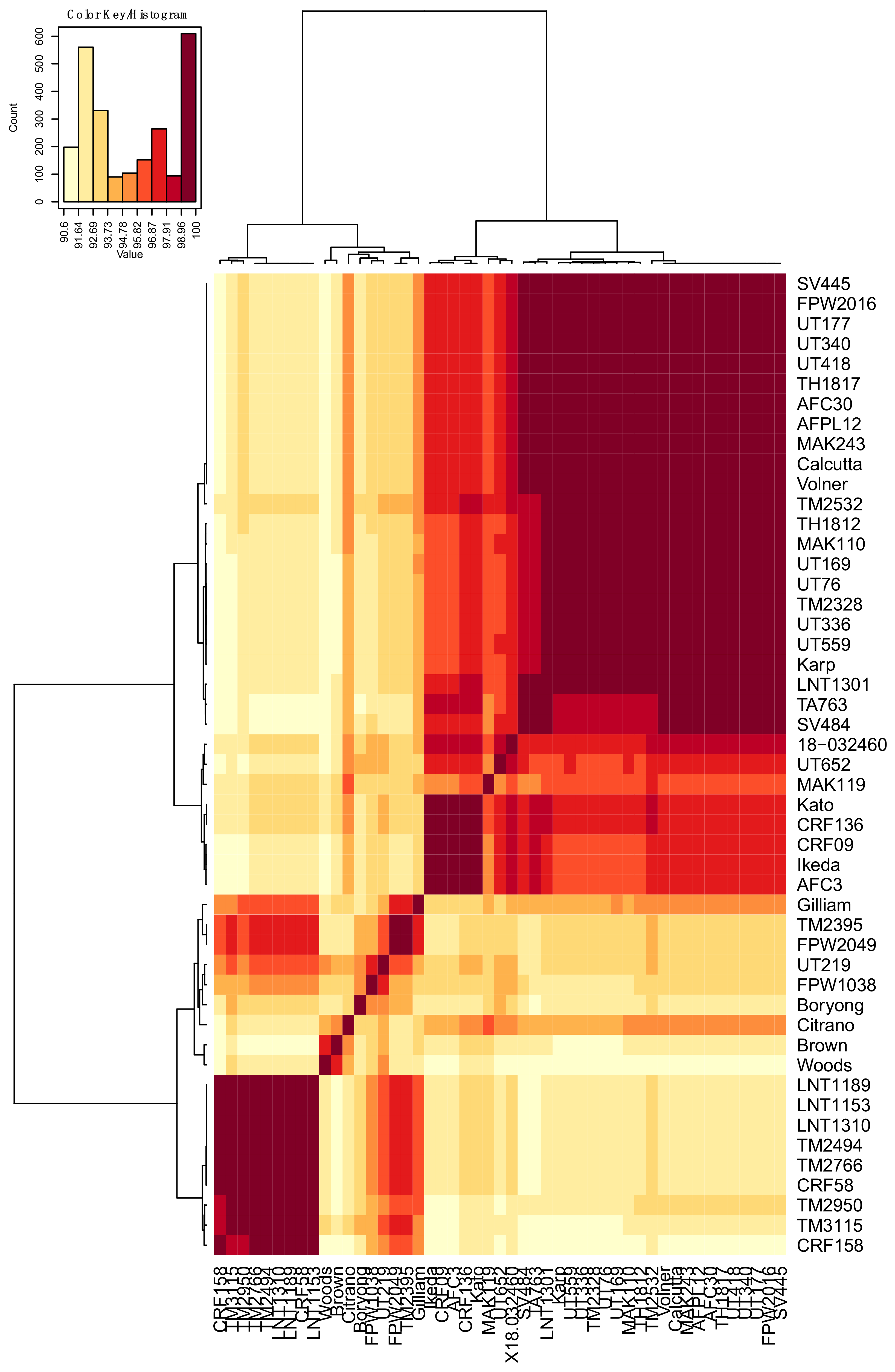
3.1. Analyses of O. tsutsugamushi ompA DNA and Translated Amino Acid Sequences
3.2. Molecular Modeling of OmpA
3.3. Development of a Primer Set That Specifically Amplifies an ompA Sequence Unique to O. tsutsugamushi
3.4. Evaluation of the O. tsutsugamushi ompA-Specific Primer Set in qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bonell, A.; Lubell, Y.; Newton, P.N.; Crump, J.A.; Paris, D.H. Estimating the burden of scrub typhus: A systematic review. PLoS Negl. Trop. Dis. 2017, 11, e0005838. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Walker, D.H.; Jupiter, D.; Melby, P.C.; Arcari, C.M. A review of the global epidemiology of scrub typhus. PLoS Negl. Trop. Dis. 2017, 11, e0006062. [Google Scholar] [CrossRef] [PubMed]
- Kasper, M.R.; Blair, P.J.; Touch, S.; Sokhal, B.; Yasuda, C.Y.; Williams, M.; Richards, A.L.; Burgess, T.H.; Wierzba, T.F.; Putnam, S.D. Infectious etiologies of acute febrile illness among patients seeking health care in south-central Cambodia. Am. J. Trop. Med. Hyg. 2012, 86, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Suttinont, C.; Losuwanaluk, K.; Niwatayakul, K.; Hoontrakul, S.; Intaranongpai, W.; Silpasakorn, S.; Suwancharoen, D.; Panlar, P.; Saisongkorh, W.; Rolain, J.M.; et al. Causes of acute, undifferentiated, febrile illness in rural Thailand: Results of a prospective observational study. Ann. Trop. Med. Parasitol. 2006, 100, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Phongmany, S.; Rolain, J.M.; Phetsouvanh, R.; Blacksell, S.D.; Soukkhaseum, V.; Rasachack, B.; Phiasakha, K.; Soukkhaseum, S.; Frichithavong, K.; Chu, V.; et al. Rickettsial infections and fever, Vientiane, Laos. Emerg. Infect. Dis. 2006, 12, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, G.; Walker, D.H. Approaches to vaccines against Orientia tsutsugamushi. Front. Cell. Infect. Microbiol. 2012, 2, 170. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Richards, A.L.; Temenak, J.; Strickman, D.; Dasch, G.A. The past and present threat of rickettsial diseases to military medicine and international public health. Clin. Infect. Dis. 2002, 34, S145–S169. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Paris, D.H.; Newton, P.N. A systematic review of mortality from untreated scrub typhus (Orientia tsutsugamushi). PLoS Negl. Trop. Dis. 2015, 9, e0003971. [Google Scholar] [CrossRef] [PubMed]
- Varghese, G.M.; Abraham, O.C.; Mathai, D.; Thomas, K.; Aaron, R.; Kavitha, M.L.; Mathai, E. Scrub typhus among hospitalised patients with febrile illness in South India: Magnitude and clinical predictors. J. Infect. 2006, 52, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Rodkvamtook, W.; Kuttasingkee, N.; Linsuwanon, P.; Sudsawat, Y.; Richards, A.L.; Somsri, M.; Sangjun, N.; Chao, C.C.; Davidson, S.; Wanja, E.; et al. Scrub typhus outbreak in Chonburi Province, Central Thailand, 2013. Emerg. Infect. Dis. 2018, 24, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Sedhain, A.; Bhattarai, G.R. Renal manifestation in scrub typhus during a major outbreak in central Nepal. Indian J. Nephrol. 2017, 27, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Vivian Thangaraj, J.W.; Mittal, M.; Verghese, V.P.; Kumar, C.P.G.; Rose, W.; Sabarinathan, R.; Pandey, A.K.; Gupta, N.; Murhekar, M. Scrub typhus as an etiology of acute febrile illness in Gorakhpur, Uttar Pradesh, India, 2016. Am. J. Trop. Med. Hyg. 2017, 97, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- Takhar, R.P.; Bunkar, M.L.; Arya, S.; Mirdha, N.; Mohd, A. Scrub typhus: A prospective, observational study during an outbreak in Rajasthan, India. Natl. Med. J. India 2017, 30, 69–72. [Google Scholar] [PubMed]
- Tshokey, T.; Graves, S.; Tshering, D.; Phuntsho, K.; Tshering, K.; Stenos, J. Scrub typhus outbreak in a remote primary school, Bhutan, 2014. Emerg. Infect. Dis. 2017, 23, 1412–1414. [Google Scholar] [CrossRef] [PubMed]
- Usha, K.; Kumar, E.; Kalawat, U.; Kumar, B.S.; Chaudhury, A.; Gopal, D.V. Molecular characterization of Orientia tsutsugamushi serotypes causing scrub typhus outbreak in southern region of Andhra Pradesh, India. Indian J. Med. Res. 2016, 144, 597–603. [Google Scholar] [PubMed]
- Marks, M.; Joshua, C.; Longbottom, J.; Longbottom, K.; Sio, A.; Puiahi, E.; Jilini, G.; Stenos, J.; Dalipanda, T.; Musto, J. An outbreak investigation of scrub typhus in Western Province, Solomon Islands, 2014. West. Pac. Surveill. Response J. 2016, 7, 6–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakharia, A.; Borkakoty, B.; Biswas, D.; Yadav, K.; Mahanta, J. Seroprevalence of scrub typhus infection in Arunachal Pradesh, India. Vector Borne Zoonotic Dis. 2016, 16, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.N.A.; Oltvolgyi, C.; Islam, A.; Hussain-Yusuf, H.; Loewenthal, M.R.; Vincent, G.; Stenos, J.; Graves, S. An outbreak of scrub typhus in military personnel despite protocols for antibiotic prophylaxis: Doxycycline resistance excluded by a quantitative PCR-based susceptibility assay. Microbes Infect. 2016, 18, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H. Recent outbreak of scrub typhus in North Western part of India. Indian J. Med. Microbiol. 2016, 34, 114. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Krishna, V.P.; Manjunath; Singh, H.; Shrivastava, S.; Singh, V.; Dariya, S.S.; Soni, M.; Sharma, S. Analysis of two outbreaks of scrub typhus in Rajasthan: A clinico-epidemiological study. J. Assoc. Physicians India 2014, 62, 24–29. [Google Scholar] [PubMed]
- Hu, J.; Tan, Z.; Ren, D.; Zhang, X.; He, Y.; Bao, C.; Liu, D.; Yi, Q.; Qian, W.; Yin, J.; et al. Clinical characteristics and risk factors of an outbreak with scrub typhus in previously unrecognized areas, Jiangsu province, China 2013. PLoS ONE 2015, 10, e0125999. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Bi, Z.; Kou, Z.; Yang, H.; Zhang, A.; Zhang, S.; Meng, X.; Zheng, L.; Zhang, M.; Yang, H.; et al. Scrub typhus caused by Orientia tsutsugamushi Kawasaki-related genotypes in Shandong Province, northern China. Infect. Genet. Evol. 2015, 30, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Luo, L.; Jing, Q.; Li, X.; Huang, Y.; Xiao, X.; Liu, L.; Wu, X.; Yang, Z. A city park as a potential epidemic site of scrub typhus: A case-control study of an outbreak in Guangzhou, China. Parasites Vectors 2014, 7, 513. [Google Scholar] [PubMed]
- Weitzel, T.; Dittrich, S.; Lopez, J.; Phuklia, W.; Martinez-Valdebenito, C.; Velasquez, K.; Blacksell, S.D.; Paris, D.H.; Abarca, K. Endemic scrub typhus in South America. N. Engl. J. Med. 2016, 375, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Horton, K.C.; Jiang, J.; Maina, A.; Dueger, E.; Zayed, A.; Ahmed, A.A.; Pimentel, G.; Richards, A.L. Evidence of Rickettsia and Orientia infections among abattoir workers in Djibouti. Am. J. Trop. Med. Hyg. 2016, 95, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Thiga, J.W.; Mutai, B.K.; Eyako, W.K. High seroprevalence of antibodies against spotted fever and scrub typhus bacteria in patients with febrile Illness, Kenya. Emerg. Infect. Dis. 2015, 21, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Balcells, M.E.; Rabagliati, R.; Garcia, P.; Poggi, H.; Oddo, D.; Concha, M.; Abarca, K.; Jiang, J.; Kelly, D.J.; Richards, A.L.; et al. Endemic scrub typhus-like illness, Chile. Emerg. Infect. Dis. 2011, 17, 1659–1663. [Google Scholar] [CrossRef] [PubMed]
- Groen, J.; Nur, Y.A.; Dolmans, W.; Ligthelm, R.J.; Osterhaus, A.D. Scrub and murine typhus among Dutch travellers. Infection 1999, 27, 291–292. [Google Scholar] [PubMed]
- Osuga, K.; Kimura, M.; Goto, H.; Shimada, K.; Suto, T. A case of tsutsugamushi disease probably contracted in Africa. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Kocher, C.; Jiang, J.; Morrison, A.C.; Castillo, R.; Leguia, M.; Loyola, S.; Ampuero, J.S.; Cespedes, M.; Halsey, E.S.; Bausch, D.G.; et al. Serologic evidence of scrub typhus in the Peruvian Amazon. Emerg. Infect. Dis. 2017, 23, 1389–1391. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Fuerst, P.A.; Ching, W.M.; Richards, A.L. Scrub typhus: The geographic distribution of phenotypic and genotypic variants of Orientia tsutsugamushi. Clin. Infect. Dis. 2009, 48 (Suppl. 3), S203–S230. [Google Scholar] [CrossRef] [PubMed]
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P.; et al. Isolation of a novel Orientia species (O. chuto sp. nov.) from a patient infected in Dubai. J. Clin. Microbiol. 2010, 48, 4404–4409. [Google Scholar] [CrossRef] [PubMed]
- Janardhanan, J.; Trowbridge, P.; Varghese, G.M. Diagnosis of scrub typhus. Expert Rev. Anti-Infect. Ther. 2014, 12, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Paris, D.H.; Aukkanit, N.; Jenjaroen, K.; Blacksell, S.D.; Day, N.P. A highly sensitive quantitative real-time PCR assay based on the groEL gene of contemporary Thai strains of Orientia tsutsugamushi. Clin. Microbiol. Infect. 2009, 15, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M.; Kim, H.L.; Park, C.Y.; Yang, T.Y.; Lee, J.H.; Yang, J.T.; Shim, S.K.; Lee, S.H. Clinical usefulness of eschar polymerase chain reaction for the diagnosis of scrub typhus: A prospective study. Clin. Infect. Dis. 2006, 43, 1296–1300. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chan, T.C.; Temenak, J.J.; Dasch, G.A.; Ching, W.M.; Richards, A.L. Development of a quantitative real-time polymerase chain reaction assay specific for Orientia tsutsugamushi. Am. J. Trop. Med. Hyg. 2004, 70, 351–356. [Google Scholar] [PubMed]
- Furuya, Y.; Yoshida, Y.; Katayama, T.; Yamamoto, S.; Kawamura, A., Jr. Serotype-specific amplification of Rickettsia tsutsugamushi DNA by nested polymerase chain reaction. J. Clin. Microbiol. 1993, 31, 1637–1640. [Google Scholar] [PubMed]
- Cheng, Z.; Miura, K.; Popov, V.L.; Kumagai, Y.; Rikihisa, Y. Insights into the CtrA regulon in development of stress resistance in obligatory intracellular pathogen Ehrlichia chaffeensis. Mol. Microbiol. 2011, 82, 1217–1234. [Google Scholar] [CrossRef] [PubMed]
- Confer, A.W.; Ayalew, S. The OmpA family of proteins: Roles in bacterial pathogenesis and immunity. Vet. Microbiol. 2013, 163, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Godlewska, R.; Wisniewska, K.; Pietras, Z.; Jagusztyn-Krynicka, E.K. Peptidoglycan-associated lipoprotein (Pal) of Gram-negative bacteria: Function, structure, role in pathogenesis and potential application in immunoprophylaxis. FEMS Microbiol. Lett. 2009, 298, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hebert, K.S.; Seidman, D.; Oki, A.T.; Izac, J.; Emani, S.; Oliver, L.D., Jr.; Miller, D.P.; Tegels, B.K.; Kannagi, R.; Marconi, R.T.; et al. Anaplasma marginale outer membrane protein A is an adhesin that recognizes sialylated and fucosylated glycans and functionally depends on an essential binding domain. Infect. Immun. 2017, 85, e00968-16. [Google Scholar] [CrossRef] [PubMed]
- Hillman, R.D., Jr.; Baktash, Y.M.; Martinez, J.J. OmpA-mediated rickettsial adherence to and invasion of human endothelial cells is dependent upon interaction with α2β1 integrin. Cell. Microbiol. 2013, 15, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Prasadarao, N.V. Outer membrane protein A and OprF: Versatile roles in Gram-negative bacterial infections. FEBS J. 2012, 279, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Ojogun, N.; Kahlon, A.; Ragland, S.A.; Troese, M.J.; Mastronunzio, J.E.; Walker, N.J.; Viebrock, L.; Thomas, R.J.; Borjesson, D.L.; Fikrig, E.; et al. Anaplasma phagocytophilum outer membrane protein A interacts with sialylated glycoproteins to promote infection of mammalian host cells. Infect. Immun. 2012, 80, 3748–3760. [Google Scholar] [CrossRef] [PubMed]
- Seidman, D.; Hebert, K.S.; Truchan, H.K.; Miller, D.P.; Tegels, B.K.; Marconi, R.T.; Carlyon, J.A. Essential domains of Anaplasma phagocytophilum invasins utilized to infect mammalian host cells. PLoS Pathog. 2015, 11, e1004669. [Google Scholar] [CrossRef] [PubMed]
- Do, E.J.; Kim, J.E.; Park, J.M.; Lee, K.M.; Jung, M.Y.; Lee, H.J.; Cho, H.W.; Choi, Y.J.; Lee, S.H.; Park, K.H.; et al. Development of recombinant OmpA and OmpB proteins as diagnostic antigens for rickettsial disease. Microbiol. Immunol. 2009, 53, 368–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, P.E.; Roux, V.; Raoult, D. Phylogenetic analysis of spotted fever group rickettsiae by study of the outer surface protein rOmpA. Int. J. Syst. Bacteriol. 1998, 48 Pt 3, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gracner, M.; Avsic-Zupanc, T.; Punda-Polic, V.; Dolinsek, J.; Bouyer, D.; Walker, D.H.; Zavala-Castro, J.E.; Bradaric, N.; Crocquet-Valdes, P.A.; Duh, D. Comparative ompA gene sequence analysis of Rickettsia felis-like bacteria detected in Haemaphysalis sulcata ticks and isolated in the mosquito C6/36 cell line. Clin. Microbiol. Infect. 2009, 15 (Suppl. 2), 265–266. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wen, B.; Chen, M.; Niu, D.; Zhang, J.; Qiu, L. Analysis of immunoprotectivity of the recombinant OmpA of Rickettsia heilongjiangensis. Ann. N. Y. Acad. Sci. 2005, 1063, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Beyer, A.R.; Rodino, K.G.; VieBrock, L.; Green, R.S.; Tegels, B.K.; Oliver, L.D., Jr.; Marconi, R.T.; Carlyon, J.A. Orientia tsutsugamushi Ank9 is a multifunctional effector that utilizes a novel GRIP-like Golgi localization domain for Golgi-to-endoplasmic reticulum trafficking and interacts with host COPB2. Cell. Microbiol. 2017, 19, e12727. [Google Scholar] [CrossRef] [PubMed]
- Mayxay, M.; Castonguay-Vanier, J.; Chansamouth, V.; Dubot-Peres, A.; Paris, D.H.; Phetsouvanh, R.; Tangkhabuanbutra, J.; Douangdala, P.; Inthalath, S.; Souvannasing, P.; et al. Causes of non-malarial fever in Laos: A prospective study. Lancet Glob. Health 2013, 1, e46–e54. [Google Scholar] [CrossRef]
- Paris, D.H.; Blacksell, S.D.; Nawtaisong, P.; Jenjaroen, K.; Teeraratkul, A.; Chierakul, W.; Wuthiekanun, V.; Kantipong, P.; Day, N.P. Diagnostic accuracy of a loop-mediated isothermal PCR assay for detection of Orientia tsutsugamushi during acute scrub typhus infection. PLoS Negl. Trop. Dis. 2011, 5, e1307. [Google Scholar] [CrossRef] [PubMed]
- Blacksell, S.D.; Luksameetanasan, R.; Kalambaheti, T.; Aukkanit, N.; Paris, D.H.; McGready, R.; Nosten, F.; Peacock, S.J.; Day, N.P. Genetic typing of the 56-kDa type-specific antigen gene of contemporary Orientia tsutsugamushi isolates causing human scrub typhus at two sites in north-eastern and western Thailand. FEMS Immunol. Med. Microbiol. 2008, 52, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.H.; Kang, J.S.; Lee, W.K.; Choi, M.S.; Lee, J.H. Serological classification by monoclonal antibodies of Rickettsia tsutsugamushi isolated in Korea. J. Clin. Microbiol. 1990, 28, 685–688. [Google Scholar] [PubMed]
- Rights, F.L.; Smadel, H.I.; Jackson, E.B. Studies on scrub typhus (tsutsugamushi disease). III. Heterogeneity of strains of R. tsutsugamushi, as demonstrated by cross-vaccination studies. J. Exp. Med. 1948, 87, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Watt, G.; Kantipong, P.; Jongsakul, K.; Watcharapichat, P.; Phulsuksombati, D. Azithromycin activities against Orientia tsutsugamushi strains isolated in cases of scrub typhus in Northern Thailand. Antimicrob. Agents Chemother. 1999, 43, 2817–2818. [Google Scholar] [PubMed]
- Bengtson, I.A. Apparent serological heterogeneity among strains of tsutsugamushi disease (scrub typhus). Public Health Rep. 1945, 60, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Takahashi, K.; Tsuruhara, T.; Urakami, H.; Miyamura, S.; Sekikawa, H.; Kenmotsu, M.; Shibata, M.; Abe, S.; Nezu, H. Isolation of Rickettsia tsutsugamushi antigenically different from Kato, Karp, and Gilliam strains from patients. Microbiol. Immunol. 1984, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Elisberg, B.L.; Campbell, J.M.; Bozeman, F.M. Antigenic diversity of Rickettsia tsutsugamushi: Epidemiologic and ecologic significance. J. Hyg. Epidemiol. Microbiol. Immunol. 1968, 12, 18–25. [Google Scholar] [PubMed]
- Philip, C.B.; Woodward, T.E.; Sullivan, R.R. Tsutsugamushi disease (scrub or mite-borne typhus) in the Philippine Islands during American re-occupation in 1944–45. Am. J. Trop. Med. Hyg. 1946, 26, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Carley, J.G.; Doherty, R.L.; Derrick, E.H.; Pope, J.H.; Emanuel, M.L.; Ross, C.J. The investigation of fevers in North Queensland by mouse inoculation, with particular reference to scrub typhus. Australas. Ann. Med. 1955, 4, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Bourgeois, A.L. Rickettsia tsutsugamushi infection and scrub typhus incidence among Chinese military personnel in the Pescadores Islands. Am. J. Epidemiol. 1977, 106, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.; Sivarajah, A.; Kelly, D.J.; Lewis, G.E., Jr. An analysis of febrile illnesses among members of the Malaysian Police Field Force. Mil. Med. 1986, 151, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Shishido, A.; Ohtawara, M.; Tateno, S.; Mizuno, S.; Ogura, M.; Kitaoka, M. The nature of immunity against scrub typhus in mice. I. The resistance of mice, surviving subcutaneous infection of scrub typhus rickettsia, to intraperitoneal reinfection of the same agent. Jpn. J. Med. Sci. Biol. 1958, 11, 383–399. [Google Scholar] [CrossRef]
- Nakayama, K.; Yamashita, A.; Kurokawa, K.; Morimoto, T.; Ogawa, M.; Fukuhara, M.; Urakami, H.; Ohnishi, M.; Uchiyama, I.; Ogura, Y.; et al. The whole-genome sequencing of the obligate intracellular bacterium Orientia tsutsugamushi revealed massive gene amplification during reductive genome evolution. DNA Res. 2008, 15, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Sternberg, M.J. Partial protein domains: Evolutionary insights and bioinformatics challenges. Genome Biol. 2015, 16, 100. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.C.; Jiang, J.; Temenak, J.J.; Richards, A.L. Development of a rapid method for determining the infectious dose (ID)50 of Orientia tsutsugamushi in a scrub typhus mouse model for the evaluation of vaccine candidates. Vaccine 2003, 21, 4550–4554. [Google Scholar] [CrossRef]
- Sunyakumthorn, P.; Paris, D.H.; Chan, T.C.; Jones, M.; Luce-Fedrow, A.; Chattopadhyay, S.; Jiang, J.; Anantatat, T.; Turner, G.D.; Day, N.P.; et al. An intradermal inoculation model of scrub typhus in Swiss CD-1 mice demonstrates more rapid dissemination of virulent strains of Orientia tsutsugamushi. PLoS ONE 2013, 8, e54570. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noriea, N.F.; Clark, T.R.; Hackstadt, T. Targeted knockout of the Rickettsia rickettsii OmpA surface antigen does not diminish virulence in a mammalian model system. MBio 2015, 6, e00323-15. [Google Scholar] [CrossRef] [PubMed]
- Riley, S.P.; Goh, K.C.; Hermanas, T.M.; Cardwell, M.M.; Chan, Y.G.; Martinez, J.J. The Rickettsia conorii autotransporter protein Sca1 promotes adherence to nonphagocytic mammalian cells. Infect. Immun. 2010, 78, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Cardwell, M.M.; Martinez, J.J. The Sca2 autotransporter protein from Rickettsia conorii is sufficient to mediate adherence to and invasion of cultured mammalian cells. Infect. Immun. 2009, 77, 5272–5280. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.G.; Cardwell, M.M.; Hermanas, T.M.; Uchiyama, T.; Martinez, J.J. Rickettsial outer-membrane protein B (rOmpB) mediates bacterial invasion through Ku70 in an actin, c-Cbl, clathrin and caveolin 2-dependent manner. Cell. Microbiol. 2009, 11, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M.; Park, G.; Kim, H.S.; Lee, J.Y.; Neupane, G.P.; Graves, S.; Stenos, J. Comparison of conventional, nested, and real-time quantitative PCR for diagnosis of scrub typhus. J. Clin. Microbiol. 2011, 49, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Tantibhedhyangkul, W.; Wongsawat, E.; Silpasakorn, S.; Waywa, D.; Saenyasiri, N.; Suesuay, J.; Thipmontree, W.; Suputtamongkol, Y. Use of multiplex real-time PCR to diagnose scrub typhus. J. Clin. Microbiol. 2017, 55, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Le Viet, N.; Laroche, M.; Thi Pham, H.L.; Viet, N.L.; Mediannikov, O.; Raoult, D.; Parola, P. Use of eschar swabbing for the molecular diagnosis and genotyping of Orientia tsutsugamushi causing scrub typhus in Quang Nam province, Vietnam. PLoS Negl. Trop. Dis. 2017, 11, e0005397. [Google Scholar] [CrossRef] [PubMed]
- Premaratna, R.; Blanton, L.S.; Samaraweera, D.N.; de Silva, G.N.; Chandrasena, N.T.; Walker, D.H.; de Silva, H.J. Genotypic characterization of Orientia tsutsugamushi from patients in two geographical locations in Sri Lanka. BMC Infect. Dis. 2017, 17, 67. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Belinskaya, T.; Zhang, Z.; Ching, W.M. Development of recombinase polymerase amplification assays for detection of Orientia tsutsugamushi or Rickettsia typhi. PLoS Negl. Trop. Dis. 2015, 9, e0003884. [Google Scholar] [CrossRef] [PubMed]
- Watthanaworawit, W.; Turner, P.; Turner, C.; Tanganuchitcharnchai, A.; Richards, A.L.; Bourzac, K.M.; Blacksell, S.D.; Nosten, F. A prospective evaluation of real-time PCR assays for the detection of Orientia tsutsugamushi and Rickettsia spp. for early diagnosis of rickettsial infections during the acute phase of undifferentiated febrile illness. Am. J. Trop. Med. Hyg. 2013, 89, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Prakash, J.A.; Reller, M.E.; Barat, N.; Dumler, J.S. Assessment of a quantitative multiplex 5′ nuclease real-time PCR for spotted fever and typhus group rickettsioses and Orientia tsutsugamushi. Clin. Microbiol. Infect. 2009, 15 (Suppl. 2), 292–293. [Google Scholar] [CrossRef] [PubMed]
- Singhsilarak, T.; Leowattana, W.; Looareesuwan, S.; Wongchotigul, V.; Jiang, J.; Richards, A.L.; Watt, G. Short report: Detection of Orientia tsutsugamushi in clinical samples by quantitative real-time polymerase chain reaction. Am. J. Trop. Med. Hyg. 2005, 72, 640–641. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Geographic Origin | Reference | ompA GenBank Accession Number or Locus Tag |
|---|---|---|---|
| 18-032460 | Malaysia | [63] | MH167971 |
| AFC3 | Thailand | [56] | MH167972 |
| AFC30 | Thailand | NR | MH167973 |
| AFPL12 | Thailand | NR | MH167974 |
| Boryong | South Korea | [54] | OTBS_RS08365 |
| Brown | Australia | [61] | MH167975 |
| Calcutta | India | NR | MH167976 |
| Citrano | Australia | [61] | MH167977 |
| CRF09 | Northern Thailand | [52] | MH167978 |
| CRF58 | Northern Thailand | [52] | MH167979 |
| CRF136 | Northern Thailand | [52] | MH167980 |
| CRF158 | Northern Thailand | [52] | MH167981 |
| FPW1038 | Western Thailand | [53] | MH167982 |
| FPW2016 | Western Thailand | [53] | MH167983 |
| FPW2049 | Western Thailand | [53] | MH167984 |
| Gilliam | Burma | [57] | MH167985 |
| Ikeda | Japan | [58] | OTT_RS06375 |
| Karp | New Guinea | [55] | OTSKARP_0358 |
| Kato | Japan | [64] | OTSKATO_0610 |
| LNT1153 | Northwestern Laos | [51] | MH167986 |
| LNT1189 | Northwestern Laos | [51] | MH167987 |
| LNT1301 | Northwestern Laos | [51] | MH167988 |
| LNT1310 | Northwestern Laos | [51] | MH167989 |
| MAK110 | Taiwan | [62] | MH167990 |
| MAK119 | Taiwan | [62] | MH167991 |
| MAK243 | Taiwan | [62] | MH167992 |
| SV400 | Southern Laos | [51] | MH167993 |
| SV445 | Southern Laos | [51] | MH167994 |
| SV484 | Southern Laos | [51] | MH167995 |
| TA763 | Thailand | [59] | OTSTA763_0977 |
| TH1812 | Thailand | [59] | MH167996 |
| TH1817 | Thailand | [59] | MH167997 |
| TM2328 | Central Laos | [51] | MH167998 |
| TM2395 | Central Laos | [51] | MH167999 |
| TM2494 | Central Laos | [51] | MH168000 |
| TM2532 | Central Laos | [51] | MH168001 |
| TM2766 | Central Laos | [51] | MH168002 |
| TM2950 | Central Laos | [51] | MH168003 |
| TM3115 | Central Laos | [51] | MH168004 |
| UT76 | Northeastern Thailand | [53] | MH168005 |
| UT125 | Northeastern Thailand | [53] | MH168006 |
| UT169 | Northeastern Thailand | [53] | MH168007 |
| UT177 | Northeastern Thailand | [53] | MH168008 |
| UT219 | Northeastern Thailand | [53] | MH168009 |
| UT336 | Northeastern Thailand | [53] | MH168010 |
| UT340 | Northeastern Thailand | [53] | MH168011 |
| UT418 | Northeastern Thailand | [53] | MH168012 |
| UT559 | Northeastern Thailand | [53] | MH168013 |
| UT652 | Northeastern Thailand | [53] | MH168014 |
| Volner | New Guinea | [60] | MH168015 |
| Wood | Australia | [61] | MH168016 |
| Primer Designation a | Sequence (5′ to 3′) |
|---|---|
| OTT_RS06375-1F | ATGATTAAAAAGTCAATTATTAGTATATGTGTATTAGTGC |
| OTT_RS06375-615R | CTATGCTATATTACTTTTAATAATTGTGACAGACC |
| OTT_RS06375-64F | TGTTTATGGCAAAGATCTAAACATAGTAAC |
| ompA-57F | GTGGAAATGTTTATGGCAAAGATCTAAAC |
| ompA-260R | GCTTGTAAAAACTGTTCATGCTTTACATC |
| Eubacterial 16S-F | GTTCGGAATTACTGGGCGTA |
| Eubacterial 16S-R | AATTAAACCGCATGCTCCAC |
| R17K-135 | ATGAATAAACAACGKbCANGGHACAC |
| R17K-249 | RAAGTAATGCRCCTACACCTACTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, S.M.; Adcox, H.E.; VieBrock, L.; Green, R.S.; Luce-Fedrow, A.; Chattopadhyay, S.; Jiang, J.; Marconi, R.T.; Paris, D.; Richards, A.L.; et al. Outer Membrane Protein A Conservation among Orientia tsutsugamushi Isolates Suggests Its Potential as a Protective Antigen and Diagnostic Target. Trop. Med. Infect. Dis. 2018, 3, 63. https://doi.org/10.3390/tropicalmed3020063
Evans SM, Adcox HE, VieBrock L, Green RS, Luce-Fedrow A, Chattopadhyay S, Jiang J, Marconi RT, Paris D, Richards AL, et al. Outer Membrane Protein A Conservation among Orientia tsutsugamushi Isolates Suggests Its Potential as a Protective Antigen and Diagnostic Target. Tropical Medicine and Infectious Disease. 2018; 3(2):63. https://doi.org/10.3390/tropicalmed3020063
Chicago/Turabian StyleEvans, Sean M., Haley E. Adcox, Lauren VieBrock, Ryan S. Green, Alison Luce-Fedrow, Suschsmita Chattopadhyay, Ju Jiang, Richard T. Marconi, Daniel Paris, Allen L. Richards, and et al. 2018. "Outer Membrane Protein A Conservation among Orientia tsutsugamushi Isolates Suggests Its Potential as a Protective Antigen and Diagnostic Target" Tropical Medicine and Infectious Disease 3, no. 2: 63. https://doi.org/10.3390/tropicalmed3020063
APA StyleEvans, S. M., Adcox, H. E., VieBrock, L., Green, R. S., Luce-Fedrow, A., Chattopadhyay, S., Jiang, J., Marconi, R. T., Paris, D., Richards, A. L., & Carlyon, J. A. (2018). Outer Membrane Protein A Conservation among Orientia tsutsugamushi Isolates Suggests Its Potential as a Protective Antigen and Diagnostic Target. Tropical Medicine and Infectious Disease, 3(2), 63. https://doi.org/10.3390/tropicalmed3020063