Bats and Viruses: Emergence of Novel Lyssaviruses and Association of Bats with Viral Zoonoses in the EU

 , ,
, ,
Abstract
:1. Introduction
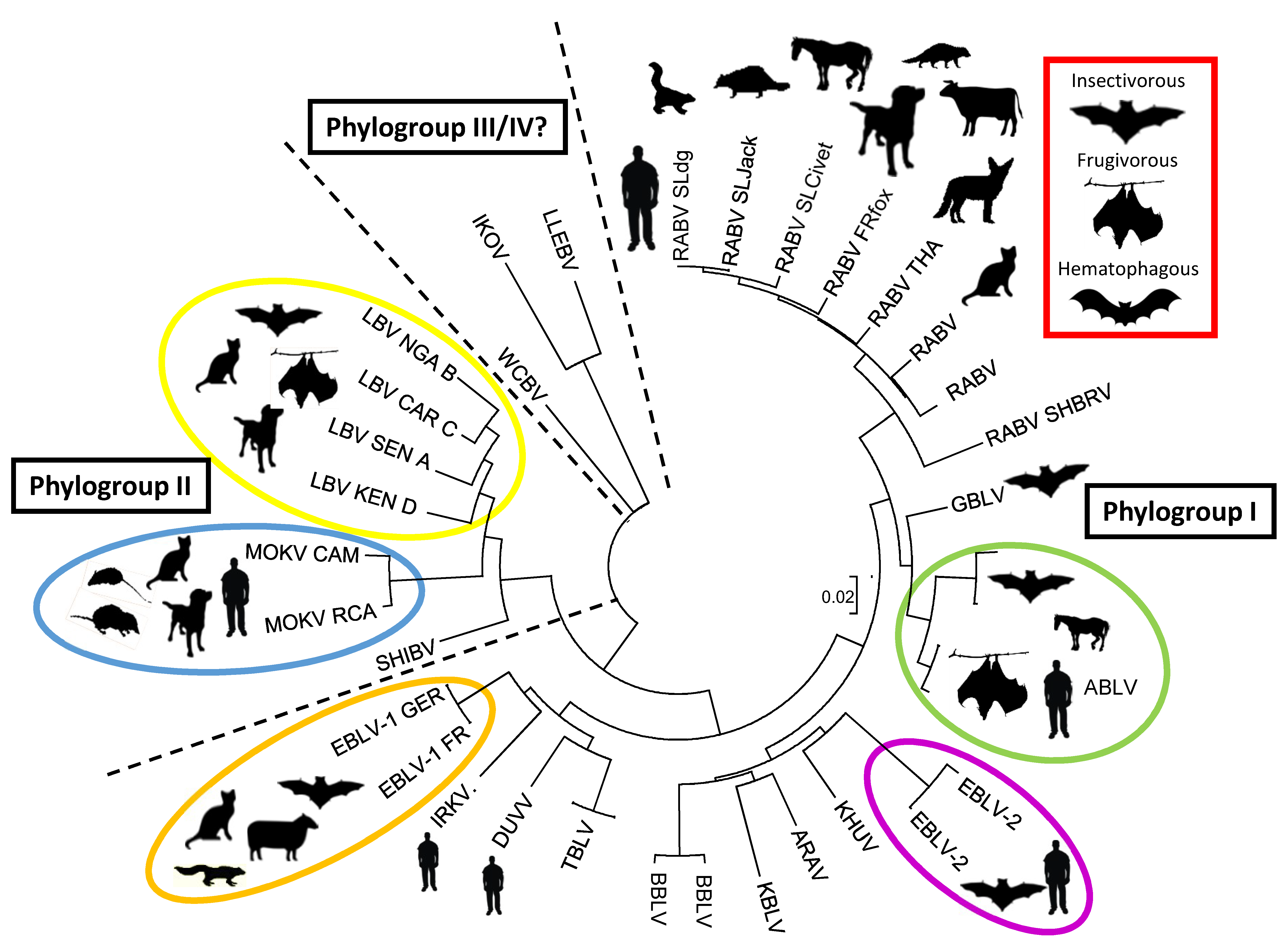
2. The Increasing Diversity of the Bat Lyssaviruses
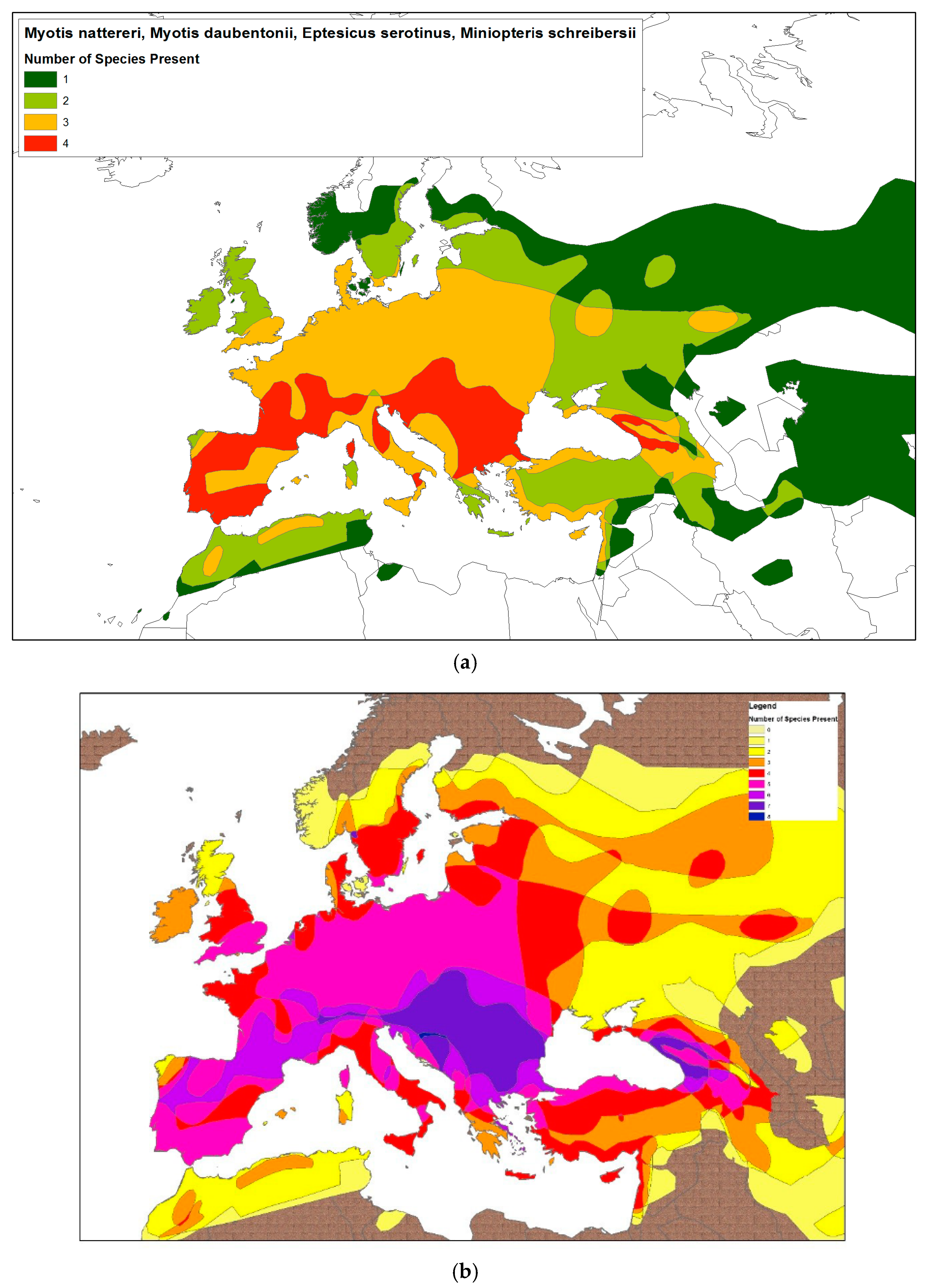
3. The Association of Other Viral Zoonoses with European Bat Species
4. Evolution of Viral Species within Bats
5. Bats and Their Role as a Reservoir for Viral Pathogens
6. Availability of Human Vaccines for Bat Pathogens
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fooks, A.R.; Banyard, A.C.; Horton, D.L.; Johnson, N.; McElhinney, L.M.; Jackson, A.C. Current status of rabies and prospects for elimination. Lancet 2014, 384, 1389–1399. [Google Scholar] [CrossRef]
- Banyard, A.C.; Fooks, A.R. The impact of novel lyssavirus discovery. Microbiol. Aust. 2017, 38, 18–21. [Google Scholar] [CrossRef]
- Dato, V.M.; Campagnolo, E.R.; Long, J.; Rupprecht, C.E. A Systematic Review of Human Bat Rabies Virus Variant Cases: Evaluating Unprotected Physical Contact with Claws and Teeth in Support of Accurate Risk Assessments. PLoS ONE 2016, 11, e0159443. [Google Scholar] [CrossRef] [PubMed]
- Banyard, A.C.; Horton, D.; Freuling, C.; Müller, T.; Fooks, A.R. Control and prevention of canine rabies: The need for building laboratory based surveillance capacity. Antivir. Res. 2013, 98, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Marston, D.A.; Horton, D.L.; Nunez, J.; Ellis, R.J.; Orton, R.J.; Johnson, N.; Banyard, A.C.; McElhinney, L.M.; Freuling, C.M.; Firat, M.; et al. Genetic analysis of a rabies virus host shift event reveals within-host viral dynamics in a new host. Virus Evol. 2017, 3, vex038. [Google Scholar] [CrossRef] [PubMed]
- Streicker, D.G.; Turmelle, A.S.; Vonhof, M.J.; Kuzmin, I.V.; McCracken, G.F.; Rupprecht, C.E. Host phylogeny constrains cross-species emergence and establishment of rabies virus in bats. Science 2010, 329, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M.J.; Messenger, S.; Rohde, R.E.; Smith, J.; Cheshier, R.; Hanlon, C.; Rupprecht, C.E. Bat-associated rabies virus in Skunks. Emerg. Infect. Dis. 2006, 12, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Daoust, P.Y.; Wandeler, A.I.; Casey, G.A. Cluster of rabies cases of probable bat origin among red foxes in Prince Edward Island, Canada. J. Wildl. Dis. 1996, 32, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.M.; Gilbert, A.; Slate, D.; Chipman, R.; Singh, A.; Cassie, W.; Blanton, J.D. Right place, wrong species: A 20-year review of rabies virus cross species transmission among terrestrial mammals in the United States. PLoS ONE 2014, 9, e107539. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Shi, M.; Orciari, L.A.; Yager, P.A.; Velasco-Villa, A.; Kuzmina, N.A.; Streicker, D.G.; Bergman, D.L.; Rupprecht, C.E. Molecular inferences suggest multiple host shifts of rabies viruses from bats to mesocarnivores in Arizona during 2001–2009. PLoS Pathog. 2012, 8, e1002786. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.; Kuzmin, I.; Meslin, F. Lyssaviruses and rabies: Current conundrums, concerns, contradictions and controversies. F1000Res 2017, 6, 184. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Muller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Primers 2017, 3, 17091. [Google Scholar] [CrossRef] [PubMed]
- Coertse, J.; Markotter, W.; le Roux, K.; Stewart, D.; Sabeta, C.T.; Nel, L.H. New isolations of the rabies-related Mokola virus from South Africa. BMC Vet. Res. 2017, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Markotter, W.; Kgaladi, J.; Nel, L.H.; Marston, D.; Wright, N.; Coertse, J.; Müller, T.F.; Sabeta, C.T.; Fooks, A.R.; Freuling, C.M. Diversity and Epidemiology of Mokola Virus. PLoS Negl. Trop. Dis. 2013, 7, e2511. [Google Scholar]
- Marston, D.A.; Horton, D.L.; Ngeleja, C.; Hampson, K.; McElhinney, L.M.; Banyard, A.C.; Haydon, D.; Cleaveland, S.; Rupprecht, C.E.; Bigambo, M.; et al. Ikoma lyssavirus, highly divergent novel lyssavirus in an African civet. Emerg. Infect. Dis. 2012, 18, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.C.; Hsu, C.L.; Lee, M.S.; Tu, Y.C.; Chang, J.C.; Wu, C.H.; Lee, S.H.; Ting, L.J.; Tsai, K.R.; Cheng, M.C.; et al. Lyssavirus in Japanese Pipistrelle, Taiwan. Emerg. Infect. Dis. 2018, 24, 782–785. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R. The challenge of new and emerging lyssaviruses. Expert Rev. Vaccines 2004, 3, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Banyard, A.C.; Evans, J.S.; Luo, T.R.; Fooks, A.R. Lyssaviruses and bats: Emergence and zoonotic threat. Viruses 2014, 6, 2974–2990. [Google Scholar] [CrossRef] [PubMed]
- Schindler, R.; Denning, H.K.D. Unter-suchungen über die Bedeutung der Fledermäusefür den gegenwärtigen Tollwut-Seuchenzug inDeutschland. Monatsschrift für Tierheilkunde 1958, 10, 169–177. [Google Scholar]
- Jelesic, Z.; Nikolic, M. Isolation of rabies virus from insectivorous bats in Yugoslavia. Bull. World Health Organ. 1956, 14, 801–804. [Google Scholar] [PubMed]
- King, A.A.; Haagsma, J.; Kappeler, A. Lyssavirus infections in European Bats. In Historical Perspective of Rabies in Europe and the Mediterranean Basin; King, A.A., Fooks, A.R., Aubert, M., Wandeler, A.I., Eds.; OIE: Paris, France, 2004; pp. 221–241. [Google Scholar]
- Marston, D.A.; McElhinney, L.M.; Ellis, R.J.; Horton, D.L.; Wise, E.L.; Leech, S.L.; David, D.; de Lambellerie, X.; Fooks, A.R. Next Generation Sequencing of viral RNA genomes. BMC Genom. 2013, 14, 444. [Google Scholar] [CrossRef] [PubMed]
- Arechiga Ceballos, N.; Vazquez Moron, S.; Berciano, J.M.; Nicolas, O.; Aznar Lopez, C.; Juste, J.; Rodriguez Nevado, C.; Aguilar Setien, A.; Echevarria, J.E. Novel lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef] [PubMed]
- Gunawardena, P.S.; Marston, D.A.; Ellis, R.J.; Wise, E.L.; Karawita, A.C.; Breed, A.C.; McElhinney, L.M.; Johnson, N.; Banyard, A.C.; Fooks, A.R. Lyssavirus in Indian Flying Foxes, Sri Lanka. Emerg. Infect. Dis. 2016, 22, 1456–1459. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; McElhinney, L.M.; Horton, D.; Banyard, A.C.; Johnson, N.; Marston, D.; Freuling, C.; Hoffmann, B.; Tu, C.; Fehlner-Gardiner, C.; et al. Molecular tools for rabies diagnosis in animals. In Proceedings of the Compendium of the OIE Global Conference on Rabies Control, Incheon-Seoul, Korea, 7–9 September 2011; OIE: Paris, France, 2012; pp. 75–85. [Google Scholar]
- Nokireki, T.; Tammiranta, N.; Kokkonen, U.M.; Kantala, T.; Gadd, T. Tentative novel lyssavirus in a bat in Finland. Transbound. Emerg. Dis. 2018, 65, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Afelt, A.; Frutos, R.; Devaux, C. Bats, Coronaviruses, and Deforestation: Toward the Emergence of Novel Infectious Diseases? Front. Microbiol. 2018, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Tordo, N. Host switching in Lyssavirus history from the Chiroptera to the Carnivora orders. J. Virol. 2001, 75, 8096–8104. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.E.; Barrett, J.; Briggs, D.; Cliquet, F.; Fooks, A.R.; Lumlertdacha, B.; Meslin, F.X.; Muler, T.; Nel, L.H.; Schneider, C.; et al. Can rabies be eradicated? Dev. Biol. (Basel) 2008, 131, 95–121. [Google Scholar] [PubMed]
- Lumio, J.; Hillbom, M.; Roine, R.; Ketonen, L.; Haltia, M.; Valle, M.; Neuvonen, E.; Lahdevirta, J. Human rabies of bat origin in Europe. Lancet 1986, 1, 378. [Google Scholar] [CrossRef]
- Fooks, A.R.; Brookes, S.M.; Johnson, N.; McElhinney, L.M.; Hutson, A.M. European bat lyssaviruses: An emerging zoonosis. Epidemiol. Infect. 2003, 131, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; McElhinney, L.M.; Pounder, D.J.; Finnegan, C.J.; Mansfield, K.; Johnson, N.; Brookes, S.M.; Parsons, G.; White, K.; McIntyre, P.G.; et al. Case report: Isolation of a European bat lyssavirus type 2a from a fatal human case of rabies encephalitis. J. Med. Virol. 2003, 71, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.; Cox, J.; Peter, W.; Schafer, R.; Johnson, N.; McElhinney, L.M.; Geue, J.L.; Tjornehoj, K.; Fooks, A.R. Spill-over of European bat lyssavirus type 1 into a stone marten (Martes foina) in Germany. J. Vet. Med. B Infect. Dis. Vet. Public Health 2004, 51, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Tjornehoj, K.; Fooks, A.R.; Agerholm, J.S.; Ronsholt, L. Natural and experimental infection of sheep with European bat lyssavirus type-1 of Danish bat origin. J. Comp. Pathol. 2006, 134, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, L.; Larrous, F.; Mailles, A.; Boisseleau, D.; Delmas, O.; Biron, C.; Bouchier, C.; Capek, I.; Muller, M.; Ilari, F.; et al. European bat Lyssavirus transmission among cats, Europe. Emerg. Infect. Dis. 2009, 15, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.; Johnson, N.; Freuling, C.M.; Fooks, A.R.; Selhorst, T.; Vos, A. Epidemiology of bat rabies in Germany. Arch. Virol. 2007, 152, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Serra-Cobo, J.; Amengual, B.; Abellan, C.; Bourhy, H. European bat lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Selimov, M.A.; Tatarov, A.G.; Botvinkin, A.D.; Klueva, E.V.; Kulikova, L.G.; Khismatullina, N.A. Rabies-related Yuli virus; identification with a panel of monoclonal antibodies. Acta Virol. 1989, 33, 542–546. [Google Scholar] [PubMed]
- Annand, E.; Reid, P. Clinical review of two fatal equine cases of infection with the insectivorous bat strain of Australian bat lyssavirus. Aust. Vet. J. 2014, 92, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Samaratunga, H.; Searle, J.W.; Hudson, N. Non-rabies Lyssavirus human encephalitis from fruit bats: Australian bat Lyssavirus (pteropid Lyssavirus) infection. Neuropathol. Appl. Neurobiol. 1998, 24, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.N.; Carney, I.K.; Smith, G.A.; Tannenberg, A.E.; Deverill, J.E.; Botha, J.A.; Serafin, I.L.; Harrower, B.J.; Fitzpatrick, P.F.; Searle, J.W. Australian bat lyssavirus infection: A second human case, with a long incubation period. Med. J. Aust. 2000, 172, 597–599. [Google Scholar] [PubMed]
- Iizuka, K. Australian Bat Lyssavirus—Human Fatality. Available online: http://www.promedmail.org/direct.php?id=20130323.1600266 (accessed on 12 November 2018).
- Familusi, J.B.; Moore, D.L. Isolation of a rabies related virus from the cerebrospinal fluid of a child with ‘aseptic meningitis’. Afr. J. Med. Sci. 1972, 3, 93–96. [Google Scholar] [PubMed]
- Familusi, J.; Osunkoya, B.; Moore, D.; Kemp, G.; Fabiyi, A. A fatal human infection with Mokola virus. Am. J. Trop. Med. Hyg. 1972, 21, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Foggin, C.M. Atypical rabies virus in cats and a dog in Zimbabwe. Vet. Rec. 1982, 110, 338. [Google Scholar] [CrossRef] [PubMed]
- Bingham, J.; Javangwe, S.; Sabeta, C.T.; Wandeler, A.I.; Nel, L.H. Report of isolations of unusual lyssaviruses (rabies and Mokola virus) identified retrospectively from Zimbabwe. J. S. Afr. Vet. Assoc. 2001, 72, 92–94. [Google Scholar] [CrossRef] [PubMed]
- Meredith, C.D.; Nel, L.H.; von Teichman, B.F. Further isolation of Mokola virus in South Africa. Vet. Rec. 1996, 138, 119–120. [Google Scholar] [PubMed]
- von Teichman, B.F.; de Koker, W.C.; Bosch, S.J.; Bishop, G.C.; Meredith, C.D.; Bingham, J. Mokola virus infection: Description of recent South African cases and a review of the virus epidemiology. J. S. Afr. Vet. Assoc. 1998, 69, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Sabeta, C.T.; Markotter, W.; Mohale, D.K.; Shumba, W.; Wandeler, A.I.; Nel, L.H. Mokola virus in domestic mammals, South Africa. Emerg. Infect. Dis. 2007, 13, 1371–1373. [Google Scholar] [CrossRef] [PubMed]
- Sabeta, C.; Blumberg, L.; Miyen, J.; Mohale, D.; Shumba, W.; Wandeler, A. Mokola virus involved in a human contact (South Africa). FEMS Immunol. Med. Microbiol. 2010, 58, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Mebatsion, T.; Cox, J.H.; Frost, J.W. Isolation and characterization of 115 street rabies virus isolates from Ethiopia by using monoclonal antibodies: Identification of 2 isolates as Mokola and Lagos bat viruses. J. Infect. Dis. 1992, 166, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Tignor, G.H.; Murphy, F.A.; Clark, H.F.; Shope, R.E.; Madore, P.; Bauer, S.P.; Buckley, S.M.; Meredith, C.D. Duvenhage Virus: Morphological, Biochemical, Histopathological and Antigenic Relationships to the Rabies Serogroup. J. Gen. Virol. 1977, 37, 595–611. [Google Scholar] [CrossRef]
- Paweska, J.T.; Blumberg, L.H.; Liebenberg, C.; Hewlett, R.H.; Grobbelaar, A.A.; Leman, P.A.; Croft, J.E.; Nel, L.H.; Nutt, L.; Swanepoel, R. Fatal human infection with rabies-related Duvenhage virus, South Africa. Emerg. Infect. Dis. 2006, 12, 1965–1967. [Google Scholar] [CrossRef] [PubMed]
- van Thiel, P.P.; de Bie, R.M.; Eftimov, F.; Tepaske, R.; Zaaijer, H.L.; van Doornum, G.J.; Schutten, M.; Osterhaus, A.D.; Majoie, C.B.; Aronica, E.; et al. Fatal human rabies due to Duvenhage virus from a bat in Kenya: Failure of treatment with coma-induction, ketamine, and antiviral drugs. PLoS Negl. Trop. Dis. 2009, 3, e428. [Google Scholar] [CrossRef] [PubMed]
- Crick, J.; Tignor, G.H.; Moreno, K. A new isolate of Lagos bat virus from the Republic of South Africa. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 211–213. [Google Scholar] [CrossRef]
- Markotter, W.; Kuzmin, I.; Rupprecht, C.E.; Nel, L.H. Phylogeny of Lagos bat virus: Challenges for lyssavirus taxonomy. Virus Res. 2008, 135, 10–21. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Crick, J. Rabies-related viruses. In Rabies; Springer: Berlin, Germany, 1988; pp. 177–199. [Google Scholar]
- Liu, Y.; Li, N.; Zhang, S.; Zhang, F.; Lian, H.; Wang, Y.; Zhang, J.; Hu, R. Analysis of the complete genome of the first Irkut virus isolate from China: Comparison across the Lyssavirus genus. Mol. Phylogenet. Evol. 2013, 69, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Miao, F.M.; Liu, Y.; Zhang, S.F.; Zhang, F.; Li, N.; Hu, R.L. Possible Transmission of Irkut Virus from Dogs to Humans. Biomed. Environ. Sci. 2018, 31, 146–148. [Google Scholar] [CrossRef] [PubMed]
- EUROBATS. Action Plan for the Conservation of Bat Species in the European Union 2016–2021; Inf.EUROBATS.AC21.5; EUROBATS: Bonn, Germany, 2006. [Google Scholar]
- Benda, P.; Abi-Said, M.; Bartonička, T.; Bilgin, R.; Faizolahi, K.; Lučan, R.K.; NICOLAOU, H.; REITER, A.; SHOHDI, W.M.; Uhrin, M. Rousettus aegyptiacus (Pteropodidae) in the Palaearctic: List of records and revision of the distribution range. Vespertilio 2011, 15, 3–36. [Google Scholar]
- Dietz, C.; Nill, D.; Von Helversen, O. Bats of Britain, Europe and Northwest Africa; A & C Black Publishers Ltd.: London, UK, 2009. [Google Scholar]
- Drexler, J.F.; Gloza-Rausch, F.; Glende, J.; Corman, V.M.; Muth, D.; Goettsche, M.; Seebens, A.; Niedrig, M.; Pfefferle, S.; Yordanov, S.; et al. Genomic characterization of severe acute respiratory syndrome-related coronavirus in European bats and classification of coronaviruses based on partial RNA-dependent RNA polymerase gene sequences. J. Virol. 2010, 84, 11336–11349. [Google Scholar] [CrossRef] [PubMed]
- Gouilh, M.A.; Puechmaille, S.J.; Diancourt, L.; Vandenbogaert, M.; Serra-Cobo, J.; Roïg, M.L.; Brown, P.; Moutou, F.; Caro, V.; Vabret, A.; et al. SARS-CoV related Betacoronavirus and diverse Alphacoronavirus members found in western old-world. Virology 2018, 517, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Pir, J.B.; Loesch, C.; Sausy, A.; Snoeck, C.J.; Hubschen, J.M.; Muller, C.P. Novel Alphacoronaviruses and Paramyxoviruses Cocirculate with Type 1 and Severe Acute Respiratory System (SARS)-Related Betacoronaviruses in Synanthropic Bats of Luxembourg. Appl. Environ. Microbiol. 2017, 83, e01326-17. [Google Scholar] [CrossRef] [PubMed]
- Rihtaric, D.; Hostnik, P.; Steyer, A.; Grom, J.; Toplak, I. Identification of SARS-like coronaviruses in horseshoe bats (Rhinolophus hipposideros) in Slovenia. Arch. Virol. 2010, 155, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Papetti, A.; Sabelli, C.; Rosti, E.; Moreno, A.; Boniotti, M.B. Detection of coronaviruses in bats of various species in Italy. Viruses 2013, 5, 2679–2689. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Moreno, A.; Lavazza, A.; Bresaola, M.; Canelli, E.; Boniotti, M.B.; Cordioli, P. Identification of Mammalian orthoreovirus type 3 in Italian bats. Zoonoses Public Health 2013, 60, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Dufkova, L.; Strakova, P.; Sirmarova, J.; Salat, J.; Moutelikova, R.; Chrudimsky, T.; Bartonicka, T.; Nowotny, N.; Ruzek, D. Detection of Diverse Novel Bat Astrovirus Sequences in the Czech Republic. Vector Borne Zoonotic Dis. 2015, 15, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Foldes, F.; Zana, B.; Kurucz, K.; Estok, P.; Boldogh, S.; Gorfol, T.; Banyai, K.; Oldal, M.; Jakab, F. Genetic Characterization of Providence Virus Isolated from Bat Guano in Hungary. Genome Announc. 2016, 4, e00403-16. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Moron, S.; Juste, J.; Ibanez, C.; Ruiz-Villamor, E.; Avellon, A.; Vera, M.; Echevarria, J.E. Endemic circulation of European bat lyssavirus type 1 in serotine bats, Spain. Emerg. Infect. Dis. 2008, 14, 1263–1266. [Google Scholar] [CrossRef] [PubMed]
- Falcon, A.; Vazquez-Moron, S.; Casas, I.; Aznar, C.; Ruiz, G.; Pozo, F.; Perez-Brena, P.; Juste, J.; Ibanez, C.; Garin, I.; et al. Detection of alpha and betacoronaviruses in multiple Iberian bat species. Arch. Virol. 2011, 156, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Dubourg-Savage, M.J.; Arthur, L.; Barataud, M.; Becu, D.; Bracco, S.; Borel, C.; Larcher, G.; Meme-Lafond, B.; Moinet, M.; et al. Active surveillance of bat rabies in France: A 5-year study (2004–2009). Vet. Microbiol. 2011, 151, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, S.; Ibanez, C.; Juste, J.; Echevarria, J.E. EBLV1 circulation in natural bat colonies of Eptesicus serotinus: A six year survey. Dev. Biol. (Basel) 2006, 125, 257–261. [Google Scholar] [PubMed]
- De Benedictis, P.; Marciano, S.; Scaravelli, D.; Priori, P.; Zecchin, B.; Capua, I.; Monne, I.; Cattoli, G. Alpha and lineage C betaCoV infections in Italian bats. Virus Genes 2014, 48, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Molnar, V.; Janoska, M.; Harrach, B.; Glavits, R.; Palmai, N.; Rigo, D.; Sos, E.; Liptovszky, M. Detection of a novel bat gammaherpesvirus in Hungary. Acta Vet. Hung. 2008, 56, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Perez, R.; Ibanez, C.; Godinez, J.M.; Arechiga, N.; Garin, I.; Perez-Suarez, G.; de Paz, O.; Juste, J.; Echevarria, J.E.; Bravo, I.G. Novel papillomaviruses in free-ranging Iberian bats: No virus-host co-evolution, no strict host specificity, and hints for recombination. Genome Biol. Evol. 2014, 6, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Roig, M.; Bourhy, H.; Lavenir, R.; Serra-Cobo, J. Seroprevalence dynamics of European bat lyssavirus type 1 in a multispecies bat colony. Viruses 2014, 6, 3386–3399. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Lelli, D.; de Sabato, L.; Zaccaria, G.; Boni, A.; Sozzi, E.; Prosperi, A.; Lavazza, A.; Cella, E.; Castrucci, M.R.; et al. Detection and full genome characterization of two beta CoV viruses related to Middle East respiratory syndrome from bats in Italy. Virol. J. 2017, 14, 239. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Muller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Cottontail, V.M.; Rasche, A.; Yordanov, S.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Gorfol, T.; Boldogh, S.; Estok, P.; Kurucz, K.; Oldal, M.; Nemeth, V.; Madai, M.; Banyai, K.; et al. Novel European lineages of bat astroviruses identified in Hungary. Acta Virol. 2014, 58, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Zeus, V.; Kwasnitschka, L.; Kerth, G.; Haase, M.; Groschup, M.H.; Balkema-Buschmann, A. Insectivorous bats carry host specific astroviruses and coronaviruses across different regions in Germany. Infect. Genet. Evol. 2016, 37, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Gottsche, M.; Panning, M.; Drexler, J.F.; Petersen, N.; Annan, A.; Grywna, K.; Muller, M.; et al. Detection and prevalence patterns of group I coronaviruses in bats, northern Germany. Emerg. Infect. Dis. 2008, 14, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.T.; Kuzmin, I.V.; Kameoka, Y.; Botvinkin, A.D. New lyssavirus genotype from the Lesser Mouse-eared Bat (Myotis blythi), Kyrghyzstan. Emerg. Infect. Dis. 2003, 9, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Serra-Cobo, J.; Lopez-Roig, M.; Segui, M.; Sanchez, L.P.; Nadal, J.; Borras, M.; Lavenir, R.; Bourhy, H. Ecological factors associated with European bat lyssavirus seroprevalence in spanish bats. PLoS ONE 2013, 8, e64467. [Google Scholar] [CrossRef] [PubMed]
- Van der Poel, W.H.; Van der Heide, R.; Verstraten, E.R.; Takumi, K.; Lina, P.H.; Kramps, J.A. European bat lyssaviruses, The Netherlands. Emerg. Infect. Dis. 2005, 11, 1854–1859. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C.M.; Kliemt, J.; Schares, S.; Heidecke, D.; Driechciarz, R.; Schatz, J.; Muller, T. Detection of European bat lyssavirus 2 (EBLV-2) in a Daubenton’s bat (Myotis daubentonii) from Magdeburg, Germany. Berliner und Munchener Tierarztliche Wochenschrift 2012, 125, 255–258. [Google Scholar] [PubMed]
- Davis, P.L.; Holmes, E.C.; Larrous, F.; Van der Poel, W.H.; Tjornehoj, K.; Alonso, W.J.; Bourhy, H. Phylogeography, population dynamics, and molecular evolution of European bat lyssaviruses. J. Virol. 2005, 79, 10487–10497. [Google Scholar] [CrossRef] [PubMed]
- Nokireki, T.; Sironen, T.; Smura, T.; Karkamo, V.; Sihvonen, L.; Gadd, T. Second case of European bat lyssavirus type 2 detected in a Daubenton’s bat in Finland. Acta Vet. Scand. 2017, 59, 62. [Google Scholar] [CrossRef] [PubMed]
- Moldal, T.; Vikoren, T.; Cliquet, F.; Marston, D.A.; van der Kooij, J.; Madslien, K.; Orpetveit, I. First detection of European bat lyssavirus type 2 (EBLV-2) in Norway. BMC Vet. Res. 2017, 13, 216. [Google Scholar] [CrossRef] [PubMed]
- Hammarin, A.L.; Berndtsson, L.T.; Falk, K.; Nedinge, M.; Olsson, G.; Lundkvist, A. Lyssavirus-reactive antibodies in Swedish bats. Infect. Ecol. Epidemiol. 2016, 6, 31262. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.L.; Mansfield, K.; Marston, D.A.; Johnson, N.; Pajamo, K.; O’Brien, N.; Black, C.; McElhinney, L.M.; Fooks, A.R. Isolation of European bat lyssavirus type 2 from a Daubenton’s bat (Myotis daubentonii) in Shropshire. Vet. Rec. 2007, 161, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Amengual, B.; Bourhy, H.; Lopez-Roig, M.; Serra-Cobo, J. Temporal dynamics of European bat Lyssavirus type 1 and survival of Myotis myotis bats in natural colonies. PLoS ONE 2007, 2, e566. [Google Scholar] [CrossRef] [PubMed]
- Wibbelt, G.; Kurth, A.; Yasmum, N.; Bannert, M.; Nagel, S.; Nitsche, A.; Ehlers, B. Discovery of herpesviruses in bats. J. Gen. Virol. 2007, 88, 2651–2655. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Orciari, L.A.; Arai, Y.T.; Smith, J.S.; Hanlon, C.A.; Kameoka, Y.; Rupprecht, C.E. Bat lyssaviruses (Aravan and Khujand) from Central Asia: Phylogenetic relationships according to N, P and G gene sequences. Virus Res. 2003, 97, 65–79. [Google Scholar] [CrossRef]
- Dacheux, L.; Cervantes-Gonzalez, M.; Guigon, G.; Thiberge, J.M.; Vandenbogaert, M.; Maufrais, C.; Caro, V.; Bourhy, H. A preliminary study of viral metagenomics of French bat species in contact with humans: identification of new mammalian viruses. PLoS ONE 2014, 9, e87194. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Lesnik, R.; Brinkmann, A.; Ebinger, A.; Radonic, A.; Nitsche, A.; Muhldorfer, K.; Wibbelt, G.; Kurth, A. Isolation and characterization of three mammalian orthoreoviruses from European bats. PLoS ONE 2012, 7, e43106. [Google Scholar] [CrossRef] [PubMed]
- Kurth, A.; Kohl, C.; Brinkmann, A.; Ebinger, A.; Harper, J.A.; Wang, L.F.; Muhldorfer, K.; Wibbelt, G. Novel paramyxoviruses in free-ranging European bats. PLoS ONE 2012, 7, e38688. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Servat, A.; Robardet, E.; Moinet, M.; Borel, C.; Cliquet, F. Isolation of Bokeloh bat lyssavirus in Myotis nattereri in France. Arch. Virol. 2013, 158, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C.M.; Beer, M.; Conraths, F.J.; Finke, S.; Hoffmann, B.; Keller, B.; Kliemt, J.; Mettenleiter, T.C.; Muhlbach, E.; Teifke, J.P.; et al. Novel lyssavirus in Natterer’s bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–1522. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, F.; Edenborough, K.M.; Toffoli, R.; Culasso, P.; Zoppi, S.; Dondo, A.; Robetto, S.; Rosati, S.; Lander, A.; Kurth, A.; et al. Coronavirus and paramyxovirus in bats from Northwest Italy. BMC Vet. Res. 2017, 13, 396. [Google Scholar] [CrossRef] [PubMed]
- Schatz, J.; Freuling, C.M.; Auer, E.; Goharriz, H.; Harbusch, C.; Johnson, N.; Kaipf, I.; Mettenleiter, T.C.; Muhldorfer, K.; Muhle, R.U.; et al. Enhanced passive bat rabies surveillance in indigenous bat species from Germany—A retrospective study. PLoS Negl. Trop. Dis. 2014, 8, e2835. [Google Scholar] [CrossRef] [PubMed]
- Strakova, P.; Dufkova, L.; Sirmarova, J.; Salat, J.; Bartonicka, T.; Klempa, B.; Pfaff, F.; Hoper, D.; Hoffmann, B.; Ulrich, R.G.; et al. Novel hantavirus identified in European bat species Nyctalus noctula. Infect. Genet. Evol. 2017, 48, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Janoska, M.; Vidovszky, M.; Molnar, V.; Liptovszky, M.; Harrach, B.; Benko, M. Novel adenoviruses and herpesviruses detected in bats. Vet. J. 2011, 189, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Prosperi, A.; Moreno, A.; Chiapponi, C.; Gibellini, A.M.; De Benedictis, P.; Leopardi, S.; Sozzi, E.; Lavazza, A. Isolation of a novel Rhabdovirus from an insectivorous bat (Pipistrellus kuhlii) in Italy. Virol. J. 2018, 15, 37. [Google Scholar] [CrossRef] [PubMed]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.; Amaducci, L.; Coluzzi, M.; et al. Ecology of viruses isolated from sand flies in Italy and characterized of a new Phlebovirus (Arabia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Vidovszky, M.Z.; Muhldorfer, K.; Dabrowski, P.W.; Radonic, A.; Nitsche, A.; Wibbelt, G.; Kurth, A.; Harrach, B. Genome analysis of bat adenovirus 2: Indications of interspecies transmission. J. Virol. 2012, 86, 1888–1892. [Google Scholar] [CrossRef] [PubMed]
- Eggerbauer, E.; Troupin, C.; Passior, K.; Pfaff, F.; Hoper, D.; Neubauer-Juric, A.; Haberl, S.; Bouchier, C.; Mettenleiter, T.C.; Bourhy, H.; et al. The Recently Discovered Bokeloh Bat Lyssavirus: Insights Into Its Genetic Heterogeneity and Spatial Distribution in Europe and the Population Genetics of Its Primary Host. Adv. Virus Res. 2017, 99, 199–232. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, M.; Muhldorfer, K.; Speck, S.; Wibbelt, G.; Kurth, A. New adenovirus in bats, Germany. Emerg. Infect. Dis. 2009, 15, 2052–2055. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.B.; Lina, P.H.; Pielaat, A.; de Vries, A.; Dam-Deisz, C.; Adema, J.; Drexler, J.F.; Drosten, C.; Kooi, E.A. Circulation of group 2 coronaviruses in a bat species common to urban areas in Western Europe. Vector Borne Zoonotic Dis. 2010, 10, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Gorfol, T.; Boldogh, S.; Estok, P.; Kurucz, K.; Kutas, A.; Foldes, F.; Oldal, M.; Nemeth, V.; et al. Molecular survey of RNA viruses in Hungarian bats: Discovering novel astroviruses, coronaviruses, and caliciviruses. Vector Borne Zoonotic Dis. 2014, 14, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Selimov, M.A.; Smekhov, A.M.; Antonova, L.A.; Shablovskaya, E.A.; King, A.A.; Kulikova, L.G. New strains of rabies-related viruses isolated from bats in the Ukraine. Acta Virol. 1991, 35, 226–231. [Google Scholar] [PubMed]
- Negredo, A.; Palacios, G.; Vazquez-Moron, S.; Gonzalez, F.; Dopazo, H.; Molero, F.; Juste, J.; Quetglas, J.; Savji, N.; de la Cruz Martinez, M.; et al. Discovery of an ebolavirus-like filovirus in europe. PLoS Pathog. 2011, 7, e1002304. [Google Scholar] [CrossRef] [PubMed]
- Wellenberg, G.J.; Audry, L.; Ronsholt, L.; van der Poel, W.H.; Bruschke, C.J.; Bourhy, H. Presence of European bat lyssavirus RNas in apparently healthy Rousettus aegyptiacus bats. Arch. Virol. 2002, 147, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Towner, J.S.; Amman, B.R.; Sealy, T.K.; Carroll, S.A.; Comer, J.A.; Kemp, A.; Swanepoel, R.; Paddock, C.D.; Balinandi, S.; Khristova, M.L.; et al. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog. 2009, 5, e1000536. [Google Scholar] [CrossRef] [PubMed]
- Chowell, G.; Nishiura, H. Transmission dynamics and control of Ebola virus disease (EVD): A review. BMC Med. 2014, 12, 196. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, I.; Amarasinghe, G.K.; Basler, C.F. Filovirus pathogenesis and immune evasion: Insights from Ebola virus and Marburg virus. Nat. Rev. Microbiol. 2015, 13, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.L.; Dhanota, J.K.; Liang, E.; et al. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Kurucz, K.; Dallos, B.; Zana, B.; Földes, F.; Boldogh, S.; Görföl, T.; Carroll, M.W.; Jakab, F. Re-emergence of Lloviu virus in Miniopterus schreibersii bats, Hungary, 2016. Emerg. Microbes Infect. 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Quetglas, J.; González, F.; Paz, Ó.D. Estudian la extraña mortandad de miles de murciélagos en cueva. Quercus 2003, 203, 50–51. [Google Scholar]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Délicat, A.; Paweska, J.T.; Gonzalez, J.-P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R.; Leman, P.A.; Burt, F.J.; Zachariades, N.A.; Braack, L.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Peters, C.J. Experimental inoculation of plants and animals with Ebola virus. Emerg. Infect. Dis. 1996, 2, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Azhar, E.I.; Lanini, S.; Ippolito, G.; Zumla, A. The Middle East Respiratory Syndrome Coronavirus—A Continuing Risk to Global Health Security. Adv. Exp. Med. Biol. 2017, 972, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Poon, L.L.; Chu, D.K.; Chan, K.H.; Wong, O.K.; Ellis, T.M.; Leung, Y.H.; Lau, S.K.; Woo, P.C.; Suen, K.Y.; Yuen, K.Y.; et al. Identification of a novel coronavirus in bats. J. Virol. 2005, 79, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Ge, X.; Wang, L.F.; Shi, Z. Bat origin of human coronaviruses. Virol. J. 2015, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Mishra, N.; Olival, K.J.; Fagbo, S.F.; Kapoor, V.; Epstein, J.H.; Alhakeem, R.; Durosinloun, A.; Al Asmari, M.; Islam, A.; et al. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg. Infect. Dis. 2013, 19, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Prevalence of IgG antibody to SARS-associated coronavirus in animal traders--Guangdong Province, China, 2003. MMWR Morb. Mortal. Wkly. Rep. 2003, 52, 986–987. [Google Scholar]
- Xu, H.F.; Wang, M.; Zhang, Z.B.; Zou, X.Z.; Gao, Y.; Liu, X.N.; Lu, E.J.; Pan, B.Y.; Wu, S.J.; Yu, S.Y. [An epidemiologic investigation on infection with severe acute respiratory syndrome coronavirus in wild animals traders in Guangzhou]. Zhonghua Yu Fang Yi Xue Za Zhi 2004, 38, 81–83. [Google Scholar] [PubMed]
- Kan, B.; Wang, M.; Jing, H.; Xu, H.; Jiang, X.; Yan, M.; Liang, W.; Zheng, H.; Wan, K.; Liu, Q.; et al. Molecular evolution analysis and geographic investigation of severe acute respiratory syndrome coronavirus-like virus in palm civets at an animal market and on farms. J. Virol. 2005, 79, 11892–11900. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Hu, Z. A review of studies on animal reservoirs of the SARS coronavirus. Virus Res. 2008, 133, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.J.; Ojeda-Flores, R.; Rico-Chávez, O.; Navarrete-Macias, I.; Zambrana-Torrelio, C.M.; Rostal, M.K.; Epstein, J.H.; Tipps, T.; Liang, E.; Sanchez-Leon, M.; et al. Coronaviruses in bats from Mexico. J. Gen. Virol. 2013, 94, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Masters, P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, T.M.; Buchmeier, M.J. Coronavirus spike proteins in viral entry and pathogenesis. Virology 2001, 279, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Song, H.D.; Tu, C.C.; Zhang, G.W.; Wang, S.Y.; Zheng, K.; Lei, L.C.; Chen, Q.X.; Gao, Y.W.; Zhou, H.Q.; Xiang, H.; et al. Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human. Proc. Natl. Acad. Sci. USA 2005, 102, 2430–2435. [Google Scholar] [CrossRef] [PubMed]
- Nadin-Davis, S.; Alnabelseya, N.; Knowles, M.K. The phylogeography of Myotis bat-associated rabies viruses across Canada. PLoS Negl. Trop. Dis. 2017, 11, e0005541. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.J.; Orciari, L.A.; Rupprecht, C.E. Evolutionary timescale of rabies virus adaptation to North American bats inferred from the substitution rate of the nucleoprotein gene. J. Gen. Virol. 2005, 86, 1467–1474. [Google Scholar] [CrossRef] [PubMed]
- Kuzmina, N.A.; Kuzmin, I.V.; Ellison, J.A.; Taylor, S.T.; Bergman, D.L.; Dew, B.; Rupprecht, C.E. A reassessment of the evolutionary timescale of bat rabies viruses based upon glycoprotein gene sequences. Virus Genes 2013, 47, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.T.; Petersen, B.W.; Recuenco, S.; Niezgoda, M.; Gomez, J.; Laguna-Torres, V.A.; Rupprecht, C. Evidence of rabies virus exposure among humans in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2012, 87, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Pajamo, K.; Harkess, G.; Goddard, T.; Marston, D.; McElhinney, L.; Johnson, N.; Fooks, A.R. Isolation of European bat lyssavirus type 2 (EBLV-2) in a Daubenton’s bat in the UK with a minimum incubation period of 9 months. Rabies Bull. Eur. 2008, 32, 6–7. [Google Scholar]
- Warrell, M.J. Current rabies vaccines and prophylaxis schedules: Preventing rabies before and after exposure. Travel Med. Infect. Dis. 2012, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Brookes, S.M.; Healy, D.M.; Fooks, A.R. Ability of rabies vaccine strains to elicit cross-neutralising antibodies. Dev. Biol. (Basel) 2006, 125, 185–193. [Google Scholar] [PubMed]
- Brookes, S.M.; Parsons, G.; Johnson, N.; McElhinney, L.M.; Fooks, A.R. Rabies human diploid cell vaccine elicits cross-neutralising and cross-protecting immune responses against European and Australian bat lyssaviruses. Vaccine 2005, 23, 4101–4109. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, C.A.; Kuzmin, I.V.; Blanton, J.D.; Weldon, W.C.; Manangan, J.S.; Rupprecht, C.E. Efficacy of rabies biologics against new lyssaviruses from Eurasia. Virus Res. 2005, 111, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.S.; Horton, D.L.; Easton, A.J.; Fooks, A.R.; Banyard, A.C. Rabies virus vaccines: Is there a need for a pan-lyssavirus vaccine? Vaccine 2012, 30, 7447–7454. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; McElhinney, L.M.; Marston, D.A.; Wood, J.L.; Russell, C.A.; Lewis, N.; Kuzmin, I.V.; Fouchier, R.A.; Osterhaus, A.D.; Fooks, A.R.; et al. Quantifying antigenic relationships among the Lyssaviruses. J. Virol. 2010, 84, 11841–11848. [Google Scholar] [CrossRef] [PubMed]
- Nolden, T.; Banyard, A.C.; Finke, S.; Fooks, A.R.; Hanke, D.; Hoper, D.; Horton, D.L.; Mettenleiter, T.C.; Muller, T.; Teifke, J.P.; et al. Comparative studies on the genetic, antigenic and pathogenic characteristics of Bokeloh bat lyssavirus. J. Gen. Virol. 2014, 95, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.M.; Hanlon, C.A. Rabies-specific antibodies: Measuring surrogates of protection against a fatal disease. PLoS Negl. Trop. Dis. 2010, 4, e595. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; Banyard, A.C.; Marston, D.A.; Wise, E.; Selden, D.; Nunez, A.; Hicks, D.; Lembo, T.; Cleaveland, S.; Peel, A.J.; et al. Antigenic and genetic characterization of a divergent African virus, Ikoma lyssavirus. J. Gen. Virol. 2014, 95, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.S.; Selden, D.; Wu, G.; Wright, E.; Horton, D.L.; Fooks, A.R.; Banyard, A.C. Antigenic site changes in the rabies virus glycoprotein dictates functionality and neutralizing capability against divergent lyssaviruses. J. Gen. Virol. 2018, 99, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Possible emergence of West Caucasian bat virus in Africa. Emerg. Infect. Dis. 2008, 14, 1887–1889. [Google Scholar] [CrossRef] [PubMed]
- Marzi, A.; Hanley, P.W.; Haddock, E.; Martellaro, C.; Kobinger, G.; Feldmann, H. Efficacy of Vesicular Stomatitis Virus-Ebola Virus Postexposure Treatment in Rhesus Macaques Infected With Ebola Virus Makona. J. Infect. Dis. 2016, 214, S360–S366. [Google Scholar] [CrossRef] [PubMed]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef]
- Mire, C.E.; Geisbert, J.B.; Agans, K.N.; Satterfield, B.A.; Versteeg, K.M.; Fritz, E.A.; Feldmann, H.; Hensley, L.E.; Geisbert, T.W. Durability of a Vesicular Stomatitis Virus-Based Marburg Virus Vaccine in Nonhuman Primates. PLoS ONE 2014, 9, e94355. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Becker, S.; Ebihara, H.; Geisbert, T.W.; Johnson, K.M.; Kawaoka, Y.; Lipkin, W.I.; Negredo, A.I.; Netesov, S.V.; Nichol, S.T.; et al. Proposal for a revised taxonomy of the family Filoviridae: Classification, names of taxa and viruses, and virus abbreviations. Arch. Virol. 2010, 155, 2083–2103. [Google Scholar] [CrossRef] [PubMed]
- Okba, N.M.A.; Raj, V.S.; Haagmans, B.L. Middle East respiratory syndrome coronavirus vaccines: Current status and novel approaches. Curr. Opin. Virol. 2017, 23, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Li, Y.; Wang, L.; Zhao, G.; Tao, X.; Tseng, C.-T.K.; Zhou, Y.; Du, L.; Jiang, S. Intranasal vaccination with recombinant receptor-binding domain of MERS-CoV spike protein induces much stronger local mucosal immune responses than subcutaneous immunization: Implication for designing novel mucosal MERS vaccines. Vaccine 2014, 32, 2100–2108. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhao, J.; Mangalam, A.K.; Channappanavar, R.; Fett, C.; Meyerholz, D.K.; Agnihothram, S.; Baric, R.S.; David, C.S.; Perlman, S. Airway Memory CD4+ T Cells Mediate Protective Immunity against Emerging Respiratory Coronaviruses. Immunity 2016, 44, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Stading, B.; Ellison, J.A.; Carson, W.C.; Satheshkumar, P.S.; Rocke, T.E.; Osorio, J.E. Protection of bats (Eptesicus fuscus) against rabies following topical or oronasal exposure to a recombinant raccoon poxvirus vaccine. PLoS Negl. Trop. Dis. 2017, 11, e0005958. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Lyssavirus Species | Common Bat Name | Bat Species Associated with Lyssavirus Infection | Countries Reporting Lyssavirus in Bats | Vaccine Protection Predicted? |
|---|---|---|---|---|
| Aravan lyssavirus (ARAV) | Lesser mouse-eared bat | Myotis blythi | Kyrgystan | Y |
| Australian bat lyssavirus (ABLV) | Black flying fox and related sp. | Pteropus alecto | Australia | Y |
| Yellow-bellied sheath-tailed bat | Saccolaimus flaviventris | |||
| Bokeloh bat lyssavirus (BBLV) | Natterer’s bat | Myotis nattereri | Germany, France | Y |
| Duvenhage lyssavirus (DUVV) | Undefined | Miniopterus sp. | South Africa, Kenya | Y |
| Egyptian slit-faced bat | Nycteris thebaica | Zimbabwe | ||
| European bat 1 Lyssavirus (EBLV-1) | Serotine bat | Eptesicus serotinus | France, Germany, and Spain | Y |
| European bat 2 lyssavirus (EBLV-2) | Daubenton’s bat | Myotis daubentonii | The Netherlands, Switzerland, UK, France, Germany, Luxembourg, and Finland | Y |
| Gannoruwa bat lyssavirus (GBLV) | Indian flying fox | Pteropus medius | Sri Lanka | Y |
| Ikoma lyssavirus (IKOV) | N/A | N/A | Tanzania | N |
| Irkut lyssavirus (IRKV) | Greater tube-nosed bat | Murina leucogaster | Russian Federation and China | Y |
| Kotolahti Bat Lyssavirus (KBLV)$ | Brandt’s bat | Myotis brandti | Finland | Y |
| Khujand lyssavirus (KHUV) | Whiskered bat | Myotis mystacinus | Tajikistan | Y |
| Lagos bat lyssavirus (LBV) | Straw coloured fruit bat | Eidolon helvum | Nigeria, Senegal, Ghana, and Kenya | N |
| Egyptian fruit bat | Rousettus aegyptiacus | France (ex-Togo or Egypt), and Kenya | ||
| Dwarf epaulet fruit bat | Micropteropus pussilus | Central African Republic | ||
| Gambian epauletted fruit bat | Epomorphus giambianus | Ghana | ||
| Buettikofer’s epauletted fruit bat | Epomops buettikoferi | Ghana | ||
| Gambian slit-faced bat | Nycteris gambiensis | Guinea | ||
| Wahlberg’s epauletted fruit bat | Epomorphorus wahlbergi | South Africa | ||
| Lleida bat lyssavirus (LLEBV) | Common bent-winged bat | Miniopterus schreibersii | Spain and France | N |
| Mokola Lyssavirus (MOKV) | N/A | N/A | N | |
| Rabies lyssavirus (RABV)○ | Big brown bat | Eptesicus fuscus | North and South America | Y |
| Mexican/Brazilian free-tail bat | Tadarida brasiliensis | |||
| Silver-haired bat | Lasionycteris noctivagens | |||
| Tri-coloured bat | Perimyotis subflavus | |||
| Vampire bat | Desmodus rotundus | |||
| Shimoni bat lyssavirus (SHIBV) | Commerson’s leaf-nosed bat | Hipposideros commersoni | Kenya | N |
| Taiwan bat lyssavirus (TBLV)$ | Japanese house bat | Pipistrellus abramus | Taiwan | Y |
| West Caucasian bat lyssavirus (WCBV) | Common bent-winged bat | Miniopterus schreibersii | Russian Federation and Kenya^ | N |
| Continent | Lyssavirus Species | Country | Presumed Host Species | Spill over Species | Reference |
|---|---|---|---|---|---|
| Europe | EBLV-1 | Germany | Eptesicus serotinus | Stone marten ×1 | [33] |
| Denmark | Eptesicus serotinus | Sheep ×4 (1998), 1 × (2002) | [34] | ||
| France | Eptesicus serotinus | Cat ×1 (2003), 1 × Cat (2007) | [35] | ||
| Germany | Eptesicus serotinus | Myotis myotis | [36] | ||
| Eptesicus serotinus | Myotis daubentonii | ||||
| Eptesicus serotinus | Nytalus noctula | ||||
| Eptesicus serotinus | Pipistrellus pipistrellus | ||||
| Eptesicus serotinus | Pipistrellus nathusii | ||||
| Eptesicus serotinus | Plecotus auritus | ||||
| Spain | Eptesicus serotinus | Myotis nattereri | [37] | ||
| Eptesicus serotinus | Minipterus schreibersii | ||||
| Eptesicus serotinus | Rhinolophus ferrumequinum | ||||
| Eptesicus serotinus | Barbastella barbastellus | ||||
| Russia | Eptesicus serotinus | Human | [38] | ||
| EBLV-2 | Finland | Myotis daubentonii | Human | [30] | |
| Scotland | Myotis daubentonii | Human | [32] | ||
| Oceania | ABLV | Australia | Chalinolobus gouldii | 2× Horses | [39] |
| Saccolaimus flaviventris | Human | [40] | |||
| Pteropus spp. | Human | [41] | |||
| Pteropus spp. | Human | [42] | |||
| Africa | MOKV | Nigeria | Unknown | Human | [43] |
| Unknown | Human | [44] | |||
| Zimbabwe | Unknown | 5 cats; 1 dog | [45] | ||
| Unknown | 1 cat | [46] | |||
| South Africa | Unknown | Cat ×1 (1970); Cat ×1 (1995) | [47] | ||
| Unknown | cat × 2 (1996), cat ×2 (1997), cat ×1 (1998) | [48] | |||
| Unknown | 1 cat | [49] | |||
| Unknown | dog ×1 (2005), cat ×1 (2006), cat ×1 (2008) | [50] | |||
| Unknown | cat ×2 (2012), cat ×1 (2014) | [13] | |||
| Ethiopia | Unknown | 1 cat | [51] | ||
| DUVV | South Africa | Microchiroptera | Human | [52] | |
| Microchiroptera | Human | [53] | |||
| Kenya | Microchiroptera | Human | [54] | ||
| LBV | South Africa | Epomophorus wahlbergi | Cat | [55] | |
| Epomophorus wahlbergi | Mongoose | [56] | |||
| Zimbabwe | Epomophorus wahlbergi/Eidolon helvum | Cat | [57] | ||
| Ethiopia | Epomophorus wahlbergi/Eidolon helvum | Dog | [51] | ||
| Asia | IRKV | Siberia | Murina leucogaster | Human | [58] |
| China | Murina leucogaster | Dog | [59] |
| Family | Species Common Name | Species Latin Name | Association with Viral Pathogen? | References |
|---|---|---|---|---|
| Rhinolophidae | Blasius’s horseshoe bat | Rhinolophus blasii | Coronaviruses | [63] |
| Mediterranean horseshoe bat | Rhinolophus euryale | Coronaviruses | [63] | |
| Greater horseshoe bat | Rhinolophus ferrumequinum | Coronaviruses/Gammaherpesvirus/Adenovirus/Papillomaviruses | [37,63,64,65] | |
| Lesser horseshoe bat | Rhinolophus hipposideros | Coronaviruses/Orthoreovirus/Astrovirus | [66,67,68,69] | |
| Mehely’s horseshoe bat | Rhinolophus mehelyi | Coronavirus | [63] | |
| Vespertilionidae | Western Barbastelle bat | Barbastella barbastellus | Carmovirus | [70] |
| Isabelline Serotine bat | Eptesicus isabellinus | Coronavirus | [71,72] | |
| Common Serotine | Eptesicus serotinus | Coronavirus/Herpesvirus/Papillomavirus | [36,73,74,75,76,77] | |
| Savi’s pipistrelle | Hypsugo savii | Coronavirus | [67,78,79] | |
| Alcathoe whiskered bat | Myotis alcathoe | Paramyxovirus | [80] | |
| Bechstein’s bat | Myotis bechsteinii | Astrovirus/Coronavirus/Paramyxovirus | [81,82,83] | |
| Lesser mouse-eared bat | Myotis blythii | Coronavirus | [72,75,84] | |
| Long-fingered bat | Myotis capaccinii | Paramyxovirus | [80,85] | |
| Pond bat | Myotis dasycneme | Coronavirus | [83,86,87,88] | |
| Daubenton’s bat | Myotis daubentonii | Atrovirus/Coronavirus/Paramyxovirus | [63,80,81,83,85,87,89,90,91,92] | |
| Geoffroy’s bat | Myotis emarginatus | Paramyxovirus/Coronavirus | [65] | |
| Greater mouse-eared bat | Myotis myotis | Coronavirus/Herpesvirus/Paramyxovirus | [72,73,80,93,94] | |
| Whiskered bat | Myotis mystacinus | Bunyavirus/Reovirus/Paramyxovirus | [95,96,97,98] | |
| Natterer’s bat | Myotis nattereri | Coronavirus/Herpesvirus | [94,99,100,101] | |
| Greater noctule bat | Nyctalus lasiopterus | Coronavirus | [72] | |
| Leisler’s bat | Nyctalus leisleri | Coronavirus | [63] | |
| Common noctule | Nyctalus noctula | Hantavirus/Coronavirus/Adenovirus | [67,102,103,104] | |
| Kuhl’s pipistrelle | Pipistrellus kuhlii | Rhabdovirus/Coronavirus/Bunyavirus/Reovirus | [79,105,106] | |
| Nathusius’s pipistrelle | Pipistrellus nathusii | Adenovirus/Coronavirus/Reovirus | [83,97,102,107] | |
| Common pipistrelle | Pipistrellus pipistrellus | Adenovirus/Coronavirus/Herpesvirus/Paramyxovirus | [78,94,102,108,109,110] | |
| Pygmy pipistrelle | Pipistrellus pygmaeus | Astrovirus/Coronavirus | [111] | |
| Brown long-eared bat | Plecotus auritus | Coronavirus/Astrovirus/Herpesvirus/Reovirus | [81,94,101,102] | |
| Parti-coloured bat | Vespertilio murinus | Reovirus/Astrovirus | [68,69,112] | |
| Miniopteridae | Schreiber’s bat | Miniopterus schreibersii | Lloviu (filovirus) | [23,113] |
| Molossidae | European free-tailed bat | Tadarida teniotis | Mammalian orthoreovirus (reovirus) | [68,78] |
| Pteropodidae | Egyptian fruit bat | Rousettus aegyptiacus | Marburg and Ravn (filovirus) | [114,115] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shipley, R.; Wright, E.; Selden, D.; Wu, G.; Aegerter, J.; Fooks, A.R.; Banyard, A.C. Bats and Viruses: Emergence of Novel Lyssaviruses and Association of Bats with Viral Zoonoses in the EU. Trop. Med. Infect. Dis. 2019, 4, 31. https://doi.org/10.3390/tropicalmed4010031
Shipley R, Wright E, Selden D, Wu G, Aegerter J, Fooks AR, Banyard AC. Bats and Viruses: Emergence of Novel Lyssaviruses and Association of Bats with Viral Zoonoses in the EU. Tropical Medicine and Infectious Disease. 2019; 4(1):31. https://doi.org/10.3390/tropicalmed4010031
Chicago/Turabian StyleShipley, Rebecca, Edward Wright, David Selden, Guanghui Wu, James Aegerter, Anthony R Fooks, and Ashley C Banyard. 2019. "Bats and Viruses: Emergence of Novel Lyssaviruses and Association of Bats with Viral Zoonoses in the EU" Tropical Medicine and Infectious Disease 4, no. 1: 31. https://doi.org/10.3390/tropicalmed4010031
APA StyleShipley, R., Wright, E., Selden, D., Wu, G., Aegerter, J., Fooks, A. R., & Banyard, A. C. (2019). Bats and Viruses: Emergence of Novel Lyssaviruses and Association of Bats with Viral Zoonoses in the EU. Tropical Medicine and Infectious Disease, 4(1), 31. https://doi.org/10.3390/tropicalmed4010031