Drug Discovery for Chagas Disease: Impact of Different Host Cell Lines on Assay Performance and Hit Compound Selection

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
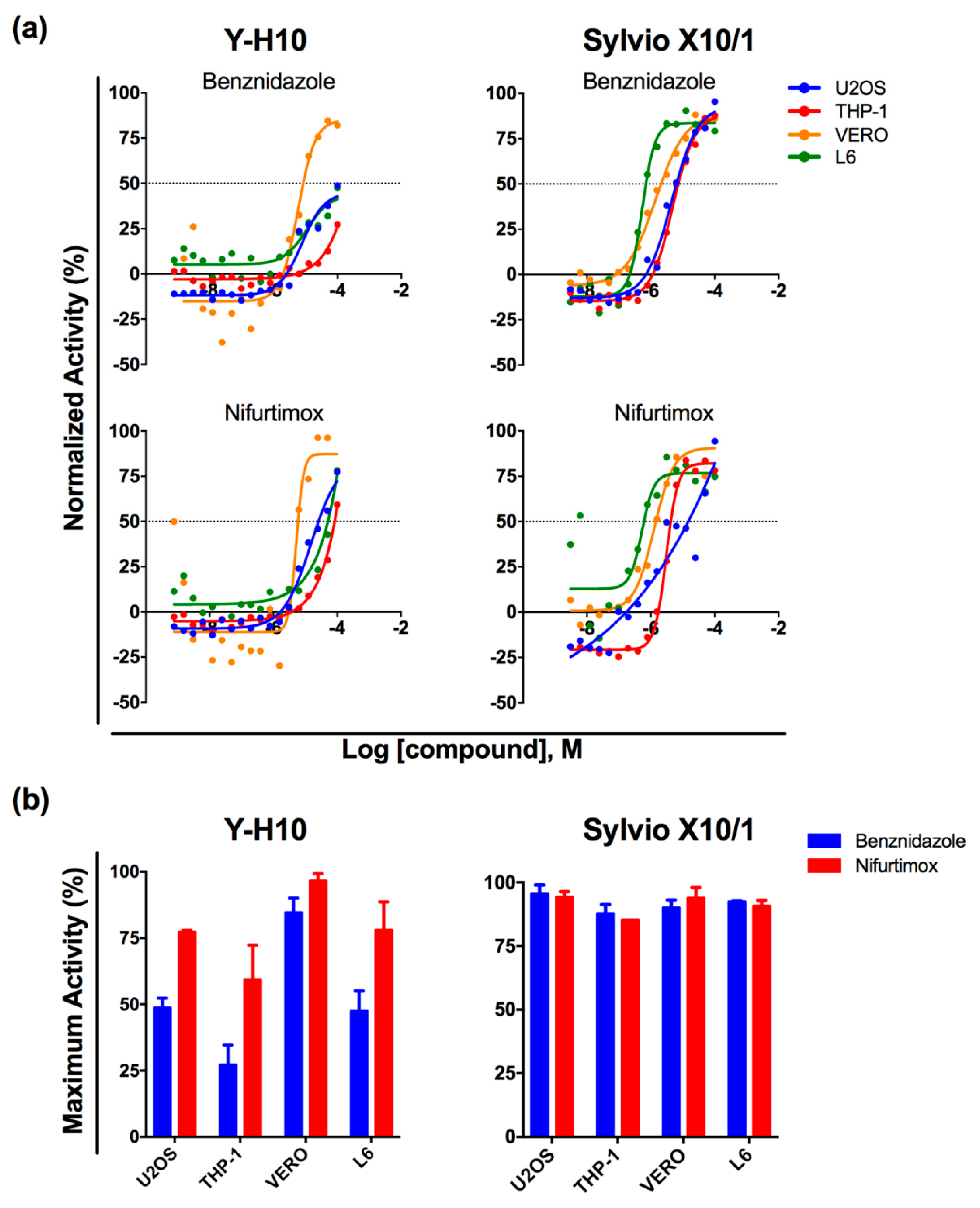
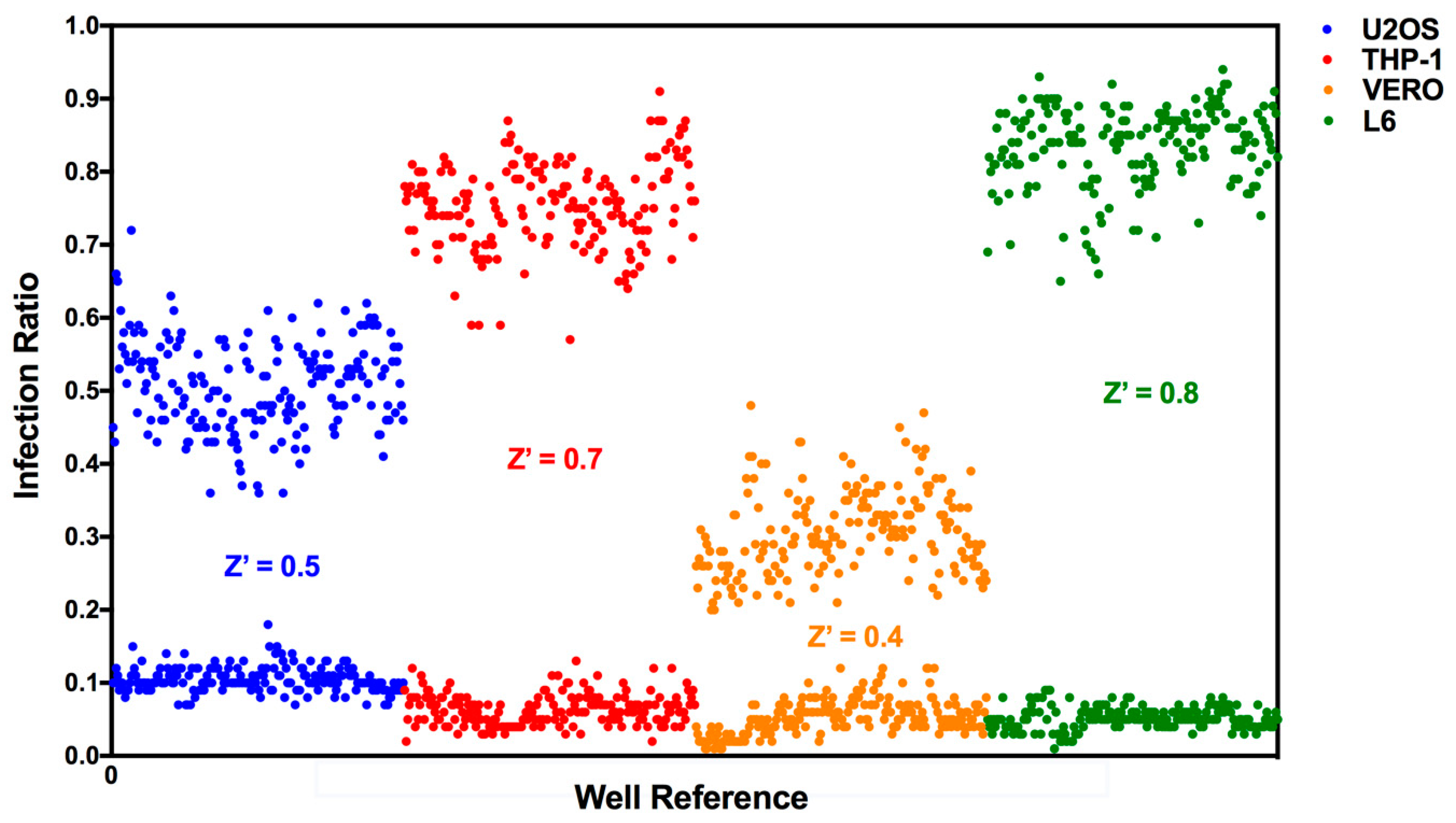
3.1. T. cruzi High-Content Assay Development for Different Host Cell Lines
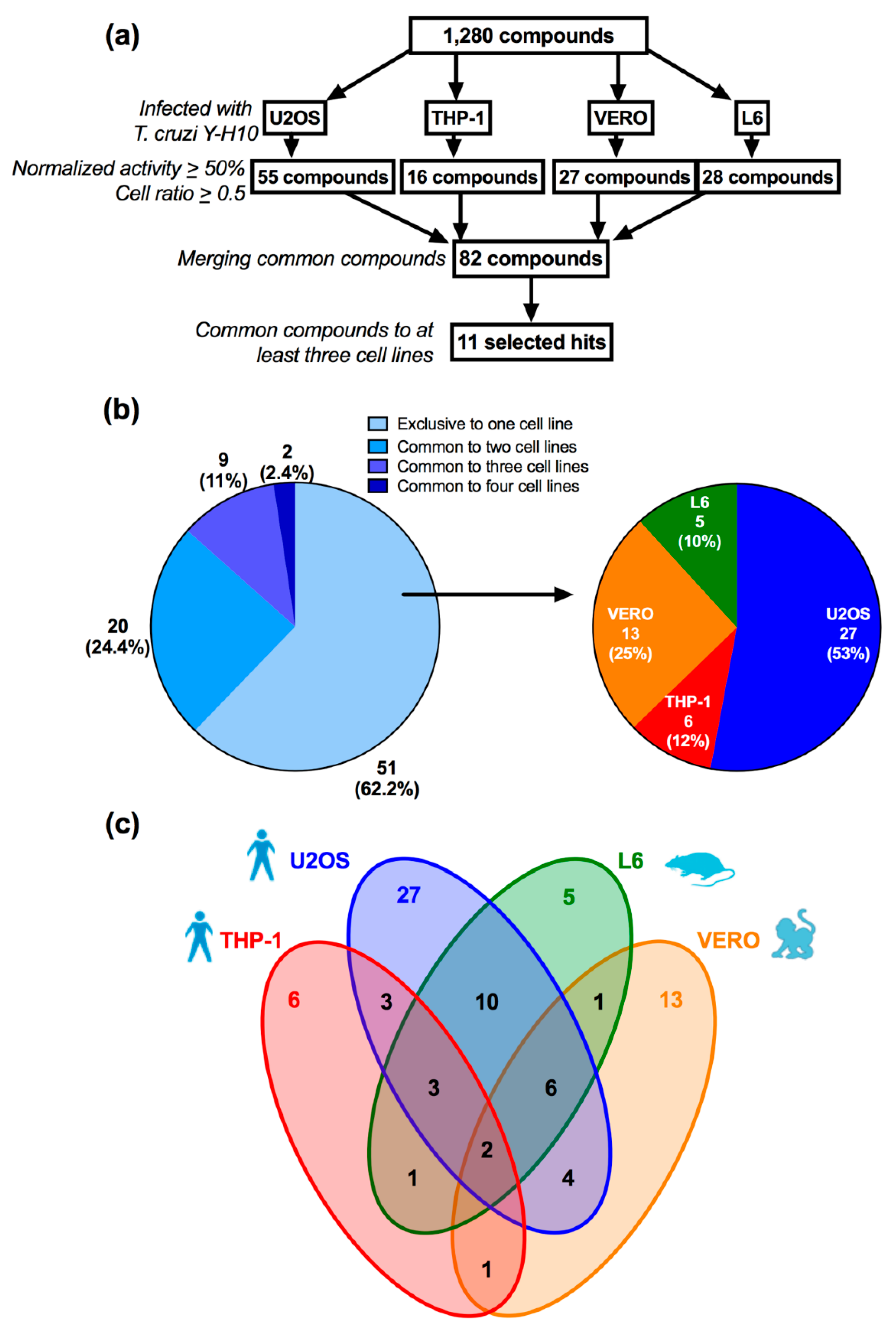
3.2. Assay Performance in Compound Library Screening
3.3. Hit-Compound Activity Confirmation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization: Chagas Disease (American trypanosomiasis) Fact Sheet. Available online: https://www.who.int/en/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 12 December 2018).
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Martins-Melo, F.R.; Alencar, C.H.; Ramos, A.N.; Heukelbach, J. Epidemiology of mortality related to Chagas’ disease in Brazil, 1999–2007. PLoS Negl. Trop. Dis. 2012, 6, e1508. [Google Scholar] [CrossRef] [PubMed]
- Capuani, L.; Bierrenbach, A.L.; Pereira Alencar, A.; Mendrone, A.; Ferreira, J.E.; Custer, B.P.; Ribeiro, A.L.; Cerdeira Sabino, E. Mortality among blood donors seropositive and seronegative for Chagas disease (1996–2000) in São Paulo, Brazil: A death certificate linkage study. PLoS Negl. Trop. Dis. 2017, 11, e0005542. [Google Scholar] [CrossRef] [PubMed]
- Yun, O.; Lima, M.A.; Ellman, T.; Chambi, W.; Castillo, S.; Flevaud, L.; Roddy, P.; Parreño, F.; Albajar Viñas, P.; Palma, P.P. Feasibility, drug safety and effectiveness of etiological treatment programs for Chagas disease in Honduras, Guatemala and Bolivia: 10-year experience of Médecins Sans Frontières. PLoS Negl. Trop. Dis. 2009, 3, e488. [Google Scholar] [CrossRef] [PubMed]
- Viotti, R.; Vigliano, C.; Lococo, B.; Alvarez, M.G.; Petti, M.; Bertocchi, G.; Armenti, A. Side effects of benznidazole as treatment in chronic Chagas disease: Fears and realities. Expert Rev. Anti. Infect. Ther. 2009, 7, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Viotti, R.; Vigliano, C.; Lococo, B.; Bertocchi, G.; Petti, M.; Alvarez, M.G.; Postan, M.; Armenti, A. Long-term cardiac outcomes of treating chronic Chagas disease with benznidazole versus no treatment: A nonrandomized trial. Ann. Intern. Med. 2006, 144, 724–734. [Google Scholar] [CrossRef]
- Pinazo, M.-J.; Espinosa, G.; Cortes-Lletget, C.; Posada, E.D.J.; Aldasoro, E.; Oliveira, I.; Muñoz, J.; Gállego, M.; Gascon, J. Immunosuppression and Chagas disease: A management challenge. PLoS Negl. Trop. Dis. 2013, 7, e1965. [Google Scholar] [CrossRef]
- Velazquez, E.; Meeks, B.; Bonilla, L.; Rassi, A.; Bonilla, R.; Lazdins, J.; Pogue, J.; Sosa-Estani, S.; Rao-Melacini, P.; Britto, C.; et al. Randomized trial of benznidazole for chronic chagas’ cardiomyopathy. N. Engl. J. Med. 2015, 373, 1295–1306. [Google Scholar]
- Molina, I.; Gómez I Prat, J.; Salvador, F.; Treviño, B.; Sulleiro, E.; Serre, N.; Pou, D.; Roure, S.; Cabezos, J.; Valerio, L.; et al. Randomized trial of posaconazole and benznidazole for chronic Chagas’ disease. N. Engl. J. Med. 2014, 370, 1899–1908. [Google Scholar] [CrossRef]
- Torrico, F.; Gascon, J.; Ortiz, L.; Alonso-Vega, C.; Pinazo, M.J.; Schijman, A.; Almeida, I.C.; Alves, F.; Strub-Wourgaft, N.; Ribeiro, I.; et al. Treatment of adult chronic indeterminate Chagas disease with benznidazole and three E1224 dosing regimens: A proof-of-concept, randomised, placebo-controlled trial. Lancet Infect. Dis. 2018, 18, 419–430. [Google Scholar] [CrossRef]
- Field, M.C.; Fairlamb, A.H.; Horn, D.; Gray, D.W.; Ferguson, M.A.J.; Torrie, L.S.; Read, K.D.; Gilbert, I.H.; Wyllie, S.; De Rycker, M.; et al. Anti-trypanosomatid drug discovery: An ongoing challenge and a continuing need. Nat. Rev. Microbiol. 2017, 15, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Valdéz, F.J.; Padilla, A.; Wang, W.; Orr, D.; Tarleton, R.L. Spontaneous dormancy protects Trypanosoma cruzi during extended drug exposure. Elife 2018, 7, e34039. [Google Scholar] [CrossRef] [PubMed]
- MacLean, L.M.; Thomas, J.; Lewis, M.D.; Cotillo, I.; Gray, D.W.; De Rycker, M. Development of Trypanosoma cruzi in vitro assays to identify compounds suitable for progression in Chagas’ disease drug discovery. PLoS Negl. Trop. Dis. 2018, 12, e0006612. [Google Scholar] [CrossRef] [PubMed]
- Moraes, C.B.; Franco, C.H. Novel drug discovery for Chagas disease. Expert Opin. Drug Discov. 2016, 11, 447–455. [Google Scholar] [CrossRef]
- Chatelain, E.; Ioset, J.-R. Phenotypic screening approaches for Chagas disease drug discovery. Expert Opin. Drug Discov. 2018, 13, 141–153. [Google Scholar] [CrossRef]
- Duran-Rehbein, G.A.; Vargas-Zambrano, J.C.; Cuéllar, A.; Puerta, C.J.; Gonzalez, J.M. Mammalian cellular culture models of Trypanosoma cruzi infection: A review of the published literature. Parasite 2014, 21, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Engel, J.C.; Ang, K.K.H.; Chen, S.; Arkin, M.R.; McKerrow, J.H.; Doyle, P.S. Image-based high-throughput drug screening targeting the intracellular stage of Trypanosoma cruzi, the agent of Chagas’ disease. Antimicrob. Agents Chemother. 2010, 54, 3326–3334. [Google Scholar] [CrossRef]
- Nohara, L.L.; Lema, C.; Bader, J.O.; Aguilera, R.J.; Almeida, I.C. High-content imaging for automated determination of host-cell infection rate by the intracellular parasite Trypanosoma cruzi. Parasitol. Int. 2010, 59, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Sykes, M.L.; Avery, V.M. Development and application of a sensitive, phenotypic, high-throughput image-based assay to identify compound activity against Trypanosoma cruzi amastigotes. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 215–228. [Google Scholar] [CrossRef]
- Neitz, R.J.; Chen, S.; Supek, F.; Yeh, V.; Kellar, D.; Gut, J.; Bryant, C.; Gallardo-Godoy, A.; Molteni, V.; Roach, S.L.; et al. Lead identification to clinical candidate selection: drugs for Chagas disease. J. Biomol. Screen. 2014, 20, 101–111. [Google Scholar] [CrossRef]
- Moon, S.; Siqueira-Neto, J.L.; Moraes, C.B.; Yang, G.; Kang, M.; Freitas, L.H., Jr.; Hansen, M.A.E. An image-based algorithm for precise and accurate high throughput assessment of drug activity against the human parasite Trypanosoma cruzi. PLoS ONE 2014, 9, e87188. [Google Scholar] [CrossRef]
- Alonso-Padilla, J.; Cotillo, I.; Presa, J.L.; Cantizani, J.; Peña, I.; Bardera, A.I.; Martín, J.J.; Rodriguez, A. Automated high-content assay for compounds selectively Toxic to Trypanosoma cruzi in a myoblastic cell line. PLoS Negl. Trop. Dis. 2015, 9, e0003493. [Google Scholar] [CrossRef] [PubMed]
- De Rycker, M.; Thomas, J.; Riley, J.; Brough, S.J.; Miles, T.J.; Gray, D.W. Identification of Trypanocidal activity for known clinical compounds using a new Trypanosoma cruzi hit-discovery screening cascade. PLoS Negl. Trop. Dis. 2016, 10, e0004584. [Google Scholar] [CrossRef]
- Filardi, L.S.; Brener, Z. Susceptibility and natural resistance of Trypanosoma cruzi strains to drugs used clinically in Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 755–759. [Google Scholar] [CrossRef]
- Neal, R.A.; van Bueren, J. Comparative studies of drug susceptibility of five strains of Trypanosoma cruzi in vivo and in vitro. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 709–714. [Google Scholar] [CrossRef]
- Camandaroba, E.L.P.; Reis, E.A.G.; Gonçalves, M.S.; Reis, M.G.; Andrade, S.G. Trypanosoma cruzi: Susceptibility to chemotherapy with benznidazole of clones isolated from the highly resistant Colombian strain. Rev. Soc. Bras. Med. Trop. 2003, 36, 201–209. [Google Scholar] [CrossRef]
- Moraes, C.B.; Giardini, M.A.; Kim, H.; Franco, C.H.; Araujo, A.M., Jr.; Schenkman, S.; Chatelain, E.; Freitas-Junior, L.H. Nitroheterocyclic compounds are more efficacious than CYP51 inhibitors against Trypanosoma cruzi: Implications for Chagas disease drug discovery and development. Sci. Rep. 2014, 4, 4703. [Google Scholar] [CrossRef]
- Melo, R.C.; Brener, Z. Tissue tropism of different Trypanosoma cruzi strains. J. Parasitol. 1978, 64, 475–482. [Google Scholar] [CrossRef]
- Andrade, L.O.; Machado, C.R.S.; Chiari, E.; Pena, S.D.J.; Macedo, A.M. Differential tissue distribution of diverse clones of Trypanosoma cruzi in infected mice. Mol. Biochem. Parasitol. 1999, 100, 163–172. [Google Scholar] [CrossRef]
- Lewis, M.D.; Fortes Francisco, A.; Taylor, M.C.; Burrell-Saward, H.; McLatchie, A.P.; Miles, M.A.; Kelly, J.M. Bioluminescence imaging of chronic Trypanosoma cruzi infections reveals tissue-specific parasite dynamics and heart disease in the absence of locally persistent infection. Cell. Microbiol. 2014, 16, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Caradonna, K.L.; Engel, J.C.; Jacobi, D.; Lee, C.-H.; Burleigh, B.A. Host metabolism regulates intracellular growth of Trypanosoma cruzi. Cell Host Microbe 2013, 13, 108–117. [Google Scholar] [CrossRef]
- Machado, F.C.; Franco, C.H.; dos Santos Neto, J.V.; Dias-Teixeira, K.L.; Moraes, C.B.; Lopes, U.G.; Aktas, B.H.; Schenkman, S. Identification of di-substituted ureas that prevent growth of trypanosomes through inhibition of translation initiation. Sci. Rep. 2018, 8, 4857. [Google Scholar] [CrossRef] [Green Version]
- Delvecchio, R.; Higa, L.; Pezzuto, P.; Valadão, A.; Garcez, P.; Monteiro, F.; Loiola, E.; Dias, A.; Silva, F.; Aliota, M.; et al. Chloroquine, an endocytosis blocking agent, inhibits zika virus infection in different cell models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef]
- Franco, C.H.; David, C.W.; Tapan, B.; Ho, Y.A.A.; Hai, L.; Miriam, A.G.; Bruno, S.P.; Lucio, H.F.-J.; Eric, C.; Michael, A.M.; et al. Novel structural CYP51 mutation in Trypanosoma cruzi associated with multidrug resistance to CYP51 inhibitors and reduced infectivity. Unpublished work. 2019. [Google Scholar]
- Bosc, D.; Mouray, E.; Cojean, S.; Franco, C.H.; Loiseau, P.M.; Freitas, L.H., Jr.; Moraes, C.B.; Grellier, P.; Dubois, J. Highly improved antiparasitic activity after introduction of an N -benzylimidazole moiety on protein farnesyltransferase inhibitors. Eur. J. Med. Chem. 2016, 109, 173–186. [Google Scholar] [CrossRef]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef]
- Silva, F.T.; Franco, C.H.; Favaro, D.C.; Freitas, L.H., Jr.; Moraes, C.B.; Ferreira, E.I. Design, synthesis and antitrypanosomal activity of some nitrofurazone 1,2,4-triazolic bioisosteric analogues. Eur. J. Med. Chem. 2016, 121, 553–560. [Google Scholar] [CrossRef]
- Balfour, M.N.; Franco, C.H.; Moraes, C.B.; Freitas, L.H., Jr.; Stefani, H.A. Synthesis and trypanocidal activity of a library of 4-substituted 2-(1H-pyrrolo[3,2-c]pyridin-2-yl)propan-2-ols. Eur. J. Med. Chem. 2017, 128, 202–212. [Google Scholar] [CrossRef]
- Auwerx, J. The human leukemia cell line, THP-1: A multifacetted model for the study of monocyte-macrophage differentiation. Experientia 1991, 47, 22–31. [Google Scholar] [CrossRef]
- Nahapetian, A.T.; Thomas, J.N.; Thilly, W.G. Optimization of environment for high density Vero cell culture: Effect of dissolved oxygen and nutrient supply on cell growth and changes in metabolites. J. Cell Sci. 1986, 81, 65–103. [Google Scholar]
- Jorge, T.C.A.; Barbosa, H.S.; Moreira, A.L.; De Souza, W.; Meirelles, M.N.L. The interaction of myotropic and macrophagotropic strains of Trypanosoma cruzi with myoblasts and fibers of skeletal muscle. Z. Parasitenkd. 1986, 72, 577–584. [Google Scholar] [CrossRef]
- Andrade, L.O.; Galvão, L.M.C.C.; Nazareth, M.D.; Meirelles, S.L.; Chiari, E.; Pena, S.D.J.J.; Macedo, A.M.; de Nazareth SL Meirelles, M.; Chiari, E.; Pena, S.D.J.J.; et al. Differential tissue tropism of Trypanosoma cruzi strains: An in vitro study. Mem. Inst. Oswaldo Cruz 2010, 105, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.A.; Carpenter, A. Advanced assay development guidelines for image-based high content screening and analysis. In Assay Guidance Manual from Imaging Platform; MIT and Harvard: Cambridge, MA, USA, 2004; pp. 1–33. [Google Scholar]
- Luna, K.P.; Hernández, I.P.; Rueda, C.M.; Zorro, M.M.; Croft, S.L.; Escobar, P. In vitro susceptibility of Trypanosoma cruzi strains from Santander, Colombia, to hexadecylphosphocholine (miltefosine), nifurtimox and benznidazole. Biomedica 2009, 29, 448–455. [Google Scholar] [PubMed]
- Seifert, K.; Escobar, P.; Croft, S.L. In vitro activity of anti-leishmanial drugs against Leishmania donovani is host cell dependent. J. Antimicrob. Chemother. 2010, 65, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Genovesio, A.; Giardini, M.A.; Kwon, Y.J.; de Macedo Dossin, F.; Choi, S.Y.; Kim, N.Y.; Kim, H.C.; Jung, S.Y.; Schenkman, S.; Almeida, I.C.; et al. Visual genome-wide RNAi screening to identify human host factors required for Trypanosoma cruzi infection. PLoS ONE 2011, 6, e19733. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Dobson, P.D.; Bilsland, E.; Oliver, S.G. The promiscuous binding of pharmaceutical drugs and their transporter-mediated uptake into cells: What we (need to) know and how we can do so. Drug Discov. Today 2013, 18, 218–239. [Google Scholar] [CrossRef]
- Kunze, D.L.; Rampe, D. Characterization of the effects of a new Ca2+ channel activator, FPL 64176, in GH3 cells. Mol. Pharmacol. 1992, 42, 666–670. [Google Scholar] [PubMed]
- Benaim, G.; Garcia, C.R.S. Targeting calcium homeostasis as the therapy of Chagas’ disease and leishmaniasis—A review. Trop. Biomed. 2011, 28, 471–481. [Google Scholar]
- Benaim, G.; Sanders, J.M.; Garcia-Marchán, Y.; Colina, C.; Lira, R.; Caldera, A.R.; Payares, G.; Sanoja, C.; Burgos, J.M.; Leon-Rossell, A.; et al. Amiodarone has intrinsic anti-Trypanosoma cruzi activity and acts synergistically with posaconazole. J. Med. Chem. 2006, 49, 892–899. [Google Scholar] [CrossRef]
- Reimão, J.Q.; Mesquita, J.T.; Ferreira, D.D.; Tempone, A.G. Investigation of calcium channel blockers as antiprotozoal agents and their interference in the metabolism of Leishmania (L.) infantum. Evid. Based Complement. Altern. Med. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | MOI * | Infection Ratio (%) | Number of Host Cells | Intracellular Parasites/Infected Cell | Number of Intracellular Parasites |
|---|---|---|---|---|---|
| U2OS | 20 | 52 ± 0.1 | 1241 ± 124 | 11 ± 0.4 | 6493 ± 48 |
| VERO | 20 | 28 ± 4 | 2952 ± 101 | 5 ± 1 | 3943 ± 402 |
| L6 | 20 | 77 ± 7 | 1926 ± 846 | 7 ± 1 | 10,708 ±3846 |
| THP-1 | 4 | 73 ± 4 | 320 ± 41 | 15 ± 1 | 3644 ± 893 |
| Compound | T. cruzi Strains | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Y-H10 | Sylvio X10/1 | CL Brener | |||||||
| EC50 (µM) | S.I. | MA (%) | EC50 (µM) | S.I. | MA (%) | EC50 (µM) | S.I. | MA (%) | |
| Benznidazole | 23.3 ± 6.8 | >8.6 | 85.9 | 3.4 ± 1.5 | >70 | 98.4 | 4.0 ± 0.3 | >208 | 107.6 |
| FPL64176 | 2.3 ± 0.2 | 57.7 | 99.8 | 3.0 ± 1.6 | 35.4 | 97.6 | 2.2 ± 0.6 | >90 | 111.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franco, C.H.; Alcântara, L.M.; Chatelain, E.; Freitas-Junior, L.; Moraes, C.B. Drug Discovery for Chagas Disease: Impact of Different Host Cell Lines on Assay Performance and Hit Compound Selection. Trop. Med. Infect. Dis. 2019, 4, 82. https://doi.org/10.3390/tropicalmed4020082
Franco CH, Alcântara LM, Chatelain E, Freitas-Junior L, Moraes CB. Drug Discovery for Chagas Disease: Impact of Different Host Cell Lines on Assay Performance and Hit Compound Selection. Tropical Medicine and Infectious Disease. 2019; 4(2):82. https://doi.org/10.3390/tropicalmed4020082
Chicago/Turabian StyleFranco, Caio Haddad, Laura Maria Alcântara, Eric Chatelain, Lucio Freitas-Junior, and Carolina Borsoi Moraes. 2019. "Drug Discovery for Chagas Disease: Impact of Different Host Cell Lines on Assay Performance and Hit Compound Selection" Tropical Medicine and Infectious Disease 4, no. 2: 82. https://doi.org/10.3390/tropicalmed4020082
APA StyleFranco, C. H., Alcântara, L. M., Chatelain, E., Freitas-Junior, L., & Moraes, C. B. (2019). Drug Discovery for Chagas Disease: Impact of Different Host Cell Lines on Assay Performance and Hit Compound Selection. Tropical Medicine and Infectious Disease, 4(2), 82. https://doi.org/10.3390/tropicalmed4020082