Cholera and Pancreatic Cholera: Is VIP the Common Pathophysiologic Factor?

 ,
, 
Abstract
:Key Point’s/Summary
1. Introduction
2. Background
3. Study Design and Settings
4. Study Population and Site
5. Stool and Blood Samples for VIP Assay
6. Statistics
7. Results
8. Discussion
9. Limitations of this Study
10. Conclusions
11. Authors’ Translational Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Bloom, S.R.; Polak, J.M.; Pearse, A.G. Vasoactive intestinal peptide and watery-diarrhoea syndrome. Lancet 1973, 2, 14–16. [Google Scholar] [CrossRef]
- Said, S.I. Vasoactive intestinal polypeptide (VIP) as a mediator of the watery diarrhea syndrome. World J. Surg. 1979, 3, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Editorial. VIP and diarrhea. Lancet 1984, 28, 202. [Google Scholar]
- Bloom, S.R.; Nalin, D.R.; Mitchell, S.J.; Bryant, M.G. High Levels of Vip in Cholera Stool Water. Gut 1976, 17, 817. [Google Scholar]
- Chen, L.C.; Rohde, J.E.; Sharp, G.W. Intestinal adenyl-cyclase activity in human cholera. Lancet 1971, 1, 939–941. [Google Scholar] [CrossRef]
- Field, M. Intestinal Secretion—Effect of Cyclic Amp and Its Role in Cholera. N. Engl. J. Med. 1971, 284, 1137–1144. [Google Scholar]
- Eklund, S.; Cassuto, J.; Jodal, M.; Lundgren, O. The involvement of the enteric nervous system in the intestinal secretion evoked by cyclic adenosine 3’5’-monophosphate. Acta Physiol. Scand. 1984, 120, 311–316. [Google Scholar] [CrossRef]
- Schwartz, C.J.; Kimberg, D.V.; Sheerin, H.E.; Field, M.; Said, S.I. Vasoactive intestinal peptide stimulation of adenylate cyclase and active electrolyte secretion in intestinal mucosa. J. Clin. Investig. 1974, 54, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Eklund, S.; Brunsson, I.; Jodal, M.; Lundgren, O. Changes in cyclic 3’5’-adenosine monophosphate tissue concentration and net fluid transport in the cat’s small intestine elicited by cholera toxin, arachidonic acid, vasoactive intestinal polypeptide and 5-hydroxytryptamine. Acta Physiol. Scand. 1987, 129, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Loeschke, K.; Farack, U.M.; Gerzer, R.; Keravis, T. Evidence That the Turnover Rather Than the Concentration of Camp Determines Cholera-Toxin Induced Fluid Secretion in Rat Intestine. Z. Fur Gastroenterol. 1987, 25, 388. [Google Scholar]
- Sjoqvist, A.; Fahrenkrug, J.; Jodal, M.; Lundgren, O. The effect of splanchnic nerve stimulation and neuropeptide Y on cholera secretion and release of vasoactive intestinal polypeptide in the feline small intestine. Acta Physiol. Scand. 1988, 133, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Bennett, V.; Mong, L.; Cuatrecasas, P. Mechanism of activation of adenylate cyclase by Vibrio cholerae enterotoxin: Relations to the mode of activation by hormones. J. Membr. Biol. 1975, 24, 107–129. [Google Scholar] [CrossRef]
- Brunsson, I.; Eklund, S.; Fahrenkrug, J.; Jodal, M.; Lundgren, O.; Sjoqvist, A. Effects of substance P on intestinal secretion, blood flow, motility and release of vasoactive intestinal polypeptide in vivo in the rat and cat. J. Physiol. 1995, 483, 727–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassuto, J.; Jodal, M.; Tuttle, R.; Lundgren, O. 5-hydroxytryptamine and cholera secretion. Physiological and pharmacological studies in cats and rats. Scand. J. Gastroenterol. 1982, 17, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Cassuto, J.; Larsson, P.A.; Jodal, M.; Lidberg, P.; Ahlman, H.; Dahlstrom, A.; Lundgren, O. 5-Hydroxytryptamine and cholera secretion: A histochemical and physiological study in cats. Gut 1983, 24, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Mourad, F.H.; O’Donnell, L.J.D.; Dias, J.A.; Ogutu, E.; Andre, E.A.; Turvill, J.L.; Farthing, M.J.G. Role of 5-hydroxytriptamine type 3 receptors in rat intestinal fluid and electrolyte secretion induced by cholera and Escherichia coli enterotoxins. Gut 1995, 37, 340–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, A.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T. Enhancement of aquaporin-3 by vasoactive intestinal polypeptide in a human colonic epithelial cell line. J. Gastroenterol. Hepatol. 2003, 18, 203–221. [Google Scholar] [CrossRef]
- Carpenter, C.C.; Greenough, W.B. Response of the canine duodenum to intraluminal challenge with cholera exotoxin. J. Clin. Investig. 1968, 47, 2600–2607. [Google Scholar] [CrossRef]
- Perez-Rector, D.; Jana, V.; Pavez, L.; Navrante, P.; Garcia, K. Acceessory toxins of vibrio pathogens and their role in epithelial disruption during infection. Front. Microbiol. 2018, 9, 2248. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M.; Nullens, S.; Nelsen, T. Enteroendocrine and neuronal mechanisms in pathophysiology of acute infectious diarrhea. Dig. Dis. Sci. 2012, 57, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Barua, D.; Burrows, W. Cholera; Saunders: Philadelphia, PA, USA, 1974; pp. 18–23. [Google Scholar]
- Iwasaki, M.; Akiba, Y.; Kaunitz, J.D. Recent Advances in Vasoactive Intestinal Peptide Physiology and Pathophysiology: Focus on the Gastrointestinal System; F1000Research: London, UK, 2019. [Google Scholar]
- Davis, G.R.; Santa Ana, C.A.; Morawski, S.G.; Fordtran, J.S. Effect of vasoactive intestinal polypeptide on active and passive transport in the human jejunum. J. Clin. Investig. 1981, 67, 1687–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreis, G.J.; Fordtran, J.S.; Fahrenkrug, J.; Schaffalitzky, D.E.; Muckadell, O.B.; Fischer, J.E.; Humphrey, C.S.; Said, S.I.; Walsh, J.H.; Shulkes, A.A. O’dorisio TMA, Effect of VIP infusion in water and ion transport in the human jejunum. Gastroenteritis 1980, 78, 722–727. [Google Scholar]
- Kane, M.G.; O’Dorisio, T.M.; Krejs, G.J. Production of secretory diarrhea by intravenous infusion of vasoactive intestinal polypeptide. N. Engl. J. Med. 1983, 309, 1482–1485. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M.; Hirsch, J.R.; Loo, D.D.; Zampighi, G.A. Regulation of Na+/glucose cotransporters. J. Exp. Biol. 1997, 200, 287–293. [Google Scholar] [PubMed]
- Reymann, A.; Braun, W.; Woermann, C. Proabsorptive properties of forskolin: Disposition of glycine, leucine and lysine in rat jejunum. Naunyn Schm. Arch. Pharmacol. 1986, 334, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Nalin, D.R.; Cash, R.A.; Rahman, M.; Yunus, M. Effect of glycine and glucose on sodium and water adsorption in patients with cholera. Gut 1970, 11, 768–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robichaux, W.G.; Xiaodong, C. Intracellular cAMP sensor EPAC: Physiology, pathophysiology and therapeutics development. Physiol. Rev. 2018, 98, 919–1053. [Google Scholar] [CrossRef] [PubMed]
- Sakio, H.; Matsuzaki, Y.; Said, S.I. Release of Vasoactive Intestinal Polypeptide during Hemorrhagic-Shock. Fed. Proc. 1979, 38, 1114. [Google Scholar]
- Hirschhorn, N.; Kinzie, J.L.; Sachar, D.B.; Northrup, R.S.; Taylor, J.O.; Ahmad, S.Z.; Phillips, R.A. Decrease in net stool output in cholera during intestinal perfusion with glucose-containing solutions. N. Engl. J. Med. 1968, 279, 176–181. [Google Scholar] [CrossRef]
- Alam, N.H.; Ashraf, H. Treatment of infectious diarrhea in children. Paediatr. Drugs 2003, 5, 151–165. [Google Scholar] [CrossRef]
- Cash, R.A.; Toha, K.M.; Nalin, D.R.; Huq, Z.; Phillips, R.A. Acetate in the correction of acidosis secondary to diarrhoea. Lancet 1969, 2, 302–303. [Google Scholar] [CrossRef]
- World Health Organization. Programme for the Control of Diarrhoeal Diseases: A Manual for the Treatment of Diarrhoea for Use by Physicians and Other Health Workers; Geneva World Health Organization: Geneva, Switzerland, 1990. [Google Scholar]
- World Health Organization. The Treatment of Diarrhoea: A Manual for Physicians and Other Senior Health Workers; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Mitchell, S.J.; Bloom, S.R. Measurement of fasting and postprandial plasma VIP in man. Gut 1978, 19, 1043–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeger, S.L.; Liang, K.Y.; Albert, P.S. Models for longitudinal data: A generalized estimating equation approach. Biometrics 1988, 44, 1049–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besterman, H.S.; Christofides, N.D.; Welsby, P.D.; Adrian, T.E.; Sarson, D.L.; Bloom, S.R. Gut hormones in acute diarrhoea. Gut 1983, 24, 665–671. [Google Scholar] [CrossRef]
- Manfredi, R.; Vezzadini, P.; Costigliola, P.; Ricchi, E.; Fanti, M.P.; Chiodo, F. Elevated plasma levels of vasoactive intestinal peptide in AIDS patients with refractory idiopathic diarrhoea. Effects of treatment with octreotide. AIDS 1993, 7, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Molla, A.M.; Sarker, S.A.; Hossain, M.; Molla, A.; Greenough, W.B. Rice-powder electrolyte solution as oral-therapy in diarrhoea due to Vibrio cholerae and Escherichia coli. Lancet 1982, 1, 1317–1319. [Google Scholar] [CrossRef]
- Amankwah, E.N.; Adu, E.; Barimah, V.M.J.; Van Twisk, C. Amino acid profiles of some varieties of rice, soybean and groundnut grown in Ghana. J. Food Process. Technol. 2015, 6, 420–423. [Google Scholar] [CrossRef]
- Kalman, D.S. Amino Acid Composition of an Organic Brown Rice Protein Concentrate and Isolate Compared to Soy and Whey Concentrates and Isolates. Foods 2014, 3, 394–402. [Google Scholar] [CrossRef]
- Vesikari, T.; Isolauri, E. Glycine supplemented oral rehydration solutions for diarrhoea. Arch. Dis. Child. 1986, 61, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Nalin, D.R.; Ally, K.; Hare, R.; Hare, K. Effect of cholera toxin on jejunal osmoregulation of mannitol solutions in dogs. J. Infect. Dis. 1972, 125, 528–532. [Google Scholar] [PubMed]
- Cassuto, J.; Fahrenkrug, J.; Jodal, M.; Tuttle, R.; Lundgren, O. Release of vasoactive intestinal polypeptide from the cat small intestine exposed to cholera toxin. Gut 1981, 22, 958–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, M.M.; Nalin, D.R.; Craig, J.P.; Hoover, D.; Bergquist, E.J.; Waterman, D.; Holley, H.P.; Hornick, R.B.; Pierce, N.P.; Libonati, J.P. Immunity of cholera in man: Relative role of antibacterial versus antitoxic immunity. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 3–9. [Google Scholar] [CrossRef]
- Cassuto, J.; Jodal, M.; Tuttle, R.; Lundgren, O. On the role of intramural nerves in the pathogenesis of cholera toxin-induced intestinal secretion. Scand. J. Gastroenterol. 1981, 16, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Eklund, S.; Sjoqvist, A.; Fahrenkrug, J.; Jodal, M.; Lundgren, O. Somatostatin and methionine-enkephalin inhibit cholera toxin-induced jejunal net fluid secretion and release of vasoactive intestinal polypeptide in the cat in vivo. Acta Physiol. Scand. 1988, 133, 551–557. [Google Scholar] [CrossRef]
- Molla, A.M.; Gyr, K.; Bardhan, P.K.; Molla, A. Effect of Intravenous Somatostatin on Stool Output in Diarrhea Due to Vibrio-Cholerae. Gastroenterology 1984, 87, 845–847. [Google Scholar] [CrossRef]
- Banks, M.R.; Farthing, M.J.; Robberecht, P.; Burleigh, D.E. Antisecretory actions of a novel vasoactive intestinal polypeptide (VIP) antagonist in human and rat small intestine. Br. J. Pharmacol. 2005, 144, 994–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourad, F.H.; Nassar, C.F. Effect of vasoactive intestinal polypeptide (VIP) antagonism on rat jejunal fluid and electrolyte secretion induced by cholera and Escherichia coli enterotoxins. Gut 2000, 47, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Emery, P.T.; Higgs, N.B.; Warhurst, A.C.; Carlson, G.L.; Warhurst, G. Anti-secretory properties of non-peptide somatostatin receptor agonists in isolated rat colon: Luminal activity and possible interaction with P-glycoprotein. Br. J. Pharmacol. 2002, 135, 1443–1448. [Google Scholar] [CrossRef] [Green Version]


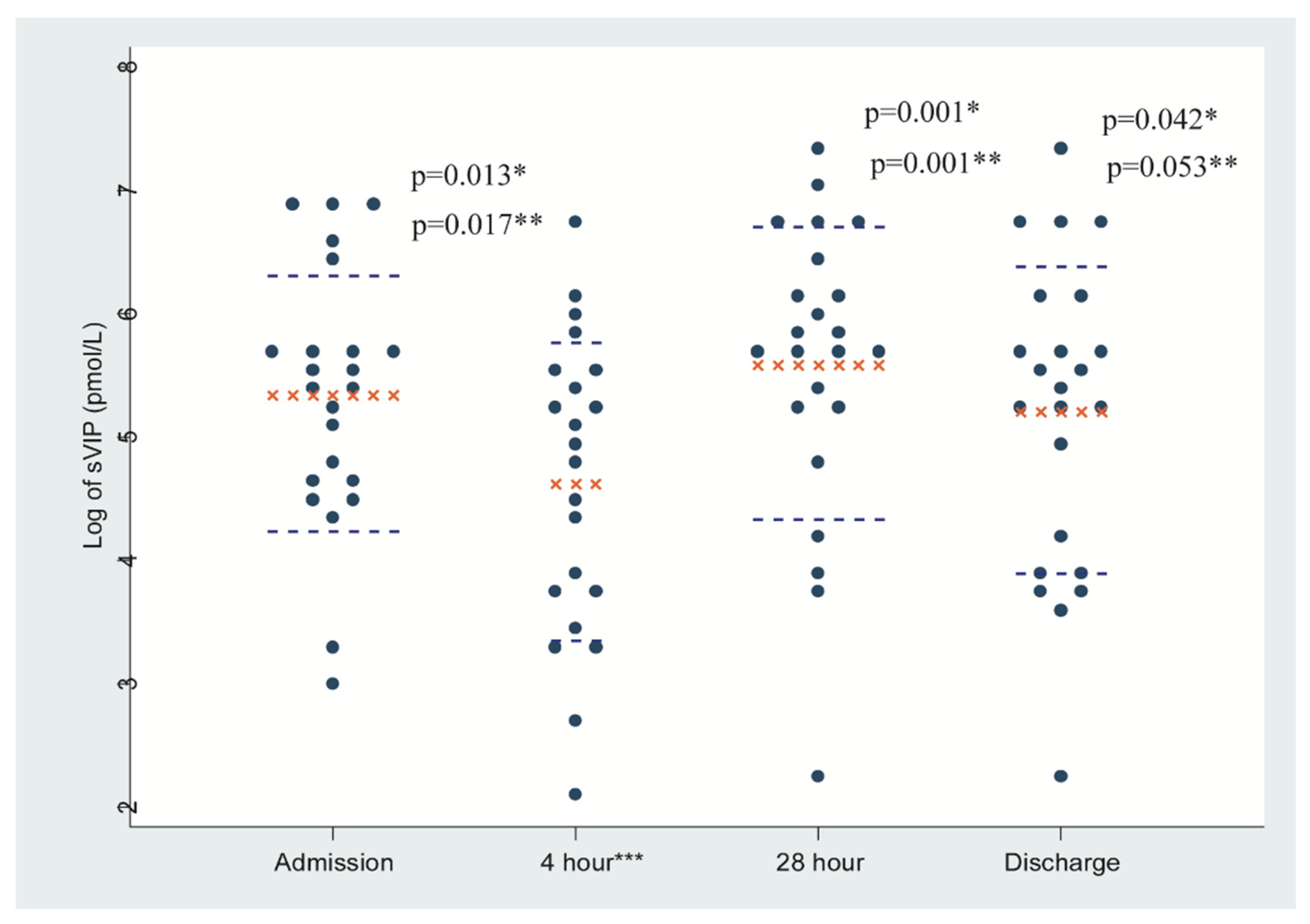{kind=link}
| Characteristics | Total (n = 23) |
|---|---|
| Age (in years) | 33 ± 10 |
| Male (n, %) | 20/23 (87) |
| Diarrhea duration (h) | 41 ± 19 |
| Emesis duration (h) | 2 (1,3) |
| Abdominal pain (n, %) | 4/23 (17) |
| Total stool volume (mL/kg) | 141 (109, 191) |
| Urine output (mL/h) | 136 ± 67 |
| Stool output in 1st 24 h (mL/day) | 8000 (5500, 10,500) |
| Total intravenous fluid (mL/kg/h) | 11 (8.4, 12.75) |
| Duration of hospital stay (h) | 63 ± 23 |
| Admission blood glucose level (mmol/L) | 8 (6,10) (4.2–7.8) |
| Admission plasma sp.gr. | 1.0480 ± 0.006 (1.0232–1.0279) |
| Admission serum.Na+ (mmol/L) | 133.28 ± 2.30 (135–146) |
| Admission serum. K+ (mmol/L) | 5.16 ± 1.15 (3.5–5.3) |
| Admission serum. Cl− (mmol/L) | 102.26 ±3.06 (97–106) |
| Admission serum TCO2 (mmol/L) | 18.14 ± 4.63 (23–30) |
| Anion gap (mmol/L) | 17.95 ±3.82 (7–21) |
| BUN (mmol/L) | 14.80 ±4.80 (5–20) |
| Urine Sp. gr. at 4 h * | 1.04 ± 0.005 (1.005–1.030) |
| Serum Na+ (mmol/L) at 4 h * | 135 ± 2.76 |
| TCO2 (mmol/L) at 4 h * | 22.5 ± 3.57 |
| Blood glucose level (mmol/L) at 4 h * | 5.4 (4.88, 7.39) |
| Mean stool VIP (pmol/L) | 307 (168,410) |
| Peak stool VIP (pmol/L) | 591 (283,855) |
| Vibrio cholerae 01, El Tor | 22/23 (96) |
| Vibrio cholerae 0139 | 1/23 (4) |
| Vibrio cholerae 01, El Tor Ogawa | 17/22 (74) |
| Vibrio cholerae 01, El Tor Inaba | 5/22 (22) |
| VIP Measurement Time Points | Stool VIP (pmol/L) | Plasma VIP (pmol/L) | ||
|---|---|---|---|---|
| Geometric Mean * | 95% CI | Geometric Mean * | 95% CI | |
| Admission (0 h) | 207.67 | 132.74, 324.90 | 8.34 | 5.53, 12.59 |
| At 4 h (after complete rehydration) | 101.55 | 59.40, 173.60 | 3.84 | 2.59, 5.67 |
| At 28 h | 265.97 | 159.17, 444.41 | 2.67 | 1.82, 3.92 |
| At discharge | 181.40 | 105.17, 310.63 | 3.31 | 2.26, 4.83 |
| Characteristics | Unadjusted exp (Coefficient) * with 95% CI | p-Value | Adjusted exp (Coefficient) * with 95% CI | p-Value | |
|---|---|---|---|---|---|
| Duration of hospital stay (h) | 1.01 (0.992, 1.022) | 0.161 | 1.01 (1.001, 1.023) | 0.026 | |
| Total stool volume (mL/kg) | 1.00 (0.993, 1.000) | 0.087 | 1.01 (1.002, 1.025) | 0.023 | |
| Stool output in first 24 h (mL) | 1.00 (0.999, 1.000) | 0.075 | 1.00 (0.999, 0.999) | 0.013 | |
| Total IVF (mL/kg/h) | 0.93 (0.864, 1.007) | 0.077 | 0.94 (0.875, 1.021) | 0.157 | |
| Stool VIP (pmol/L) measurement time points | 4 h | ref | ref | ||
| Admission | 2.01 (1.174, 3.762) | 0.013 | 2.03 (1.136, 3.640) | 0.017 | |
| 28 h | 2.69 (1.504, 4.818) | 0.001 | 2.60 (1.455, 4.662) | 0.001 | |
| Discharge | 1.83 (1.026, 3.287) | 0.042 | 1.77 (0.992, 3.179) | 0.053 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afroze, F.; Bloom, S.; Bech, P.; Ahmed, T.; Sarker, S.A.; Clemens, J.D.; Islam, F.; Nalin, D. Cholera and Pancreatic Cholera: Is VIP the Common Pathophysiologic Factor? Trop. Med. Infect. Dis. 2020, 5, 111. https://doi.org/10.3390/tropicalmed5030111
Afroze F, Bloom S, Bech P, Ahmed T, Sarker SA, Clemens JD, Islam F, Nalin D. Cholera and Pancreatic Cholera: Is VIP the Common Pathophysiologic Factor? Tropical Medicine and Infectious Disease. 2020; 5(3):111. https://doi.org/10.3390/tropicalmed5030111
Chicago/Turabian StyleAfroze, Farzana, Steven Bloom, Paul Bech, Tahmeed Ahmed, Shafiqul Alam Sarker, John D. Clemens, Farhana Islam, and David Nalin. 2020. "Cholera and Pancreatic Cholera: Is VIP the Common Pathophysiologic Factor?" Tropical Medicine and Infectious Disease 5, no. 3: 111. https://doi.org/10.3390/tropicalmed5030111
APA StyleAfroze, F., Bloom, S., Bech, P., Ahmed, T., Sarker, S. A., Clemens, J. D., Islam, F., & Nalin, D. (2020). Cholera and Pancreatic Cholera: Is VIP the Common Pathophysiologic Factor? Tropical Medicine and Infectious Disease, 5(3), 111. https://doi.org/10.3390/tropicalmed5030111