
Screening of Mosquitoes for West Nile Virus and Usutu Virus in Croatia, 2015–2020

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Collection and Identification
2.2. Virological Testing
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, L.A.; Flores, F.S.; Quaglia, A.; Contigiani, M.S. Intertwined arbovirus transmission activity: Reassessing the transmission cycle paradigm. Front. Physiol. 2012, 3, 493. [Google Scholar] [CrossRef] [Green Version]
- Hubálek, Z.; Halouzka, J. West Nile fever—A reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Savic, V.; Petrovic, T.; Toplak, I.; Barbic, L.; Petric, D.; Tabain, I.; Hrnjakovic-Cvjetkovic, I.; Bogdanic, M.; Klobucar, A.; et al. Emerging Trends in the Epidemiology of West Nile and Usutu Virus Infections in Southern Europe. Front. Vet. Sci. 2019, 6, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control (ECDC). Historical Data by Year—West Nile Fever Seasonal Surveillance. 2018. Available online: https://ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/historical (accessed on 27 February 2021).
- Pem-Novosel, I.; Vilibic-Cavlek, T.; Gjenero-Margan, I.; Pandak, N.; Peric, L.; Barbic, L.; Listes, E.; Cvitkovic, A.; Stevanovic, V.; Savini, G. First outbreak of West Nile virus neuroinvasive disease in humans, Croatia, 2012. Vector-Borne Zoonotic Dis. 2014, 14, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Barbic, L.; Stevanovic, V.; Kovac, S.; Maltar, L.; Lohman Jankovic, I.; Vilibic-Cavlek, T.; Madic, J. West Nile virus serosurveillance in horses in Croatia during the 2012 transmission season. Croatian Academy of Sciencies and Art. Med. Sci. 2013, 39, 95–104. [Google Scholar]
- Savić, V.; Barbic, L.; Vilibic-Cavlek, T.; Balenović, M.; Stevanović, V.; Listes, E.; Savini, G. Chickens and horses as sentinels for early warning system in prevention of human West Nile virus infections in Croatia. Slov. Vet. Res. 2016, 53, 292–294. [Google Scholar]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ‘One health’ context 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Barbic, L.; Mrzljak, A.; Brnic, D.; Klobucar, A.; Ilic, M.; Janev-Holcer, N.; Bogdanic, M.; Jemersic, L.; Stevanovic, V.; et al. Emerging and Neglected Viruses of Zoonotic Importance in Croatia. Pathogens 2021, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Balenghien, T.; Vazeille, M.; Grandadam, M.; Schaffner, F.; Zeller, H.; Reiter, P.; Sabatier, P.; Fouque, F.; Bicout, D.J. Vector competence of some French Culex and Aedes mosquitoes for West Nile virus. Vector Borne Zoonotic Dis. 2007, 8, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Engler, O.; Savini, G.; Papa, A.; Figuerola, J.; Groschup, M.; Kampen, H.; Medlock, J.; Vaux, A.; Wilson, A.J.; Werner, D.; et al. European surveillance for West Nile virus in mosquito populations. Int. J. Environ. Res. Public Health 2013, 10, 4869–4895. [Google Scholar] [CrossRef] [Green Version]
- Toma, L.; Cipriani, M.; Goffredo, M.; Romi, R.; Lelli, R. First report on entomological field activities for the surveillance of West Nile disease in Italy. Vet. Ital. 2008, 44, 483–512. [Google Scholar] [PubMed]
- Tamba, M.; Bonilauri, P.; Bellini, R.; Calzolari, M.; Albieri, A.; Sambri, V.; Dottori, M.; Angelini, P. Detection of Usutu Virus Within a West Nile Virus Surveillance Program in Northern Italy. Vector Borne Zoonotic Dis. 2011, 11, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Gaibani, P.; Bellini, R.; Defilippo, F.; Pierro, A.; Albieri, A.; Maioli, G.; Luppi, A.; Rossini, G.; Balzani, A.; et al. Mosquito, bird and human surveillance of West Nile and Usutu viruses in Emilia Romagna Region (Italy) in 2010. PLoS ONE 2012, 7, e38058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calzolari, M.; Chiapponi, C.; Bonilauri, P.; Lelli, D.; Baioni, L.; Barbieri, I.; Lavazza, A.; Pongolini, S.; Dottori, M.; Moreno, A. Co-circulation of two Usutu virus strains in Northern Italy between 2009 and 2014. Infect. Genet. Evol. 2017, 51, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Mavridis, K.; Fotakis, A.E.; Kioulos, I.; Mpellou, S.; Konstantas, S.; Varela, E.; Gewehr, S.; Diamantopoulos, V.; Vontas, J. Detection of West Nile Virus–Lineage 2 in Culex pipiens mosquitoes, associated with disease outbreak in Greece, 2017. Acta Trop. 2018, 182, 64–68. [Google Scholar] [CrossRef]
- Vázquez, A.; Ruiz, S.; Herrero, L.; Moreno, J.; Molero, F.; Magallanes, A.; Sánchez-Seco, M.P.; Figuerola, J.; Tenorio, A. West Nile and Usutu Viruses in Mosquitoes in Spain, 2008–2009. Am. J. Trop. Med. Hyg. 2011, 85, 178–181. [Google Scholar] [CrossRef]
- Čabanová, V.; Šikutová, S.; Straková, P.; Šebesta, O.; Vichová, B.; Zubríkova, D.; Miterpáková, M.; Mendel, J.; Hurníkova, Z.; Hubálek, Z.; et al. Co-Circulation of West Nile and Usutu Flaviviruses in Mosquitoes in Slovakia, 2018. Viruses 2019, 11, 639. [Google Scholar] [CrossRef] [Green Version]
- Petrić, D.; Petrović, T.; Cvjetković, I.H.; Zgomba, M.; Milošević, V.; Lazić, G.; Ćupina, A.I.; Lupulović, D.; Lazić, S.; Dondur, D.; et al. West Nile virus ‘circulation’ in Vojvodina, Serbia: Mosquito, bird, horse and human surveillance. Mol. Cell. Probes 2017, 31, 28–36. [Google Scholar] [CrossRef]
- Gaibani, P.; Rossini, G. An overview of Usutu virus. Microbes Infect. 2017, 19, 382–387. [Google Scholar] [CrossRef]
- Barzon, L. Ongoing and emerging arbovirus threats in Europe. J. Clin. Virol. 2018, 107, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Miesen, P.; Vogels, B.C.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.J.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.C.; Simpson, D.I.; Haddow, A.J.; Knight, E.M. The isolation of West Nile Virus from man and of Usutu virus from the bird-biting mosquito Mansonia aurites (Theobald) in the Entebbe area of Uganda. Ann. Trop. Med. Parasitol. 1964, 58, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef]
- Barbic, L.; Vilibic-Cavlek, T.; Listes, E.; Stevanovic, V.; Gjenero-Margan, I.; Ljubin-Sternak, S.; Pem-Novosel, I.; Listes, I.; Mlinaric-Galinovic, G.; Di Gennaro, A.; et al. Demonstration of Usutu virus antibodies in horses, Croatia. Vector Borne Zoonotic Dis. 2013, 13, 772–774. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Kaic, B.; Barbic, L.; Pem-Novosel, I.; Slavic-Vrzic, V.; Lesnikar, V.; Kurecic-Filipovic, S.; Babic-Erceg, A.; Listes, E.; Stevanovic, V.; et al. First evidence of simultaneous occurrence of West Nile virus and Usutu virus neuroinvasive disease in humans in Croatia during the 2013 outbreak. Infection 2014, 42, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Busquets, N.; Alba, A.; Allepuz, A.; Aranda, C.; Núñez, J.I. Usutu Virus Sequences in Culex pipiens (Diptera:Culicidae), Spain. Emerg. Infect. Dis. 2008, 14, 861–863. [Google Scholar] [CrossRef]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Galletti, G.; Barbieri, I.; Tamba, M.; Lelli, D.; Carra, E.; et al. Evidence of Simultaneous Circulation of West Nile and Usutu Viruses in Mosquitoes Sampled in Emilia-Romagna Region (Italy) in 2009. PLoS ONE 2010, 5, e14324. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Tamba, M.; Tassinari, M.; Gelati, A.; Cordioli, P.; Angelini, P.; et al. Usutu virus persistence and West Nile virus inactivity in the Emilia-Romagna region (Italy) in 2011. PLoS ONE 2013, 8, e63978. [Google Scholar] [CrossRef] [Green Version]
- Pautasso, A.; Radaelli, M.C.; Ballardini, M.; Francese, D.R.; Verna, F.; Modesto, P.; Grattarola, C.; Desiato, R.; Bertolini, S.; Vitale, N.; et al. Detection of West Nile and Usutu Viruses in Italian Free Areas: Entomological Surveillance in Piemonte and Liguria Regions, 2014. Vector-Borne Zoonotic Dis. 2016, 16, 292–294. [Google Scholar] [CrossRef]
- Kemenesi, G.; Buzás, D.; Zana, B.; Kurucz, K.; Krtinić, B.; Kepner, A.; Földes, F.; Jakab, F. First genetic characterization of Usutu virus from Culex pipiens mosquitoes Serbia, 2014. Infect. Genet. Evol. 2018, 63, 58–61. [Google Scholar] [CrossRef]
- Cvjetković, I.H.; Petrović, T.; Petrić, D.; Milošević, U.; Radovanov, J.; Kovačević, G.; Galović, A.J.; Patić, A.; Nikolić, N.; Cvjetković, D.; et al. Usutu Virus: An Emerging Flavivirus In Europe. Arch. Vet. Med. 2017, 10, 25–35. [Google Scholar] [CrossRef]
- Weidinger, P.; Kolodziejek, J.; Bakonyi, T.; Brunthaler, R.; Erdélyi, K.; Weissenböck, H.; Nowotny, N. Different dynamics of Usutu virus infections in austria and Hungary, 2017–2018. Transbound. Emerg. Dis. 2020, 67, 298–307. [Google Scholar] [CrossRef]
- Camp, J.V.; Kolodziejek, J.; Nowotny, N. Targeted surveillance reveals native and invasive mosquito species infected with Usutu virus. Parasites Vectors 2019, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, J.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, Western Europe, 2016. Eurosurveillance 2016, 22, 30452. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, I.; Bakonyi, T.; Šebesta, O.; Mendel, J.; Peško, J.; Betášová, L.; Blažejová, H.; Venclíková, K.; Strakova, P.; Nowotny, N.; et al. Co-circulation of Usutu virus and West Nile virus in a reed bed ecosystem. Parasites Vectors 2015, 8, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiden, M.; Gil, P.; Ziegler, U.; Rakotoarivony, I.; Marie, A.; Francés, B.; L’Ambert, G.; Simonin, Y.; Foulongne, V.; Groschup, M.H.; et al. Emergence of two Usutu virus lineages in Culex pipiens mosquitoes in the Camargue, France, 2015. Infect. Genet. Evol. 2018, 61, 151–154. [Google Scholar] [CrossRef]
- Sieg, M.; Schmidt, V.; Ziegler, U.; Keller, M.; Höper, D.; Heenemann, K.; Rückner, A.; Nieper, H.; Muluneh, A.; Groschup, M.H.; et al. Outbreak and Cocirculation of Three Different Usutu Virus Strains in Eastern Germany. Vector Borne Zoonotic Dis. 2017, 17, 662–664. [Google Scholar] [CrossRef]
- Merdić, E.; Klobučar, A.; Žitko, T.; Sudarić Bogojević, M.; Vrućina, I.; Turić, N.; Vignjević, G. Update checklist of the mosquitoes (Diptera: Culicidae) of Croatia. J. Vector Ecol. 2020, 45, 135–139. [Google Scholar] [CrossRef]
- Klobucar, A.; Benic, N.; Krajcar, D.; Kosanovic-Licina, M.L.; Tesic, V.; Merdic, E.; Vrucina, I.; Savic, V.; Barbic, L.; Stevanovic, V.; et al. An overview of mosquitoes and emerging arboviral infections in the Zagreb area, Croatia. J. Infect. Dev. Ctries. 2016, 10, 1286–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klobučar, A.; Lipovac, I.; Žagar, N.; Mitrović-Hamzić, S.; Tešić, V.; Vilibić-Čavlek, T.; Merdić, E. First record and spreading of the invasive mosquito Aedes japonicus japonicus (Theobald, 1901) in Croatia. Med. Vet. Entomol. 2019, 33, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Merdić, E.; Vignjević, G.; Turić, N.; Bogojević, M.S.; Milas, J.; Vrućina, I.; Zahirović, Z. Mosquito survey during West Nile virus outbreak 2012 in northeast Croatia. Coll. Antropol. 2014, 38, 423–428. [Google Scholar] [PubMed]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Dahl, C.; Madon, M.; Kaiser, A. Mosquito and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010; 577p. [Google Scholar]
- Schaffner, F.; Angel, G.; Geoffroy, B.; Hervy, J.P.; Rhaiem, A.; Brunhes, J. The Mosquitoes of Europe—An Identification and Training Programme; CD Rom, IRD Editions & EID Méditerranée: Montpellier, France, 2001. [Google Scholar]
- Tang, Y.; Hapip, C.A.; Liu, B.; Fang, C.T. Highly sensitive TaqMan RT-PCR assay for detection and quantification of both lineages of West Nile virus RNA. J. Clin. Virol. 2006, 36, 177–182. [Google Scholar] [CrossRef]
- Nikolay, B.; Weidmann, M.; Dupressoir, A.; Faye, O.; Boye, C.S.; Diallo, M.; Sall, A.A. Development of a Usutu virus specific real-time reverse transcription PCR assay based on sequenced strains from Africa and Europe. J. Virol. Methods 2014, 197, 51–54. [Google Scholar] [CrossRef] [Green Version]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN Analysis of Metagenomic Data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Klobučar, A. Spreading and Vector Role of Invasive Mosquitoes Aedes albopictus and Aedes japonicus in the Northwestern Croatia. Ph.D. Thesis, University of Zagreb, Zagreb, Croatia, 2017. [Google Scholar]
- Urbanelli, S.; Bellini, R.; Carrieri, M.; Sallicandro, P.; Celli, G. Population structure of Aedes albopictus (Skuse): The mosquito which is colonizing Mediterranean countries. Heredity 2000, 84, 331–337. [Google Scholar] [CrossRef]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Puggiolia, A.; Bonilaurib, P.; Calzolari, M.; Lellib, D.; Carrieria, M.; Urbanellid, S.; Pudare, D.; Bellini, R. Does Aedes albopictus (Diptera: Culicidae) play any role in Usutu in Northern Italy? Experimental oral infection and field evidences. Acta Trop. 2017, 172, 192–196. [Google Scholar] [CrossRef] [PubMed]



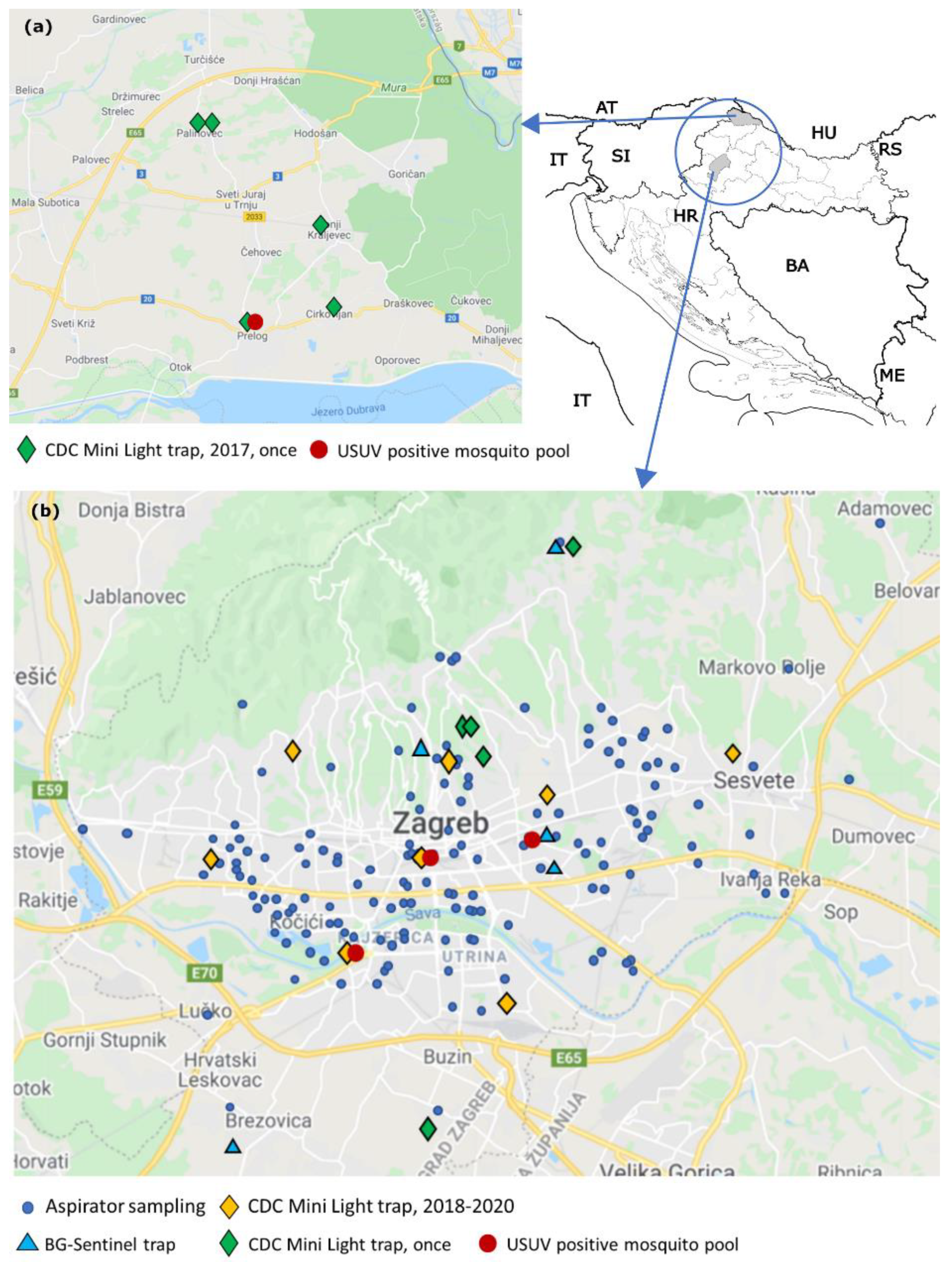{kind=link}
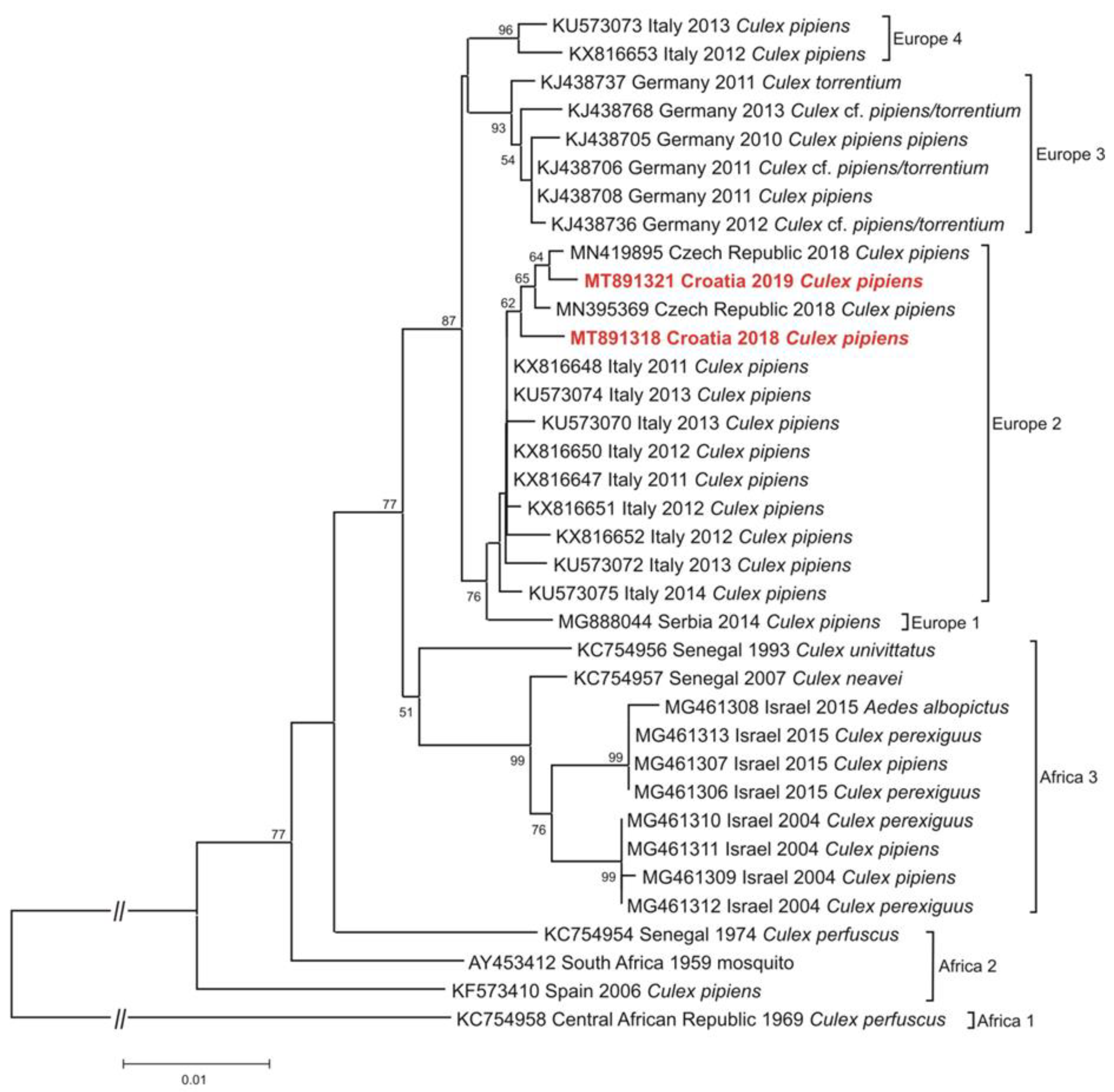{kind=link}
| Method of Collecting | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Total |
|---|---|---|---|---|---|---|---|
| CO2-baited trap | 7 | 75 | 96 | 96 | 274 | ||
| Aspirator | 116 | 23 | 57 | 41 | 43 | 51 | 331 |
| BG Sentinel | 13 | 13 | |||||
| Total | 129 | 23 | 64 | 116 | 139 | 147 | 618 |
| Mosquito Species | Sampling Method | Number of Collected Specimens per Species | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Subtotal | Total | % | ||
| City of Zagreb | ||||||||||
| Aedes albopictus | Aspirator | 1231 | 455 | 534 | 96 | 115 | 572 | 3003 | 4768 | 23.49 |
| CO2-baited trap | 380 | 619 | 537 | 1536 | ||||||
| BG Sentinel | 229 | 229 | ||||||||
| Aedes cinereus | Aspirator | 2 | 1 | 13 | 2 | 18 | 67 | 0.33 | ||
| CO2-baited trap | 49 | 49 | ||||||||
| Aedes rossicus | Aspirator | 18 | 18 | 338 | 1.67 | |||||
| CO2-baited trap | 320 | 320 | ||||||||
| Aedes vexans | Aspirator | 38 | 17 | 144 | 266 | 465 | 4359 | 21.48 | ||
| CO2-baited trap | 1 | 189 | 2290 | 1414 | 3894 | |||||
| Aedes japonicus | Aspirator | 2 | 2 | 2 | 0.01 | |||||
| Coquillettidia richiardii | CO2-baited trap | 8 | 8 | 8 | 0.04 | |||||
| Culex modestus | CO2-baited trap | 1 | 1 | 1 | 0.005 | |||||
| Culex pipiens complex | Aspirator | 475 | 53 | 345 | 2 | 257 | 1132 | 4210 | 20.75 | |
| CO2-baited trap | 20 | 1144 | 946 | 906 | 3016 | |||||
| BG Sentinel | 62 | 62 | ||||||||
| Ochlerotatus geniculatus | CO2-baited trap | 78 | 81 | 159 | 159 | 0.78 | ||||
| Ochlerotatus rusticus | Aspirator | 71 | 71 | 71 | 0.35 | |||||
| Ochlerotatus sticticus | Aspirator | 327 | 115 | 848 | 506 | 82 | 1878 | 6308 | 31.09 | |
| CO2-baited trap | 584 | 3541 | 305 | 4430 | ||||||
| Total | 2364 | 455 | 743 | 3670 | 8631 | 4428 | 20,291 | 20,291 | 100.00 | |
| Međimurje County | ||||||||||
| Culex pipiens complex | CO2-baited trap | 38 | 38 | 52.77 | ||||||
| Aedes vexans | CO2-baited trap | 30 | 30 | 41.66 | ||||||
| Ochlerotatus sticticus | CO2-baited trap | 4 | 4 | 5.55 | ||||||
| Total | 72 | 72 | 72 | 100.00 | ||||||
| Mosquito Species | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | Total |
|---|---|---|---|---|---|---|---|
| City of Zagreb | |||||||
| Aedes albopictus | 42 | 19 * | 27 | 32 | 49 | 54 | 223 |
| Aedes cinereus | 1 | 2 | 9 | 12 | |||
| Aedes rossicus | 17 | 17 | |||||
| Aedes vexans | 4 | 9 | 15 | 99 | 62 | 189 | |
| Aedes japonicus | 1 | 1 | |||||
| Coquillettidia richiardii | 2 | 2 | |||||
| Culex modestus | 1 | 1 | |||||
| Culex pipiens complex | 18 | 7 | 62 * | 55 * | 45 | 187 | |
| Ochlerotatus geniculatus | 6 | 7 | 13 | ||||
| Ochlerotatus rusticus | 9 | 9 | |||||
| Ochlerotatus sticticus | 14 | 9 | 57 | 136 | 23 | 239 | |
| Total | 79 | 19 | 52 | 177 | 372 | 193 | 893 |
| Međimurje County | |||||||
| Culex pipiens complex | 3 * | 3 | |||||
| Aedes vexans | 2 | 2 | |||||
| Ochlerotatus sticticus | 1 | 1 | |||||
| Total | 6 | 6 |
| Group | Order | Family/Genus (Species) | Number of Reads | Host |
|---|---|---|---|---|
| dsDNA | Caudovirales | Myoviridae | 39 | Bacteria |
| / | Nudiviridae | 64 | Insects | |
| ssRNA | Picornavirales | Picornaviridae (Culex picorna-like virus 1) | 12,067 | Mosquitoes |
| Nidovirales / | Mesoniviridae/Alphamesonivirus (Alphamesonivirus 1) Nodaviridae (Culex mosquito virus 4) | 2409 1171 | Mosquitoes Mosquitoes | |
| / | Flaviviridae/Flavivirus (Usutu virus) | 31 | Mammals and birds | |
| / | Unclassified RNA virus (Hubei chryso-like virus 1) | 191 | Mosquitoes | |
| / | Unclassified RNA virus (Wuhan Mosquito Virus 8) | 916 | Mosquitoes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klobucar, A.; Savic, V.; Curman Posavec, M.; Petrinic, S.; Kuhar, U.; Toplak, I.; Madic, J.; Vilibic-Cavlek, T. Screening of Mosquitoes for West Nile Virus and Usutu Virus in Croatia, 2015–2020. Trop. Med. Infect. Dis. 2021, 6, 45. https://doi.org/10.3390/tropicalmed6020045
Klobucar A, Savic V, Curman Posavec M, Petrinic S, Kuhar U, Toplak I, Madic J, Vilibic-Cavlek T. Screening of Mosquitoes for West Nile Virus and Usutu Virus in Croatia, 2015–2020. Tropical Medicine and Infectious Disease. 2021; 6(2):45. https://doi.org/10.3390/tropicalmed6020045
Chicago/Turabian StyleKlobucar, Ana, Vladimir Savic, Marcela Curman Posavec, Suncica Petrinic, Urska Kuhar, Ivan Toplak, Josip Madic, and Tatjana Vilibic-Cavlek. 2021. "Screening of Mosquitoes for West Nile Virus and Usutu Virus in Croatia, 2015–2020" Tropical Medicine and Infectious Disease 6, no. 2: 45. https://doi.org/10.3390/tropicalmed6020045
APA StyleKlobucar, A., Savic, V., Curman Posavec, M., Petrinic, S., Kuhar, U., Toplak, I., Madic, J., & Vilibic-Cavlek, T. (2021). Screening of Mosquitoes for West Nile Virus and Usutu Virus in Croatia, 2015–2020. Tropical Medicine and Infectious Disease, 6(2), 45. https://doi.org/10.3390/tropicalmed6020045