
Echinococcus multilocularis Calreticulin Interferes with C1q-Mediated Complement Activation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sera
2.3. Parasites, Antigen and Recombinant Protein Preparations
2.4. Cell Culture
2.5. Binding of EmCRT to Human C1q
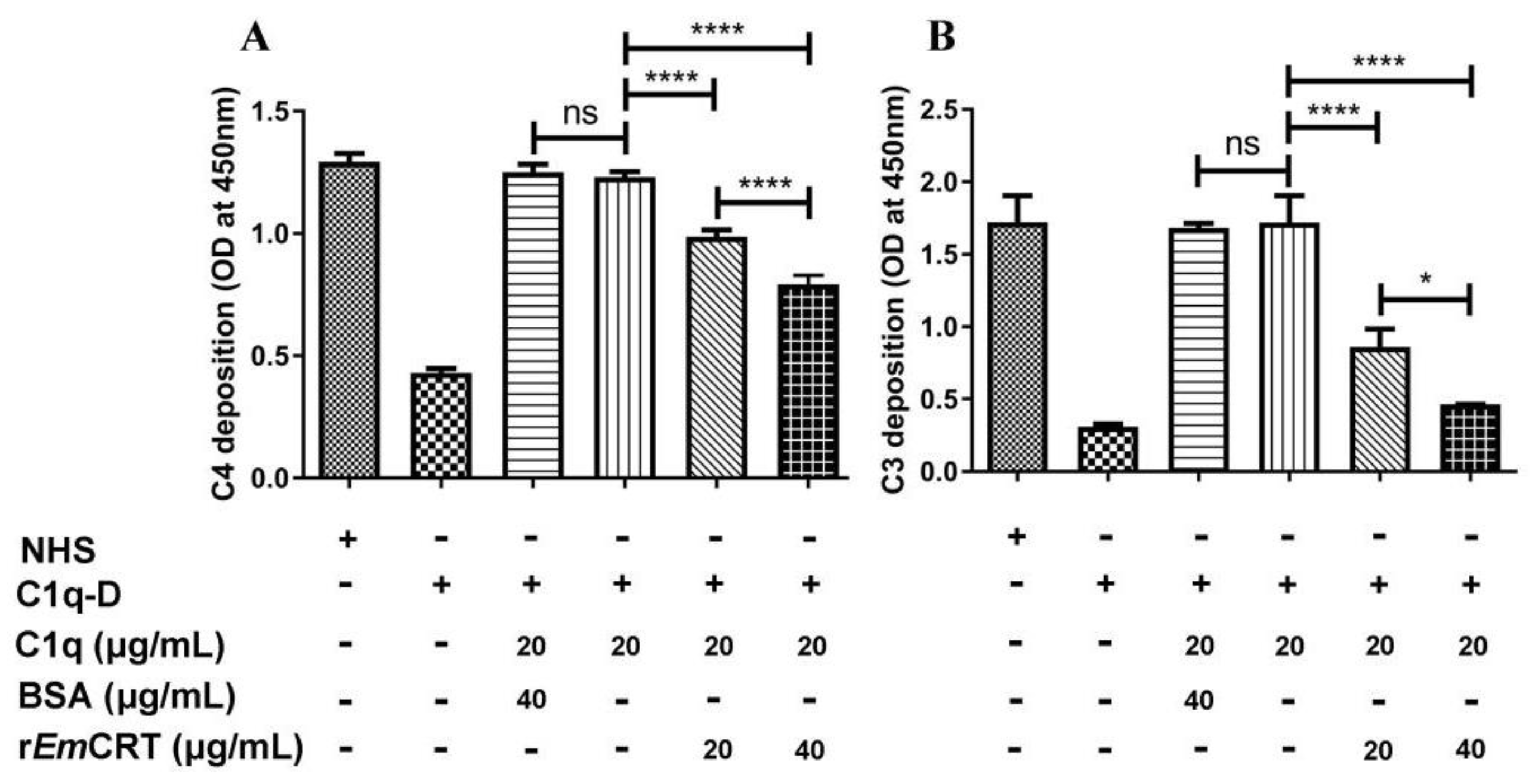
2.6. C3 and C4 Deposition Assay
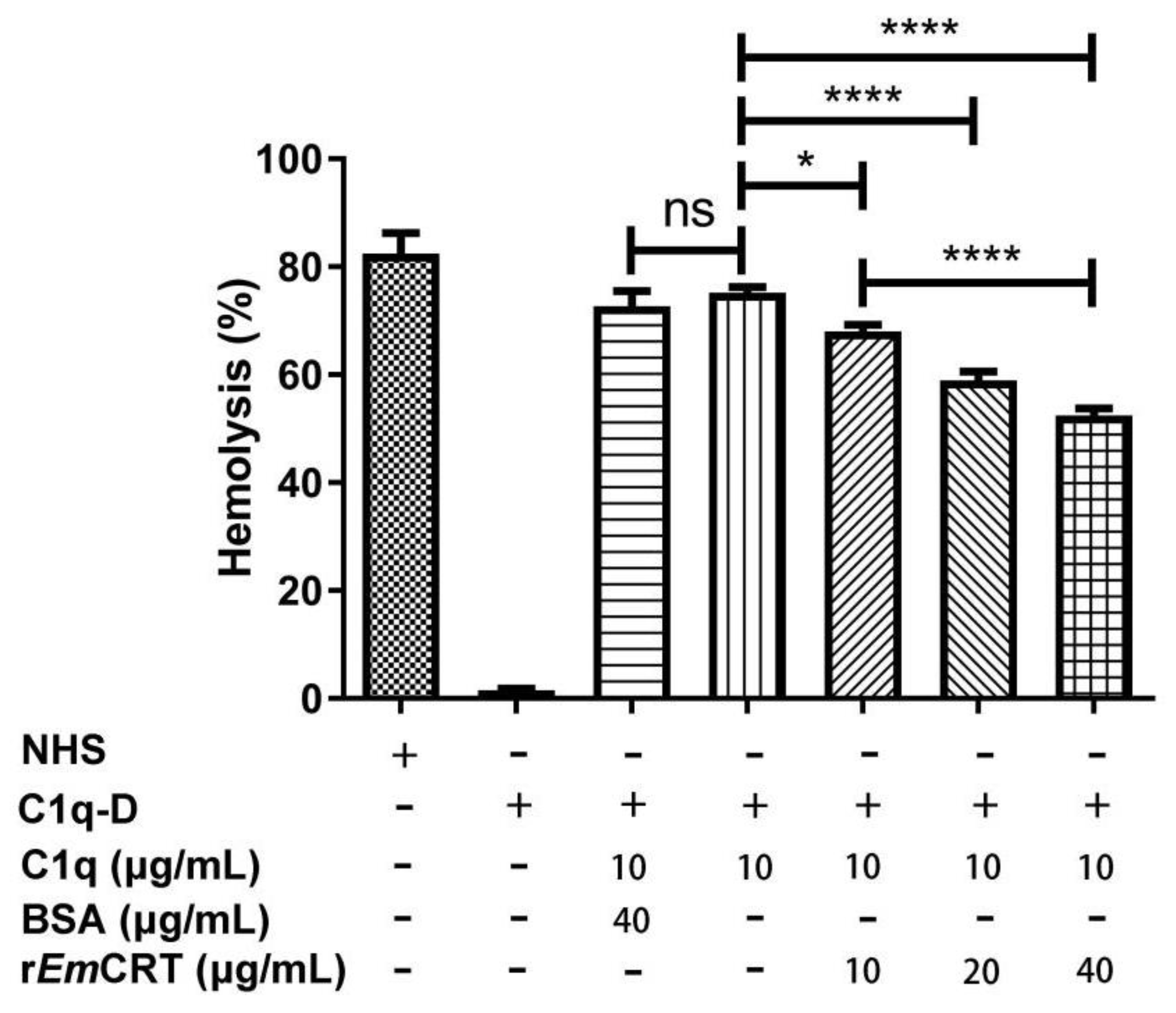
2.7. Hemolytic Assays
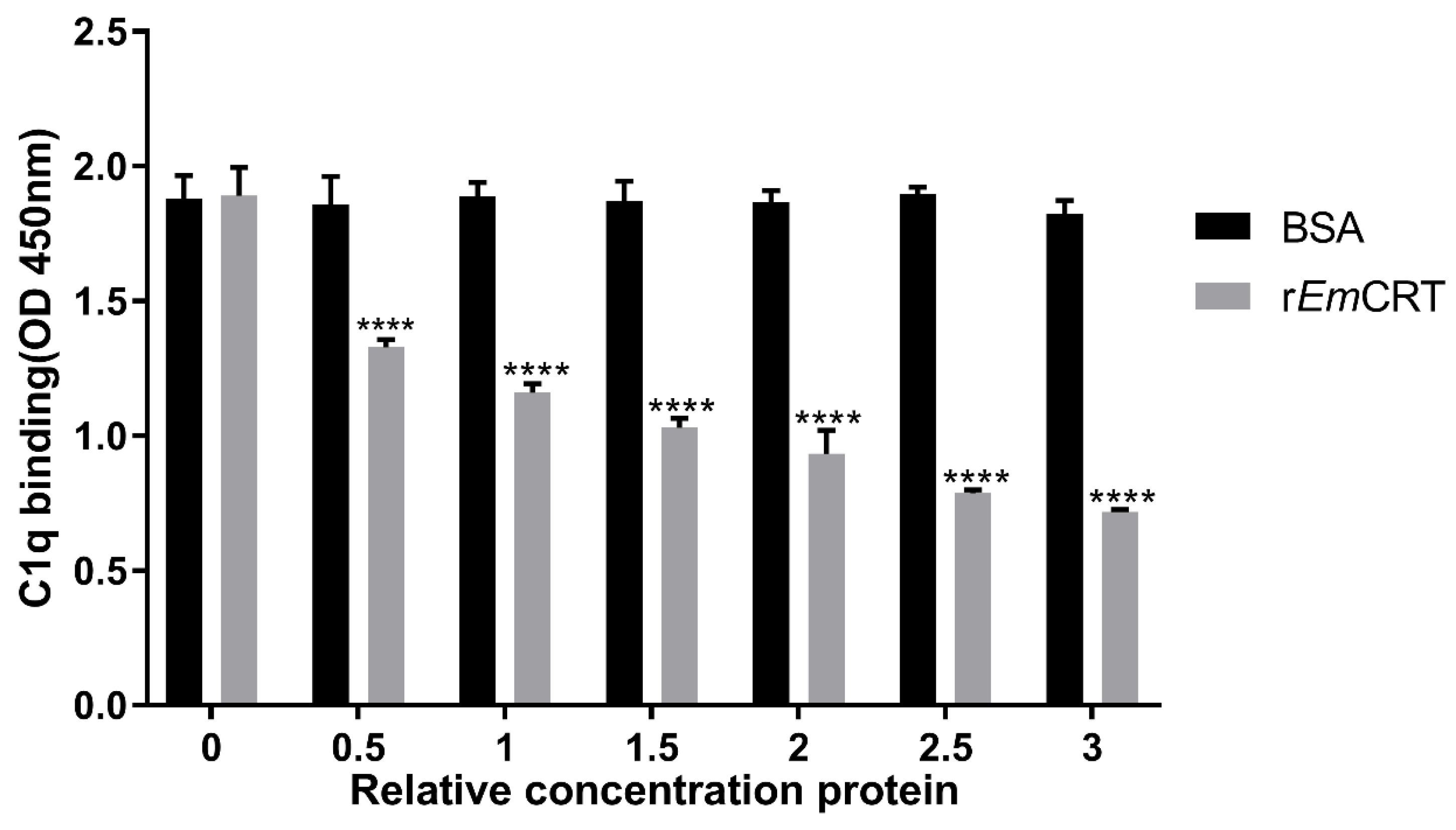
2.8. Inhibition of rEmCRT on the Binding of C1q to IgM
2.9. Cell Immunofluorescence Labeling
2.10. Transwell Chemotaxis Assay
2.11. Statistical Analysis
3. Results
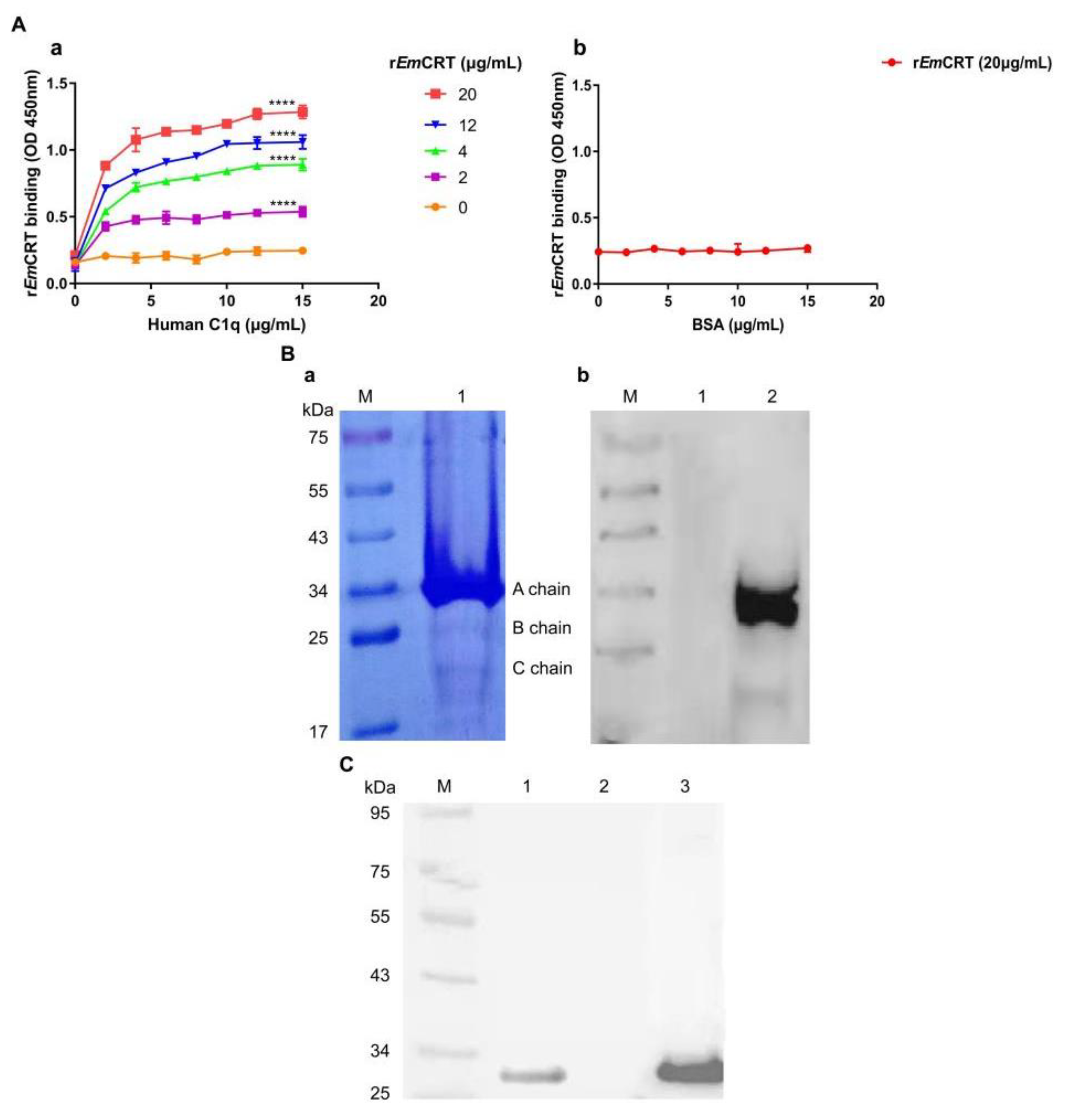
3.1. Recombinant EmCRT Binds to Human C1q
3.2. Native EmCRT from Worm Extracts Binds to Human C1q
3.3. rEmCRT Inhibits the Classical Complement Activation Pathway and Hemolysis
3.4. rEmCRT Competitively Inhibits the Binding of Human C1q to IgM
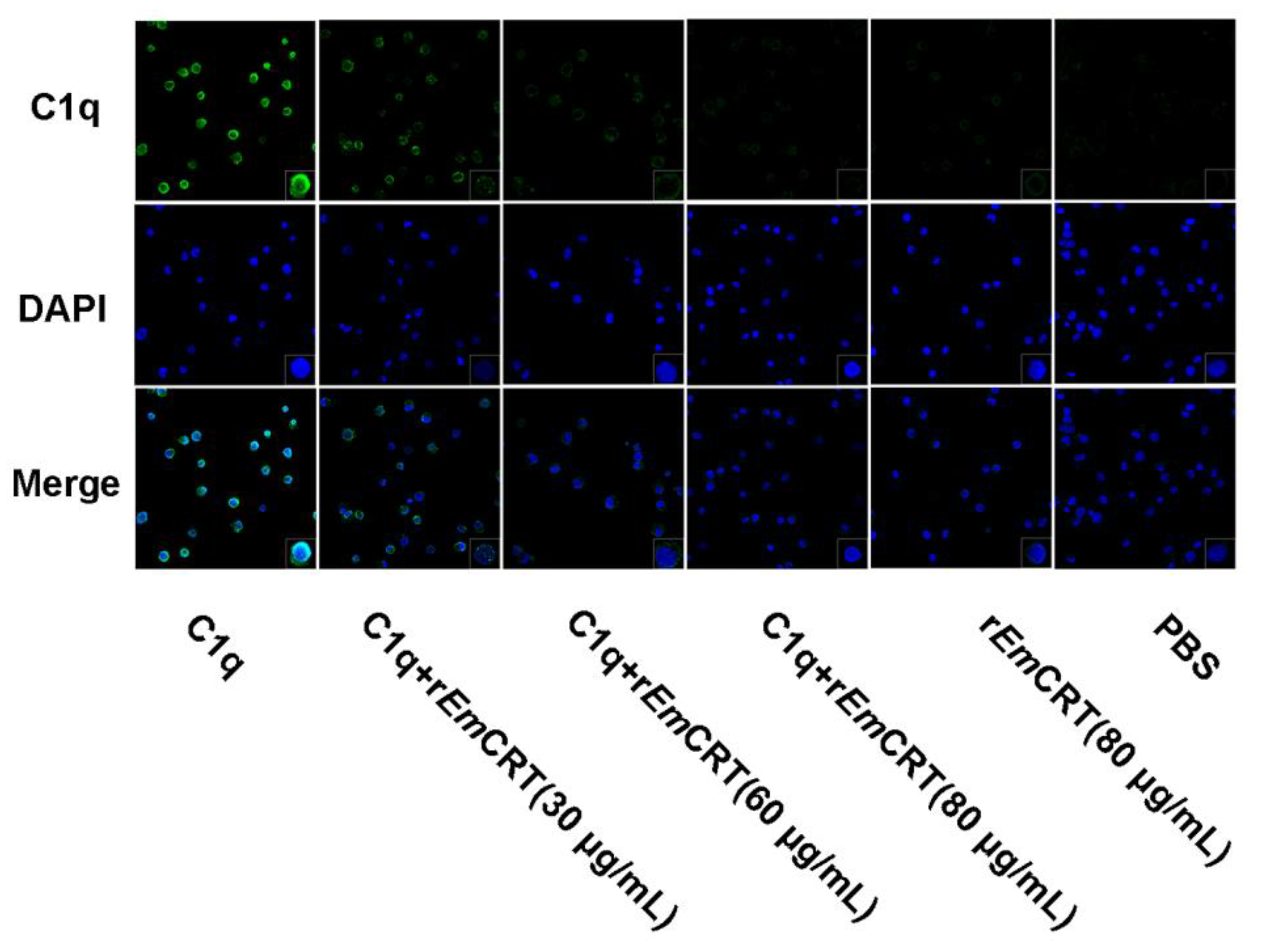
3.5. rEmCRT Inhibits C1q Binding to Mast Cells
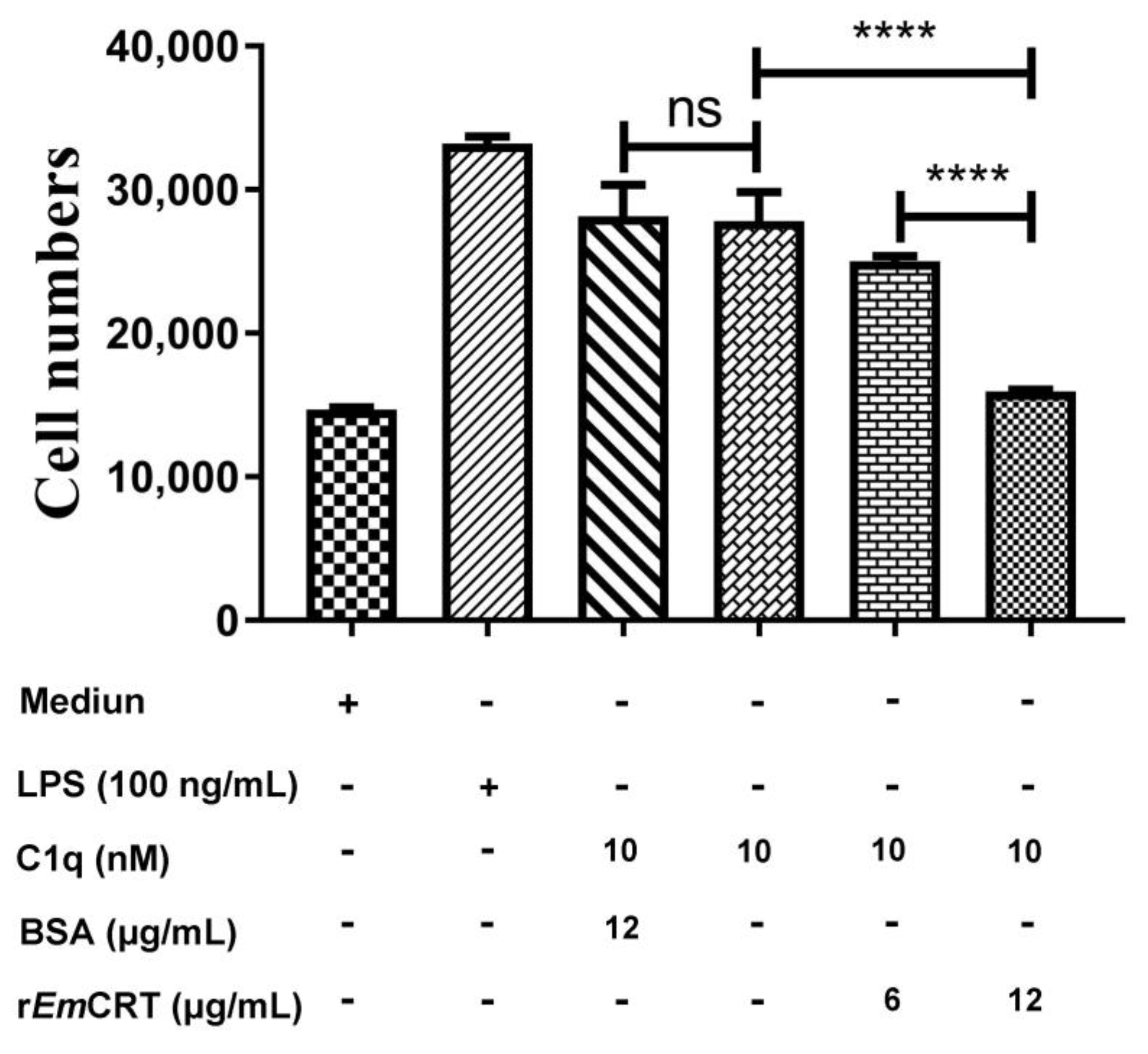
3.6. rEmCRT Inhibits C1q-Induced Mast Cells Chemotaxis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taratuto, A.L.; Venturiello, S.M. Echinococcosis. Brain Pathol. 1997, 7, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, P.R.; Keller, K.; Magnotta, M.; Ragland, N. The global burden of alveolar echinococcosis. PLoS Negl. Trop. Dis. 2010, 4, e722. [Google Scholar] [CrossRef]
- McManus, D.P.; Zhang, W.; Li, J.; Bartley, P.B. Echinococcosis. Lancet 2003, 362, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Vuitton, D.A.; McManus, D.P.; Rogan, M.T.; Romig, T.; Gottstein, B.; Naidich, A.; Tuxun, T.; Wen, H.; Menezes da Silva, A.; World Association of Echinococcosis. International consensus on terminology to be used in the field of echinococcoses. Parasite 2020, 27, 41. [Google Scholar] [CrossRef] [PubMed]
- Kassis, A.I.; Tanner, C.E. Echinococcus multilocularis: Complement’s role in vivo in hydatid disease. Exp. Parasitol. 1977, 43, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Madhavan, M.; Call, M.K.; Santiago, W.; Tsonis, P.A.; Lambris, J.D.; Del, R.T.K. Expression of complement 3 and complement 5 in newt limb and lens regeneration. J. Immunol. 2003, 170, 2331–2339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunkelberger, J.R.; Song, W.C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holers, V.M. Complement and its receptors: New insights into human disease. Annu. Rev. Immunol. 2014, 32, 433–459. [Google Scholar] [CrossRef]
- Lambris, J.D.; Ricklin, D.; Geisbrecht, B.V. Complement evasion by human pathogens. Nat. Rev. Microbiol. 2008, 6, 132–142. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.; Liu, X.; Wang, Z.; Liu, W.; Ye, X. Influenza A virus M1 blocks the classical complement pathway through interacting with C1qA. J. Gen. Virol. 2009, 90, 2751–2758. [Google Scholar] [CrossRef]
- Garcia, B.L.; Zhi, H.; Wager, B.; Höök, M.; Skare, J.T. Borrelia burgdorferi BBK32 Inhibits the Classical Pathway by Blocking Activation of the C1 Complement Complex. PLoS Pathog. 2016, 12, e1005404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Sroka, M.; Fulde, M.; Bergmann, S.; Riesbeck, K.; Blom, A.M. Binding of Streptococcus pneumoniae endopeptidase O (PepO) to complement component C1q modulates the complement attack and promotes host cell adherence. J. Biol. Chem. 2014, 289, 15833–15844. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Zhao, X.; Wang, Z.; Yang, J.; Zhao, L.; Zhan, B.; Zhu, X. Trichinella spiralis Paramyosin Binds Human Complement C1q and Inhibits Classical Complement Activation. PLoS Negl. Trop. Dis. 2015, 9, e0004310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassis, A.I.; Tanner, C.E. The role of complement in hydatid disease: In vitro studies. Int. J. Parasitol. 1976, 6, 25–35. [Google Scholar] [CrossRef]
- Kassis, A.I.; Tanner, C.E. Host serum proteins in Echinococcus multilocularis: Complement activation via the classical pathway. Immunology 1977, 33, 1–9. [Google Scholar] [PubMed]
- Reid, K.B.M. Complement Component C1q: Historical Perspective of a Functionally Versatile, and Structurally Unusual, Serum Protein. Front. Immunol. 2018, 10, 764. [Google Scholar] [CrossRef] [Green Version]
- Nayak, A.; Pednekar, L.; Reid, K.B.; Kishore, U. Complement and non-complement activating functions of C1q: A prototypical innate immune molecule. Innate Immun. 2012, 18, 350–363. [Google Scholar] [CrossRef]
- Leigh, L.E.; Ghebrehiwet, B.; Perera, T.P.; Bird, I.N.; Strong, P.; Kishore, U.; Reid, K.B. C1q-mediated chemotaxis by human neutrophils: Involvement of gClqR and G-protein signalling mechanisms. Biochem. J. 1998, 330, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Kuna, P.; Iyer, M.; Peerschke, E.I.; Kaplan, A.P.; Reid, K.B.; Ghebrehiwet, B. Human C1q induces eosinophil migration. Clin. Immunol. Immunopathol. 1996, 81, 48–54. [Google Scholar] [CrossRef]
- Edelson, B.T.; Stricker, T.P.; Li, Z.; Dickeson, S.K.; Shepherd, V.L.; Santoro, S.A.; Zutter, M.M. Novel collectin/C1q receptor mediates mast cell activation and innate immunity. Blood 2006, 107, 143–150. [Google Scholar] [CrossRef]
- Halova, I.; Draberova, L.; Draber, P. Mast cell chemotaxis-chemoattractants and signaling pathways. Front. Immunol. 2012, 3, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, D.A.; Laust, A.K.; Nelson, E.L.; Tenner, A.J. C1q differentially modulates phagocytosis and cytokine responses during ingestion of apoptotic cells by human monocytes, macrophages, and dendritic cells. J. Immunol. 2009, 183, 6175–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schcolnik-Cabrera, A.; Oldak, B.; Juárez, M.; Cruz-Rivera, M.; Flisser, A.; Mendlovic, F. Calreticulin in phagocytosis and cancer: Opposite roles in immune response outcomes. Apoptosis 2019, 24, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Molina, M.C.; Valck, C.; Rojas, A.; Aguilar, L.; Ramírez, G.; Schwaeble, W.; Ferreira, A. Role of calreticulin from parasites in its interaction with vertebrate hosts. Mol. Immunol. 2004, 40, 1279–1291. [Google Scholar] [CrossRef]
- Ramírez-Toloza, G.; Sosoniuk-Roche, E.; Valck, C.; Aguilar-Guzmán, L.; Ferreira, V.P.; Ferreira, A. Trypanosoma cruzi Calreticulin: Immune Evasion, Infectivity, and Tumorigenesis. Trends Parasitol. 2020, 36, 368–381. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Gupta, S.; Selvara, j.C.; Doharey, P.K.; Verma, A.; Singh, S.K.; Saxena, J.K. In silico and in vitro studies on the protein-protein interactions between Brugia malayi immunomodulatory protein calreticulin and human C1q. PLoS ONE 2014, 9, e106413. [Google Scholar] [CrossRef] [Green Version]
- Suchitra, S.; Joshi, P. Characterization of Haemonchus contortus calreticulin suggests its role in feeding and immune evasion by the parasite. Biochim. Biophys. Acta 2005, 722, 293–303. [Google Scholar] [CrossRef]
- Kasper, G.; Brown, A.; Eberl, M.; Vallar, L.; Kieffer, N.; Berry, C.; Girdwood, K.; Eggleton, P.; Quinnell, R.; Pritchard, D.I. A calreticulin-like molecule from the human hookworm Necator americanus interacts with C1q and the cytoplasmic signalling domains of some integrins. Parasite Immunol. 2001, 23, 141–152. [Google Scholar] [CrossRef]
- Chen, L.; Cheng, Z.; Xian, S.; Zhan, B.; Xu, Z.; Yan, Y.; Chen, J.; Wang, Y.; Zhao, L. Immunization with EmCRT-Induced Protective Immunity against Echinococcus multilocularis Infection in BALB/c Mice. Trop. Med. Infect. Dis. 2022, 7, 279. [Google Scholar] [CrossRef]
- Tang, C.T.; Quian, Y.C.; Kang, Y.M.; Cui, G.W.; Lu, H.C.; Shu, L.M.; Wang, Y.H.; Tang, L. Study on the ecological distribution of alveolar Echinococcus in Hulunbeier Pasture of Inner Mongolia, China. Parasitology 2004, 128, 187–194. [Google Scholar] [CrossRef]
- Spiliotis, M.; Brehm, K. Axenic in vitro cultivation of Echinococcus multilocularis metacestode vesicles and the generation of primary cell cultures. Methods Mol. Biol. 2009, 470, 245–262. [Google Scholar] [PubMed]
- Nayak, A.; Ferluga, J.; Tsolaki, A.G.; Kishore, U. The non-classical functions of the classical complement pathway recognition subcomponent C1q. Immunol. Lett. 2010, 131, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Groenendyk, J.; Wang, W.A.; Robinson, A.; Michalak, M. Calreticulin and the Heart. Cells. 2022, 11, 1722. [Google Scholar] [CrossRef]
- Fucikova, J.; Spisek, R.; Kroemer, G.; Galluzzi, L. Calreticulin and cancer. Cell Res. 2021, 31, 5–16. [Google Scholar] [CrossRef]
- Shao, S.; Hao, C.; Zhan, B.; Zhuang, Q.; Zhao, L.; Chen, Y.; Huang, J.; Zhu, X. Trichinella spiralis Calreticulin S-Domain Binds to Human Complement C1q to Interfere with C1q-Mediated Immune Functions. Front. Immunol. 2020, 11, 572326. [Google Scholar] [CrossRef]
- Chen, L.; Cheng, Z.; Wang, Y.; Zhao, L. Expression of Echinococcus Multilocularis Calreticulin and Analysis of Its Interaction with Complement Component C1q. Anim. Infect. Dis. 2021, 29, 40–46. [Google Scholar]
- Gadjeva, M.G.; Rouseva, M.M.; Zlatarova, A.S.; Reid, K.B.; Kishore, U.; Kojouharova, M.S. Interaction of human C1q with IgG and IgM: Revisited. Biochemistry 2008, 47, 13093–130102. [Google Scholar] [CrossRef] [PubMed]
- Dawicki, W.; Marshall, J.S. New and emerging roles for mast cells in host defence. Curr. Opin. Immunol. 2007, 19, 31–38. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xian, S.; Chen, L.; Yan, Y.; Chen, J.; Yu, G.; Shao, Y.; Zhan, B.; Wang, Y.; Zhao, L. Echinococcus multilocularis Calreticulin Interferes with C1q-Mediated Complement Activation. Trop. Med. Infect. Dis. 2023, 8, 47. https://doi.org/10.3390/tropicalmed8010047
Xian S, Chen L, Yan Y, Chen J, Yu G, Shao Y, Zhan B, Wang Y, Zhao L. Echinococcus multilocularis Calreticulin Interferes with C1q-Mediated Complement Activation. Tropical Medicine and Infectious Disease. 2023; 8(1):47. https://doi.org/10.3390/tropicalmed8010047
Chicago/Turabian StyleXian, Siqi, Lujuan Chen, Yan Yan, Jianfang Chen, Guixia Yu, Yuxiao Shao, Bin Zhan, Yanhai Wang, and Limei Zhao. 2023. "Echinococcus multilocularis Calreticulin Interferes with C1q-Mediated Complement Activation" Tropical Medicine and Infectious Disease 8, no. 1: 47. https://doi.org/10.3390/tropicalmed8010047
APA StyleXian, S., Chen, L., Yan, Y., Chen, J., Yu, G., Shao, Y., Zhan, B., Wang, Y., & Zhao, L. (2023). Echinococcus multilocularis Calreticulin Interferes with C1q-Mediated Complement Activation. Tropical Medicine and Infectious Disease, 8(1), 47. https://doi.org/10.3390/tropicalmed8010047