

Antileishmanial Activity and In Silico Molecular Docking Studies of Malachra alceifolia Jacq. Fractions against Leishmania mexicana Amastigotes

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Parasites and Cell Culture
2.3. Extract and Fractions Preparation
Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.4. In Vitro Anti-Amastigote Activity on L. mexicana pifanoi
2.5. Cytotoxicity Assay RAW 264.7 Cell
2.6. Molecular Docking
2.6.1. Prediction of Allosteric Binding Sites
2.6.2. Virtual Screening
2.6.3. Characterization of Mechanism of Allosteric Binding Sites
2.6.4. Pharmacological Profiling
3. Results and Discussion
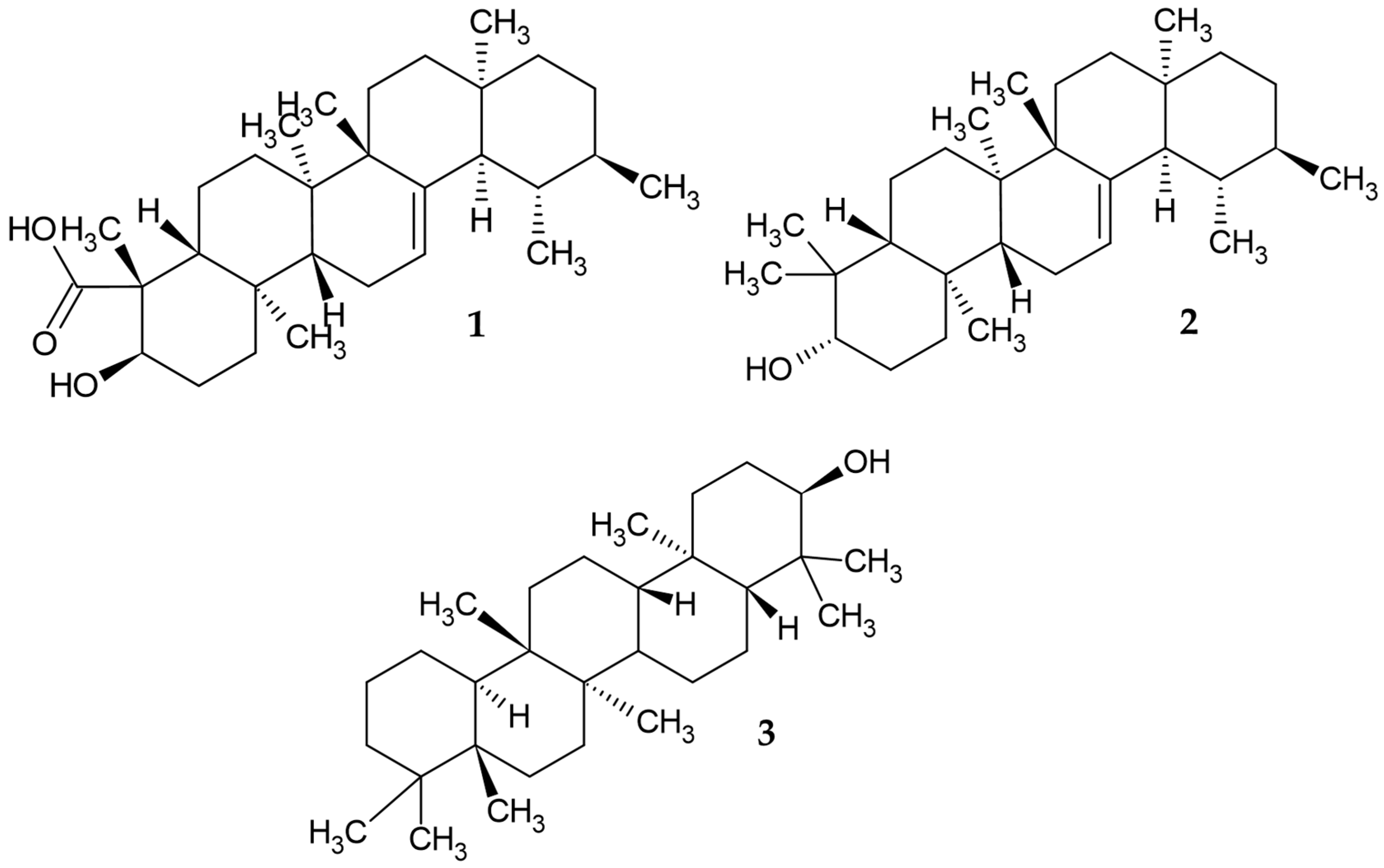
3.1. Phytochemical Screening for Thin-Layer Chromatography (TLC) and GC−MS Analysis of M. alceifolia Extract and Fraction
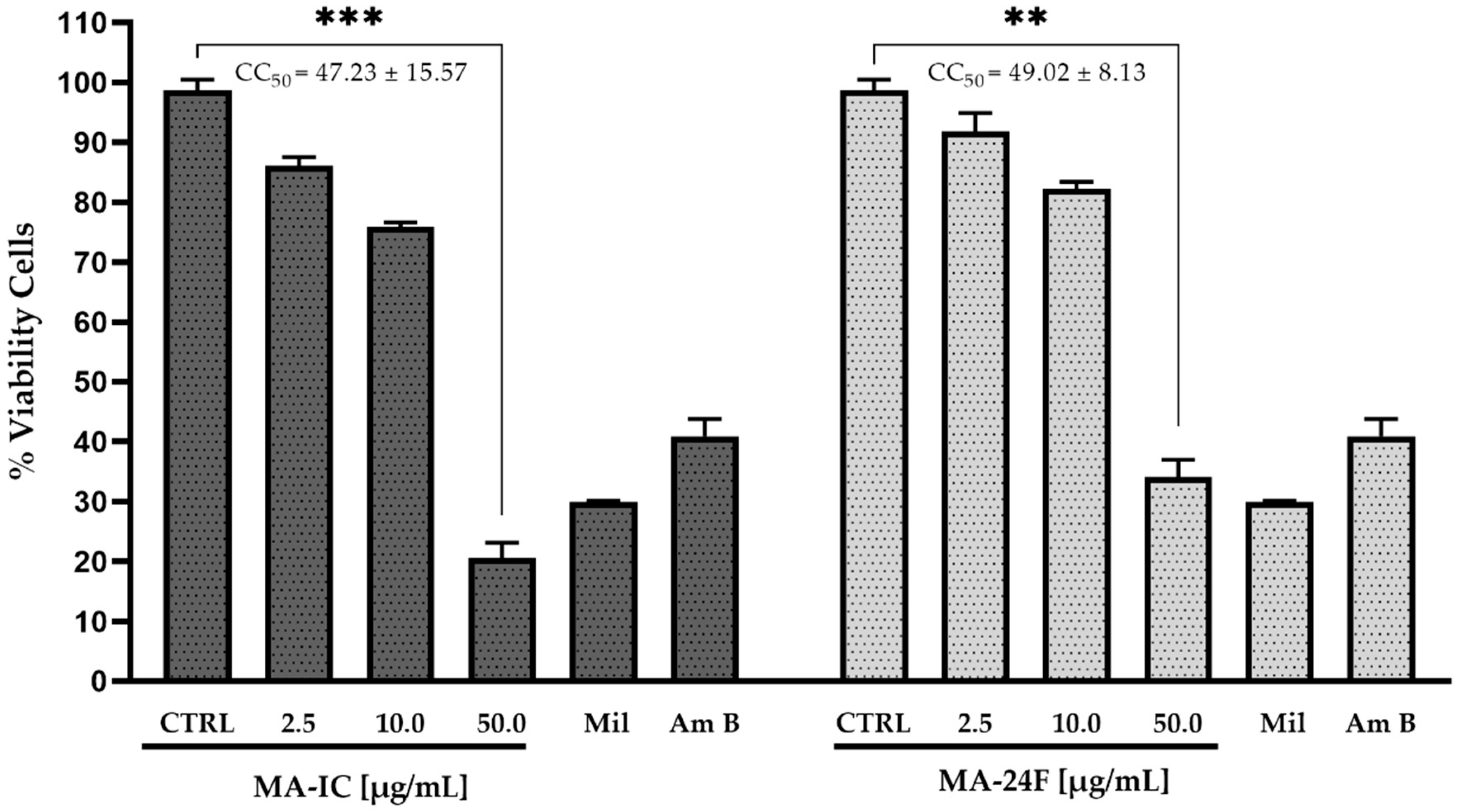
3.2. Antileishmanial and Cytotoxicity Activity of Fractions M. alceifolia axenic Amastigotes L. mexicana pifanoi and RAW 264.7 Macrophages
3.3. Molecular Docking Studies of M. alceifolia Active Fraction Ma-24F of Major Constituents with the Potential Drug Targets of L. mexicana
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cervantes-Ceballos, L.; Sánchez-Hoyos, J.; Sánchez-Hoyos, F.; Torres-Niño, E.; Mercado-Camargo, J.; Echeverry-Gómez, A.; Jotty Arroyo, K.; Del Olmo-Fernández, E.; Gómez-Estrada, H. An Overview of Genus Malachra L.-Ethnobotany, Phytochemistry, and Pharmacological Activity. Plants 2022, 11, 2808. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Estrada, H.; Díaz-Castillo, F.; Franco-Ospina, L.; Mercado-Camargo, J.; Guzmán-Ledezma, J.; Medina, J.D.; Gaitán-Ibarra, R. Folk medicine in the northern coast of Colombia: An overview. J. Ethnobiol. Ethnomed. 2011, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Kvist, L.; Christensen, S.; Rasmussen, H.; Mejia, K.; Gonzalez, A. Identification and evaluation of Peruvian plants used to treat malaria and leishmaniasis. J. Ethnopharmacol. 2006, 106, 390–402. [Google Scholar] [CrossRef] [PubMed]
- López-Cuesta, B. Studio Ethnobotanics of Plants against Malaria. 2017. Available online: http://147.96.70.122/web/TFG/TFG/Memoria/Beatriz%20Cuesta%20Lopez.pdf (accessed on 18 November 2022).
- World Health Organization. Leishmaniasis. 2021. Available online: https://www.who.int/data/gho/data/themes/topics/gho-ntd-leishmaniasis (accessed on 18 November 2022).
- World Health Organization. Leishmaniasis. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 18 November 2022).
- Ruiz, L.; Ruiz, L.; Maco, M.; Cobos, M.; Gutierrez-Choquevilca, A.-L.; Roumy, V. Plants used by native Amazonian groups from the Nanay River (Peru) for the treatment of malaria. J. Ethnopharmacol. 2011, 133, 917–921. [Google Scholar] [CrossRef]
- Metzdorf, I.P.; Junior, M.S.; Matos, M.D.; de Souza Filho, A.F.; de Souza Tsujisaki, R.A.; Franco, K.G.; Shapiro, J.T.; de Almeida Borges, F. Molecular characterization of Leishmania infantum in domestic cats in a region of Brazil endemic for human and canine visceral leishmaniasis. Acta Trop. 2017, 166, 121–125. [Google Scholar] [CrossRef]
- Instituto Nacional de Salud. Boletín Epidemiológico Semanal. Semana Epidemiológica 25. 2022. Available online: https://www.ins.gov.co/buscador-eventos/BoletinEpidemiologico/2022_Bolet%C3%ADn_epidemiologico_semana_25.pdf (accessed on 18 November 2022).
- Kaur, G.; Rajput, B. Comparative analysis of the omics technologies used to study antimonial, amphotericin B, and pentamidine resistance in leishmania. J. Parasitol. Res. 2014, 2014, 726328. [Google Scholar] [CrossRef]
- Silva-Almeida, M.; Pereira, B.A.; Ribeiro-Guimarães, M.L.; Alves, C.R. Proteinases as virulence factors in Leishmania spp. infection in mammals. Parasites Vectors 2012, 7, 160. [Google Scholar] [CrossRef]
- Silva-Almeida, M.; Souza-Silva, F.; Pereira, B.A.; Ribeiro-Guimaraes, M.L.; Alves, C.R. Overview of the organization of protease genes in the genome of Leishmania spp. Parasites Vectors 2014, 7, 387. [Google Scholar] [CrossRef]
- Chawla, B.; Madhubala, R. Drug targets in Leishmania. J. Parasit. Dis. 2010, 34, 1–13. [Google Scholar] [CrossRef]
- Berman, H.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Padilla, C.A.; Alvarez, M.J.; Combariza, A.F. Leishmania Proteomics: An in Silico Perspective. Preprints 2019, 4, 1–36. [Google Scholar] [CrossRef]
- Cervantes-Ceballos, L.; Sánchez-Hoyos, F.; Gómez-Estrada, H. Antibacterial activity of Cordia dentata Poir, Heliotropium indicum Linn and Momordica charantia Linn from the Northern Colombian Coast. Rev. Colomb. Cienc. Químico-Farm. 2017, 42, 143–159. [Google Scholar] [CrossRef]
- Sen, R.; Chatterjee, M. Plant derived therapeutics for the treatment of Leishmaniasis. Phytomedicine 2011, 18, 1056–1069. [Google Scholar] [CrossRef] [PubMed]
- Vargas, E.; Echeverri, F.; Upegui, Y.A.; Robledo, S.M.; Quiñones, W. Hydrazone Derivatives Enhance Antileishmanial Activity of Thiochroman-4-ones. Molecules 2017, 23, 70. [Google Scholar] [CrossRef]
- Luque-Ortega, J.R.; Rivas, L. Characterization of the Leishmanicidal Activity of Antimicrobial Peptides. In Antimicrobial Peptides: Methods in Molecular Biology; Giuliani, A., Rinaldi, A., Eds.; Humana Press: Totowa, NJ, USA, 2010; Volume 618. [Google Scholar] [CrossRef]
- Robledo, S.; Osorio, E.; Muñoz, D.; Jaramillo, L.M.; Restrepo, A.; Arango, G.; Vélez, I. In vitro and in vivo cytotoxicities and antileishmanial activities of thymol and hemisynthetic derivatives. Antimicrob. Agents Chemother. 2005, 49, 1652–1655. [Google Scholar] [CrossRef] [PubMed]
- Coa, J.C.; García, E.; Carda, M.; Agut, R.; Vélez, I.D.; Muñoz, J.A.; Yepes, L.M.; Robledo, S.M.; Cardona, W.I. Synthesis, leishmanicidal, trypanocidal and cytotoxic activities of quinoline-chalcone and quinoline-chromone hybrids. Med. Chem. Res. 2017, 26, 1405–1414. [Google Scholar] [CrossRef]
- Upegui-Zapata, Y.A.; Echeverri, F.; Quiñones, W.; Torres, F.; Nacher, M.; Rivas, L.I.; Meira, C.D.S.; Gedamu, L.; Escobar, G.; Archbold, R.; et al. Mode of action of a formulation containing hydrazones and saponins against leishmania spp. Role in mitochondria, proteases and reinfection process. Int. J. Parasitol. Drugs Drug Resist. 2020, 13, 94–106. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Mercado-Camargo, J.; Cervantes-Ceballos, L.; Vivas-Reyes, R.; Pedretti, A.; Serrano-García, M.L.; Gómez-Estrada, H. Homology modeling of leishmanolysin (gp63) from Leishmania panamensis and molecular docking of flavonoids. ACS Omega 2020, 5, 14741–14749. [Google Scholar] [CrossRef]
- BIOVIA. Discovery Studio Modeling, Software Version; Dassault Systèmes: San Diego, CA, USA, 2017. [Google Scholar]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Samal, D. In Silico Homology Modeling and Docking Study of Translationally Controlled Tumor Protein of Labeorohita. Int. J. For. Fish. Res. 2017, 1, 8–22. [Google Scholar] [CrossRef]
- Liu, Y.; Grimm, M.; Dai, W.T.; Hou, M.C.; Xiao, Z.X.; Cao, Y. CB-Dock: A web server for cavity detection-guided protein-ligand blind docking. Acta Pharmacol. Sin. 2020, 41, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, L. Improved protein-ligand binding affinity prediction by using a curvature-dependent surface-area model. Bioinformatics 2014, 30, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- Volkamer, A.; Kuhn, D.; Rippmann, F.; Rarey, M. DoGSiteScorer: A web server for automatic binding site prediction, analysis and druggability assessment. Bioinformatics 2012, 28, 2074–2075. [Google Scholar] [CrossRef]
- Michel, M.; Homan, E.J.; Wiita, E.; Pedersen, K.; Almlöf, I.; Gustavsson, A.L.; Lundbäck, T.; Helleday, T.; Warpman Berglund, U. In silico druggability assessment of the NUDIX hydrolase protein family as a workflow for target prioritization. Front. Chem. 2020, 8, 443. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fu, A.; Zhang, L. An Overview of Scoring Functions Used for Protein–Ligand Interactions in Molecular Docking. Interdiscip. Sci. Comput. Life Sci. 2019, 11, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Almahmoud, S.; Zhong, H.A. Molecular Modeling of Allosteric Site of Isoform-Specific Inhibition of the Peroxisome Proliferator-Activated Receptor PPARγ. Biomolecules 2022, 12, 1614. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand À Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- Couillaud, J.; Leydet, L.; Duquesne, K.; Iacazio, G. The Terpene Mini-Path, a New Promising Alternative for Terpenoids Bio-Production. Genes 2021, 12, 1974. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.K.; Moo, C.L.; Song, A.A.; Chong, C.M.; Chong, C.W.; Abushelaibi, A.; Lim, S.E.; Lai, K.S. Terpene Derivatives as a Potential Agent against Antimicrobial Resistance (AMR) Pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef]
- Raimundo, V.D.; Carvalho, R.P.R.; Machado-Neves, M.; Marques-da-Silva, E.A. Effects of terpenes in the treatment of visceral leishmaniasis: A systematic review of preclinical evidence. Pharmacol. Res. 2022, 177, 106117. [Google Scholar] [CrossRef] [PubMed]
- Guerreo, T.; Vejarano, P.; Ochoa, R. Tamizaje fitoquímico y cuantificación de flavonoides totales de las hojas y flores de Malachra alceifolia Jacq. Ethnobot. Res. Appl. 2018, 4, 70–75. [Google Scholar]
- Camargos, H.S.; Silva, A.H.M.; Anjos, J.L.V.; Alonso, A. Molecular dynamics and partitioning of di-tert-butyl nitroxide in stratum corneum membranes: Effect of terpenes. Lipids 2010, 45, 419–427. [Google Scholar] [CrossRef]
- Youssefi, M.R.; Moghaddas, E.; Tabari, M.A.; Moghadamnia, A.A.; Hosseini, S.M.; Farash, B.R.; Ebrahimi, M.A.; Mousavi, N.N.; Fata, A.; Maggi, F.; et al. In vitro and in vivo effectiveness of carvacrol, thymol and linalool against Leishmania infantum. Molecules 2019, 24, 2072. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Adhikary, A.; Al-Ahdal, M.N.; Roy, S.; Kishore, U. Host–Pathogen Interaction in Leishmaniasis: Immune Response and Vaccination Strategies. Immuno 2022, 2, 218–254. [Google Scholar] [CrossRef]
- Das, A.; Jawed, J.J.; Das, M.C.; Sandhu, P.; De, U.C.; Dinda, B.; Akhter, Y.; Bhattacharjee, S. Antileishmanial and immunomodulatory activities of lupeol, a triterpene compound isolated from Sterculia villosa. Int. J. Antimicrob. Agents 2017, 50, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Biswas, M.; Haldar, P.K. The triterpenoid fraction from Trichosanthes dioica root exhibits in vitro antileishmanial effect against Leishmania donovani promastigotes. Pharmacogn. Res. 2013, 2, 109–112. [Google Scholar] [CrossRef]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2016, 22, 58. [Google Scholar] [CrossRef]
- Greve, H.L.; Kaiser, M.; Mäser, P.; Schmidt, T.J. Boswellic Acids Show In Vitro Activity against Leishmania donovani. Molecules 2021, 26, 3651. [Google Scholar] [CrossRef]
- Montaser, M.M.; El-Sharnouby, M.E.; El-Noubi, G.; El-Shaer, H.M.; Khalil, A.A.; Hassanin, M.; Amer, S.A.; El-Araby, D.A. Boswellia serrata Resin Extract in Diets of Nile Tilapia, Oreochromis niloticus: Effects on the Growth, Health, Immune Response, and Disease Resistance to Staphylococcus aureus. Animals 2021, 11, 446. [Google Scholar] [CrossRef]
- Pramanik, P.; Chakraborti, S.; Bagchi, A.; Chakraborti, T. Bioassay-based Corchorus capsularis L. leaf-derived β-sitosterol exerts antileishmanial effects against Leishmania donovani by targeting trypanothione reductase. Sci. Rep. 2020, 10, 20440. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.H.; Michelon, A.; Banhuk, F.W.; Staffen, I.V.; Klein, E.J.; da Silva, E.A.; Menolli, R.A. Leishmanicidal, Trypanocidal and Antioxidant Activity of Amyrin-Rich Extracts from Eugenia pyriformis Cambess. Iran. J. Pharm. Res. 2020, 4, 343–353. [Google Scholar] [CrossRef]
- Ioset, J.-R. Natural Products for Neglected Diseases: A Review. Curr. Org. Chem. 2008, 8, 643–666. [Google Scholar] [CrossRef]
- Ali, R.; Tabrez, S.; Rahman, F.; Alouffi, A.S.; Alshehri, B.M.; Alshammari, F.A.; Alaidarous, M.A.; Banawas, S.; Dukhyil, A.A.B.; Rub, A. Antileishmanial Evaluation of Bark Methanolic Extract of Acacia nilotica: In Vitro and In Silico Studies. ACS Omega 2021, 6, 8548–8560. [Google Scholar] [CrossRef]
- Singh, N.; Mishra, B.B.; Bajpai, S.; Singh, R.K.; Tiwari, V.K. Natural product-based leads to fight against leishmaniasis. Bioorg. Med. Chem. 2014, 22, 18–45. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Caffrey, C.; Kelly, B.; Loke, P.; Sajid, M. Proteases in parasitic diseases. Annu. Rev. Pathol. 2006, 1, 497–536. [Google Scholar] [CrossRef]
- Elmahallawy, E.K.; Alkhaldi, A.A.M. Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite. Vet. Sci. 2021, 8, 33. [Google Scholar] [CrossRef]
- Cordeiro, A.T.; Michels, P.A.; Delboni, L.F.; Thiemann, O.H. The crystal structure of glucose-6-phosphate isomerase from Leishmania mexicana reveals novel active site features. Eur. J. Biochem. 2004, 271, 2765–2772. [Google Scholar] [CrossRef]
- Smith, B.O.; Picken, N.C.; Westrop, G.D.; Bromek, K.; Mottram, J.C.; Coombs, G.H. The structure of Leishmania mexicana ICP provides evidence for convergent evolution of cysteine peptidase inhibitors. J. Biol. Chem. 2006, 281, 5821–5828. [Google Scholar] [CrossRef]
- D’Antonio, E.L.; Ullman, B.; Roberts, S.C.; Dixit, U.G.; Wilson, M.E.; Hai, Y.; Christianson, D.W. Crystal structure of arginase from Leishmania mexicana and implications for the inhibition of polyamine biosynthesis in parasitic infections. Arch. Biochem. Biophys. 2013, 535, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Sahu, U.; Kumar, A.; Khare, P. Metabolic Pathways of Leishmania Parasite: Source of Pertinent Drug Targets and Potent Drug Candidates. Pharmaceutics 2022, 14, 1590. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, R.; Chen, Y.P. Potential therapeutic targets and the role of technology in developing novel antileishmanial drugs. Drug Discov. Today 2015, 20, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Kapil, S.; Singh, P.K.; Silakari, O. An update on small molecule strategies targeting leishmaniasis. Eur. J. Med. Chem. 2018, 157, 339–367. [Google Scholar] [CrossRef] [PubMed]
- Dorlo, T.P.; Balasegaram, M.; Beijnen, J.H.; de Vries, P.J. Miltefosine: A review of its pharmacology and therapeutic efficacy in the treatment of leishmaniasis. J. Antimicrob. Chemother. 2012, 67, 2576–2597. [Google Scholar] [CrossRef]
- de Morais, S.M.; Vila-Nova, N.S.; Bevilaqua, C.M.; Rondon, F.C.; Lobo, C.H.; Moura, A.D.; Sales, A.D.; Rodrigues, A.P.; de Figuereido, J.R.; Campello, C.C.; et al. Thymol and eugenol derivatives as potential antileishmanial agents. Bioorg. Med. Chem. 2014, 22, 6250–6255. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteases Targets L. mexicana | Ligand/Drugs | Allosteric GScore (Binding Energy kcal/mol−1) | Interacting Hydrophobic Residues | Interacting Hydrogen Bond |
|---|---|---|---|---|
| Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) * PDB: 1A7K | Episwertenol | −9.1± 0.1 | Ile13, Thr197, Ala198, Thr199, Cys166 | Thr111 |
| α-amiryn | −8.6 ± 0.1 | Arg12, Ile13, Thr197, Thr199, Ala135, Ala198 | Asp38, Met39 | |
| Amphotericin B | −9.3 ± 0.1 | Ile13, Met39, Arg92, Gly112, Ala135, Ser165, Gly22 | Cys166, Thr167, Thr225, Arg249, Asn335 | |
| Miltefosine | −4.5 ± 0.2 | Pro136, Cys166 | Thr199, Arg249 | |
| Pyruvate kinase * PDB ID: 1PKL | α-amiryn | −9.9 ± 0.1 | Gln42, Leu74, Val76, Glu438, Phe463, Ile41 | Ser439 |
| Episwertenol | −8.5 ± 0.1 | Arg19, Arg348, Leu351 | Arg19, Arg22 | |
| Methyl commate A | −8.3 ± 0.2 | Ile78, Ala347, Arg348, Glu438 | Glu348, Ser439 | |
| Amphotericin B | −9.1 ± 0.1 | Arg22, Ile 41, Gly44, Val76, Glu438, Val440, Phe463, Lys467 | Arg19, Ser46, Asn67, Asn432, Ser439, Glu462 | |
| Miltefosine | −4.9 ± 0.2 | Val76, Tyr18 | Ser439 | |
| Triose phosphate isomerase * PDB: 1AMK | Episwertenol | −8.3 ± 0.2 | Ile68, Ala69, Lys70, Phe74, Glu104, Ile108 | Arg98, Glu104, Lys112 |
| Amphotericin B | −6.9 ± 0.2 | Tyr101, Gly103, Thr105, Thr13, Gln133, Val169 | Arg99, Thr100, Glu104, Gln146 | |
| Miltefosine | −4.3 ± 0.2 | Ile68, Phe74, Ala69, Ile108 | Lys112, Arg98 | |
| Aldolase * PDB: 1EPX | α-amiryn | −8.6 ± 0.1 | Ala41, Glu44, Leu121, Leu280, Ala312, | |
| Methyl commate A | −8.3 ± 0.2 | Asp43, Glu44, Cys84, Leu121, Leu280, Ala312, Arg313, Lys239 | Asp43, Lys239, Ser286, Ala312, Arg313 | |
| Episwertenol | −8.2 ± 0.2 | Ala312, Arg313 | Arg52 | |
| Amphotericin B | −8.6 ± 0.1 | Glu44, Ser45, Leu121, Pro123, Gly130, Gln132, Lys162, Leu280, Ala312 | Asp43, Lys116, Lys156, Arg158, Glu199, Gly282 | |
| Miltefosine | −5.4 ± 0.2 | Leu280, Ala312, Arg313 | Lys156, Lys116, Arg158 | |
| Transketolase * PDB: 1R9J | α-amiryn | −8.2 ± 0.1 | Trp309, Val 55, Leu313, Phe 327, Val328, Met331, Arg332 | |
| Episwertenol | −8.2 ± 0.1 | Phe104, Pro52, Pro107, Arg 332 | Pro52, Asp53 | |
| Methyl commate A | −8.6 ± 0.1 | Phe104 | Met331 | |
| Amphotericin B | −9.6 ± 0.1 | Arg96, Asp53, Arg57, Arg103, Phe104, Asp420, Ala421, Asp 423, His450 | ||
| Miltefosine | −5.0 ± 0.1 | Phe104, Ile508 | Val109, Arg103, Asp423 | |
| Arginase ** PDB: 4ITY | Episwertenol | −8.7 ± 0.1 | Leu190, Val193, Leu201, His202, Ala208 | Lys198 |
| α-amiryn | −8.5 ± 0.1 | Val193, Lys198, Leu201, Ala208, Val331, Arg332, Trp369 | Arg191 | |
| Amphotericin B | −7.4 ± 0.2 | Pro27, His28, Asn152, Ala192, Asp194, Lys198, Met211, Val259, | Arg191, Val193, Arg260, Gly261 | |
| Miltefosine | −4.9 ± 0.2 | Lys198, Leu190, Leu201, Val193, Ala208, Ala207 | Ser210, his213 | |
| Cysteine peptidases A *** PDB: 2C34 | Methyl commate A | −8.1 ± 0.1 | Met35, Pro30, Asp45, Tyr58, Pro60, Pro95 | Cys56, Asp45, Lys57 |
| Episwertenol | −8.0 ± 0.1 | Pro30, Phe96 | Tyr58 | |
| Amphotericin B | −7.6 ± 0.2 | Thr31, Gly69, Val68 | ||
| Miltefosine | −4.4 ± 0.2 | Pro95, Met35, Phe96, Pro30, Pro60, Tyr58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cervantes-Ceballos, L.; Mercado-Camargo, J.; del Olmo-Fernández, E.; Serrano-García, M.L.; Robledo, S.M.; Gómez-Estrada, H. Antileishmanial Activity and In Silico Molecular Docking Studies of Malachra alceifolia Jacq. Fractions against Leishmania mexicana Amastigotes. Trop. Med. Infect. Dis. 2023, 8, 115. https://doi.org/10.3390/tropicalmed8020115
Cervantes-Ceballos L, Mercado-Camargo J, del Olmo-Fernández E, Serrano-García ML, Robledo SM, Gómez-Estrada H. Antileishmanial Activity and In Silico Molecular Docking Studies of Malachra alceifolia Jacq. Fractions against Leishmania mexicana Amastigotes. Tropical Medicine and Infectious Disease. 2023; 8(2):115. https://doi.org/10.3390/tropicalmed8020115
Chicago/Turabian StyleCervantes-Ceballos, Leonor, Jairo Mercado-Camargo, Esther del Olmo-Fernández, María Luisa Serrano-García, Sara M. Robledo, and Harold Gómez-Estrada. 2023. "Antileishmanial Activity and In Silico Molecular Docking Studies of Malachra alceifolia Jacq. Fractions against Leishmania mexicana Amastigotes" Tropical Medicine and Infectious Disease 8, no. 2: 115. https://doi.org/10.3390/tropicalmed8020115
APA StyleCervantes-Ceballos, L., Mercado-Camargo, J., del Olmo-Fernández, E., Serrano-García, M. L., Robledo, S. M., & Gómez-Estrada, H. (2023). Antileishmanial Activity and In Silico Molecular Docking Studies of Malachra alceifolia Jacq. Fractions against Leishmania mexicana Amastigotes. Tropical Medicine and Infectious Disease, 8(2), 115. https://doi.org/10.3390/tropicalmed8020115