



Application of Multilocus Sequence Typing for the Characterization of Leptospira Strains in Malaysia
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of Leptospira sp. and Dark-Field Microscopy Observation
2.3. DNA Extraction
2.4. Detection and Characterization of Leptospira sp. in Cultured and Non-Cultured Samples Using rrs2 Genes
2.5. Detection and Characterization of Leptospira sp. in Cultured and Non-Cultured Samples through the Mlst
3. Results
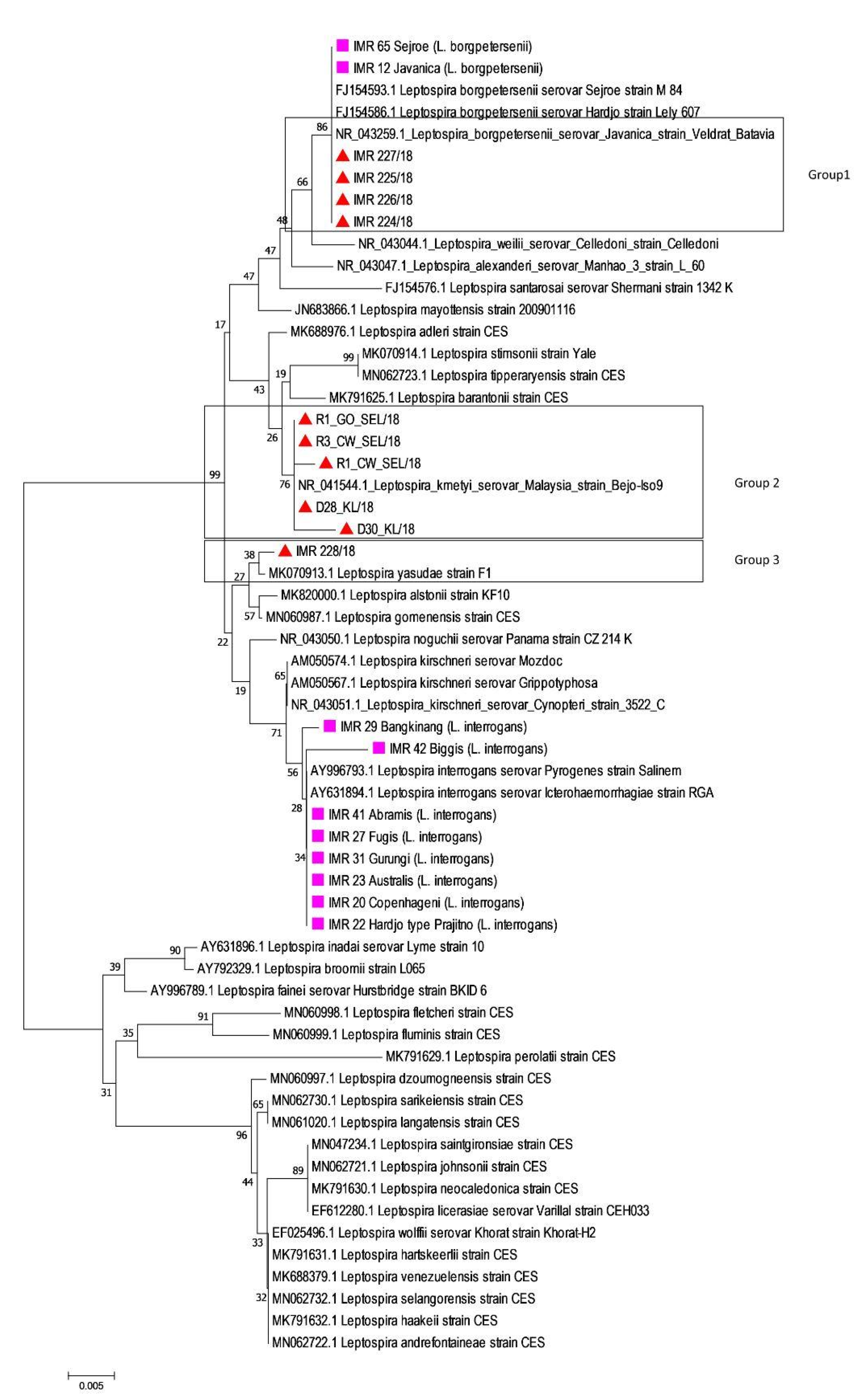
 ) (Figure 2). The rrs2 analysis showed a clustering of Leptospira into distinct species for both culture-positive and culture-negative samples. Four samples were identified as L. borgpetersenii, five samples were closely related to L. kmetyi and one sample was known as L. yasudae. Table 3 shows the estimates of the average evolutionary divergence over sequence pairs within the groups.
) (Figure 2). The rrs2 analysis showed a clustering of Leptospira into distinct species for both culture-positive and culture-negative samples. Four samples were identified as L. borgpetersenii, five samples were closely related to L. kmetyi and one sample was known as L. yasudae. Table 3 shows the estimates of the average evolutionary divergence over sequence pairs within the groups.4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Philip, N.; Bahtiar Affendy, N.; Ramli, S.N.A.; Arif, M.; Raja, P.; Nagandran, E.; Renganathan, P.; Taib, N.M.; Masri, S.N.; Yuhana, M.Y.; et al. Leptospira interrogans and Leptospira kirschneri are the dominant Leptospira species causing human leptospirosis in Central Malaysia. PLoS Negl. Trop. Dis. 2020, 14, e0008197. [Google Scholar] [CrossRef] [PubMed]
- Barnacle, J.; Gurney, S.; Ledot, S.; Singh, S. Leptospirosis as an important differential of pulmonary haemorrhage on the intensive care unit: A case managed with VV-ECMO. J. Intensive Care 2020, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, S.; Rattanavong, S.; Lee, S.J.; Panyanivong, P.; Craig, S.B.; Tulsiani, S.M.; Blacksell, S.D.; Dance, D.A.B.; FRCPath Dubot-Pérès, A.; Sengduangphachanh, A.; et al. Orientia, Rickettsia, and Leptospira pathogens as causes of CNS infections in Laos: A prospective study. Lancet Glob. Health 2015, 3, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Levett, P.N. Leptospirosis. Clin. Microbiol. 2015, 14, 296–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccarini, N.; Lutfi, S.; Piskorowski, T. Polyneuritis and rare sequelae of leptospirosis contracted while on an urban clean-up mission in Detroit: A case report of Weil’s disease and literature review. J. Med. Cases 2019, 10, 183–187. [Google Scholar] [CrossRef]
- Vincent, A.T.; Schiettekatte, O.; Goarant, C.; Neela, V.K.; Bernet, E.; Thibeaux, R.; Ismail, N.; Mohd Khalid, M.K.N.; Amran, F.; Masuzawa, T.; et al. Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl. Trop. Dis. 2019, 13, e0007270. [Google Scholar] [CrossRef] [Green Version]
- Casanovas-Massana, A.; Hamond, C.; Santos, L.A.; de Oliveira, D.; Hacker, K.P.; Balassiano, I.; Costa, F.; Medeiros, M.A.; Reis, M.G.; Ko, A.I.; et al. Leptospira yasudae sp. nov. and Leptospira stimsonii sp. nov., two new species of the pathogenic group isolated from environmental sources. Int. J. Syst. Evol. Microbiol. 2020, 70, 1450–1456. [Google Scholar]
- Picardeau, M. Virulence of the zoonotic agent of leptospirosis: Still terra incognita? Nat. Rev. Microbiol. 2017, 15, 297–307. [Google Scholar] [CrossRef]
- Thibeaux, R.; Girault, D.; Bierque, E.; Soupé-Gilbert, M.E.; Rettinger, A.; Douyère, A.; Meyer, M.; Iraola, G.; Picardeau, M.; Goarant, C. Biodiversity of environmental Leptospira: Improving identification and revisiting the diagnosis. Front. Microbiol. 2018, 9, 816. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yang, H.; Li, X.; Cao, Z.; Zhou, H.; Zeng, L.; Xu, J.; Xu, Y.; Chang, Y.F.; Guo, X.; et al. Molecular typing of pathogenic Leptospira serogroup Icterohaemorrhagiae strains circulating in China during the past 50 years. PLoS Negl. Trop. Dis. 2015, 9, e0003762. [Google Scholar] [CrossRef] [Green Version]
- Guernier, V.; Allan, K.J.; Goarant, C. Advances and challenges in barcoding pathogenic and environmental Leptospira. Parasitology 2018, 145, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [PubMed] [Green Version]
- Auch, A.F.; von Jan, M.; Klenk, H.P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, Y.I.; Rogozin, I.; Grishin, N.; Tatusov, R.; Koonin, E. Genome trees constructed using five different approaches suggest new major bacterial clades. BMC Evol. Biol. 2001, 1, 8. [Google Scholar] [CrossRef]
- Boonsilp, S.; Thaipadungpanit, J.; Amornchai, P.; Wuthiekanun, V.; Bailey, M.S.; Holden, M.T.G.; Zhang, C.; Jiang, X.; Koizumi, N.; Taylor, K.; et al. A single multilocus sequence typing (MLST) scheme for seven pathogenic Leptospira species. PLoS Negl. Trop. Dis. 2013, 7, e1954. [Google Scholar] [CrossRef] [Green Version]
- Leon, A.; Pronost, S.; Fortier, G.; Andre-Fontaine, G.; Leclercq, R. Multilocus sequence analysis for typing Leptospira interrogans and Leptospira kirschneri. J. Clin. Microbiol. 2010, 48, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Aanensen, D.M.; Spratt, B.G. The multilocus sequence typing network: Mlst.net. Nucleic Acids Res. 2005, 33, W728–W733. [Google Scholar] [CrossRef] [Green Version]
- Inouye, M.; Dashnow, H.; Raven, L.A.; Schultz, M.B.; Pope, B.J.; Tomita, T.; Zobel, J.; Holt, K.E. SRST2: Rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014, 6, 90. [Google Scholar] [CrossRef] [Green Version]
- Baust, J.G. Strategies for the storage of DNA. Biopreserv. Biobank. 2008, 6, 251–252. [Google Scholar]
- Ahmed, N.; Devi, S.M.; Valverde, M.L.V.P.; Machang’u, R.S.; Ellis, W.A.; Hartskeerl, R.A. Multilocus sequence typing method for identification and genotypic classification of pathogenic Leptospira species. Ann. Clin. Microbiol. Antimicrob. 2006, 23, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Thaipadungpanit, J.; Boonsilp, S.; Wuthiekanun, V.; Nalam, K.; Spratt, B.G.; Aaensen, D.M.; Smythe, L.D.; Ahmed, N.; Feil, E.J.; et al. Comparison of two multilocus sequence based genotyping schemes for Leptospira species. PLoS Negl. Trop. Dis. 2011, 5, e1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.I.; Goarant, C.; Picardeau, M. Leptospira: The dawn of the molecular genetics era for an emerging zoonotic pathogen. Nat. Rev. Microbiol. 2009, 7, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Engelberts, M.F.; Boer, K.R.; Ahmed, N.; Hartskeerl, R.A. Development and validation of a real-time PCR for detection of pathogenic Leptospira species in clinical materials. PLoS ONE 2009, 4, e7093. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.A.; Edwards, M.; Benschop, J.; Nisa, S. Identification of pathogenic Leptospira species and serovars in New Zealand using metabarcoding. PLoS ONE 2021, 16, e0257971. [Google Scholar] [CrossRef] [PubMed]
- Sykes, J.E.; Reagan, K.L.; Nally, J.E.; Galloway, R.L.; Haake, D.A. Role of diagnostics in epidemiology, management, surveillance, and control of leptospirosis. Pathogens 2022, 11, 395. [Google Scholar] [CrossRef]
- Thaipadungpanit, J.; Wuthiekanun, V.; Chierakul, W.; Smythe, L.D.; Petkanchanapong, W.; Limpaiboon, R.; Apiwatanaporn, A.; Slack, A.T.; Suputtamongkol, Y.; White, N.J.; et al. A dominant clone of Leptospira interrogans associated with human leptospirosis in Thailand. PLoS Negl. Trop. Dis. 2007, 1, e56. [Google Scholar] [CrossRef]
- Adler, B.; de la Peña Moctezuma, A. Leptospira and leptospirosis. Vet. Microbiol. 2010, 140, 287–296. [Google Scholar] [CrossRef]
- Slack, A.; Symonds, M.; Dohnt, M.; Harris, C.; Brookes, D.; Smythe, L. Evaluation of a modified Taqman assay detecting pathogenic Leptospira spp. against culture and Leptospira-specific IgM enzyme-linked immunosorbent assay in a clinical environment. Diagn. Microbiol. Infect. Dis. 2007, 57, 361–366. [Google Scholar] [CrossRef]
- Thaipadunpanit, J.; Chierakul, W.; Wuthiekanun, V.; Limmathurotsakul, D.; Amornchai, P.; Boonslip, S.; Smythe, L.D.; Limpaiboon, R.; Hoffmaster, A.R.; Day, N.P.J.; et al. Diagnostic accuracy of real-time PCR assays targeting 16S rRNA and lipl32 genes for human leptospirosis in Thailand: A case–control study. PLoS ONE 2011, 6, e16236. [Google Scholar] [CrossRef] [PubMed]
- Benacer, D.; Mohd Zain, S.N.; Amran, F.; Galloway, R.L.; Thong, K.L. Isolation and molecular characterization of Leptospira interrogans and Leptospira borgpetersenii isolates from the urban rat populations of Kuala Lumpur, Malaysia. Am. J. Trop. Med. Hyg. 2013, 88, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Blasdell, K.R.; Morand, S.; Perera, D.; Firth, C. Association of rodent-borne Leptospira spp. with urban environments in Malaysian Borneo. PLoS Negl. Trop. Dis. 2019, 13, e0007141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahtiar Affendy, N.; Mohd Desa, M.N.; Amran, F.; Sekawi, Z.; Masri, S.N. Isolation and molecular characterization of Leptospira interrogans and Leptospira borgpetersenii from small mammals in Selangor wet markets. Int. J. Infect. Dis. 2020, 101, 534. [Google Scholar] [CrossRef]
- Rahman, M.H.A.A.; Hairon, S.M.; Hamat, R.A.; Jamaluddin, T.Z.M.T.; Shafei, M.N.; Idris, N.; Osman, M.; Sukeri, S.; Wahab, Z.A.; Mohammad, W.M.Z.W.; et al. Seroprevalence and distribution of leptospirosis serovars among wet market workers in northeastern, Malaysia: A cross sectional study. BMC Infect. Dis. 2018, 18, 569. [Google Scholar] [CrossRef]
- Samsudin, S.; Saudi, S.N.S.; Masri, N.S.; Ithnin, N.R.; Jamaluddin, T.Z.M.T.; Hamat, R.A.; Wan Mohd, Z.W.M.; Nazri, M.S.; Surianti, S.; Daud, A.B.; et al. Awareness, knowledge, attitude and preventive practice of leptospirosis among healthy Malaysian and Non-Malaysian wet market workers in selected urban areas in Selangor, Malaysia. Int. J. Environ. Res. Public Health 2020, 17, 1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benacer, D.; Mohd Zain, S.N.; Shin, Z.S.; Mohd Khalid, M.K.N.; Galloway, R.L.; Souris, M.; Thong, K.L. Determination of Leptospira borgpetersenii serovar Javanica and Leptospira interrogans serovar Bataviae as the persistent Leptospira serovars circulating in the urban rat populations in Peninsular Malaysia. Parasites Vectors 2016, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Shafei, M.N.; Sulong, M.R.; Yaacob, N.A.; Hassan, H.; Wan Mohd Zahiruddin, W.M.; Daud, A.; Ismail, Z.; Abdullah, M.R. Seroprevalence of leptospirosis among town service workers on northeastern state of Malaysia. Int. J. Collab. Res. Intern. Med. Public Health 2012, 4, 395–403. [Google Scholar]
- Rafizah, N.A.A.; Aziah, B.D.; Azwany, Y.N.; Imran, M.K.; Mohamed Rusli, A.; Mohd Nazri, S.; Mohd Nikman, A.; Nabilah, I.; Siti Asma’, H.; Zahiruddin, W.M.; et al. A hospital-based study on seroprevalence of leptospirosis among febrile cases in northeastern Malaysia. Int. J. Infect. Dis. 2013, 17, e394–e397. [Google Scholar] [CrossRef] [Green Version]
- Boonsilp, S.; Thaipadungpanit, J.; Amornchai, P.; Wuthiekanun, V.; Chierakul, W.; Limmathurotsakul, D.; Day, N.P.; Peacock, S.J. Molecular detection and speciation of pathogenic Leptospira spp. in blood from patients with culture-negative leptospirosis. BMC Infect. Dis. 2011, 11, 338. [Google Scholar] [CrossRef] [Green Version]
- Agampodi, S.B.; Moreno, A.C.; Vinetz, J.M.; Matthias, M.A. Utility and limitations of direct multi-locus sequence typing on qPCR-positive blood to determine infecting Leptospira strain. Am. J. Trop. Med. Hyg. 2013, 88, 184–185. [Google Scholar] [CrossRef] [Green Version]
- Podgoršek, D.; Ružić-Sabljić, E.; Logar, M.; Pavlović, A.; Remec, T.; Baklan, Z.; Pal, E.; Cerar, T. Evaluation of real-time PCR targeting the lipL32 gene for diagnosis of Leptospira infection. BMC Microbiol. 2020, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Chiani, Y.; Jacob, P.; Varni, V.; Lamdolt, N.; Schmeling, M.F.; Pujato, N.; Caimi, K.; Vanasco, B. Isolation and clinical sample typing of human leptospirosis cases in Argentina. Infect. Genet. Evol. 2016, 37, 245–251. [Google Scholar] [CrossRef]
- Slack, A.T.; Khairani-Bejo, S.; Symonds, M.L.; Dohnt, M.F.; Galloway, R.L.; Steigerwalt, A.G.; Bahaman, A.R.; Craig, S.; Harrower, B.J.; Smythe, L.D. Leptospira kmetyi sp nov., isolated from an environmental source in Malaysia. Int. J. Syst. Evol. Microbiol. 2009, 59, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.T.; Amran, F.; Chee Cheong, K.; Ahmad, N. In-house ELISA screening using a locally isolated Leptospira in Malaysia: Determination of its cut-off points BMC Infect. Dis. 2014, 14, 563. [Google Scholar]
- Zaki, A.M.; Hod, R.; Shamsusah, N.A.; Isa, Z.M.; Bejo, S.K.; Agustar, H.K. Detection of Leptospira kmetyi at recreational areas in Peninsular Malaysia. Environ. Monit. Assess. 2020, 192, 703. [Google Scholar] [CrossRef]
- Shamsusah, N.A.; Agustar, H.K.; Amran, F.; Hod, R. Draft genome sequence of Leptospira yasudae strain BJ3, isolated from the soil of an ex-situ wild animal conservation area. Microbiol. Resour. Announc. 2021, 10, e0072321. [Google Scholar] [CrossRef]
- Voronina, O.L.; Kunda, M.S.; Aksenova, E.I.; Ryzhova, N.N.; Semenov, A.N.; Petrov, E.M.; Didenko, L.V.; Lunin, V.G.; Ananyina, Y.V.; Gintsburg, A.L. The characteristics of ubiquitous and unique Leptospira strains from the collection of Russian centre for leptospirosis. BioMed Res. Int. 2014, 2014, 649034. [Google Scholar] [CrossRef] [Green Version]
- Sykes, J.E.; Gamage, C.D.; Haake, D.A.; Nally, J.E. Understanding leptospirosis: Application of state-of-the-art molecular typing tools with a One Health lens. Am. J. Vet. Res. 2022, 83, ajvr.22.06.0104. [Google Scholar] [CrossRef]
- Techawiwattanaboon, T.; Patarakul, K. Update on molecular diagnosis of human leptospirosis. Asian Biomed. 2019, 13, 207–216. [Google Scholar] [CrossRef]
 ) and human (
) and human ( ) samples with reference strains from different countries based on the concatenated sequences of the six housekeeping genes using neighbor-joining phylogenetic trees with bootstraps of 100.
) and human () samples with reference strains from different countries based on the concatenated sequences of the six housekeeping genes using neighbor-joining phylogenetic trees with bootstraps of 100.
) samples with reference strains from different countries based on the concatenated sequences of the six housekeeping genes using neighbor-joining phylogenetic trees with bootstraps of 100.
) and human () samples with reference strains from different countries based on the concatenated sequences of the six housekeeping genes using neighbor-joining phylogenetic trees with bootstraps of 100.
 ) to old reference Malaysian Leptospira isolates (
) to old reference Malaysian Leptospira isolates ( ) and NCBI reference strains based on rrs2 genes using neighbor-joining phylogenetic trees with bootstraps of 100.
) to old reference Malaysian Leptospira isolates () and NCBI reference strains based on rrs2 genes using neighbor-joining phylogenetic trees with bootstraps of 100.
) and NCBI reference strains based on rrs2 genes using neighbor-joining phylogenetic trees with bootstraps of 100.
) to old reference Malaysian Leptospira isolates () and NCBI reference strains based on rrs2 genes using neighbor-joining phylogenetic trees with bootstraps of 100.

{kind=link}
{kind=link}
| Primers | Oligonucleotide Sequence (5′–3′) |
|---|---|
| adk | F-GGG CTG GAA AAG GTA CAC AA |
| R-ACG CAA GCT CCT TTT GAA TC | |
| icdA | F-GGG ACG AGA TGA CCA GGA T |
| R-TTT TTT GAG ATC CGC AGC TTT | |
| lipL32 | F-ATC TCC GTT GCA CTC TTT GC |
| R-ACC ATC ATC ATC ATC GTC CA | |
| lipL41 | F-TAG GAA ATT GCG CAG CTA CA |
| R-GCA TCG AGA GGA ATT AAC ATC A | |
| rrs2 | F-CAT GCA AGT CAA GCG GAG TA |
| R-AGT TGA GCC CGC AGT TTT C | |
| secY | F-ATG CCG ATC ATT TTT GCT TC |
| R-CCG TCC CTT AAT TTT AGA CTT CTT C |
| Group No. | Mean Distance (within Groups) | Standard Error (S.E) |
|---|---|---|
| 1 | 0.0028 | 0.0004 |
| 2 | 0.0007 | 0.0005 |
| 3 | 0.0007 | 0.0005 |
| 4 | 0.0017 | 0.0008 |
| 5 | 0.0000 | 0.0000 |
| 6 | 0.0000 | 0.0000 |
| 7 | 0.0181 | 0.0024 |
| 8 | 0.0049 | 0.0011 |
| Group No. | Mean Distance (within Groups) | Standard Error (S.E) |
|---|---|---|
| 1 | 0.000 | 0.000 |
| 2 | 0.001 | 0.001 |
| 3 | 0.002 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amran, F.; Noor Halim, N.A.; Muhammad, A.H.; Mohd Khalid, M.K.N.; Dasiman, N.M.; Shamsusah, N.A.; Abd Talib, A.K.A.; Noh, M.A.; Mohd Ali, M.R.; Hashim, R. Application of Multilocus Sequence Typing for the Characterization of Leptospira Strains in Malaysia. Trop. Med. Infect. Dis. 2023, 8, 69. https://doi.org/10.3390/tropicalmed8020069
Amran F, Noor Halim NA, Muhammad AH, Mohd Khalid MKN, Dasiman NM, Shamsusah NA, Abd Talib AKA, Noh MA, Mohd Ali MR, Hashim R. Application of Multilocus Sequence Typing for the Characterization of Leptospira Strains in Malaysia. Tropical Medicine and Infectious Disease. 2023; 8(2):69. https://doi.org/10.3390/tropicalmed8020069
Chicago/Turabian StyleAmran, Fairuz, Nurul Atiqah Noor Halim, Ayu Haslin Muhammad, Mohd Khairul Nizam Mohd Khalid, Nur Mukmina Dasiman, Nadia Aqilla Shamsusah, Abdul Khalif Adha Abd Talib, Mohamed Asyraf Noh, Mohammad Ridhuan Mohd Ali, and Rohaidah Hashim. 2023. "Application of Multilocus Sequence Typing for the Characterization of Leptospira Strains in Malaysia" Tropical Medicine and Infectious Disease 8, no. 2: 69. https://doi.org/10.3390/tropicalmed8020069
APA StyleAmran, F., Noor Halim, N. A., Muhammad, A. H., Mohd Khalid, M. K. N., Dasiman, N. M., Shamsusah, N. A., Abd Talib, A. K. A., Noh, M. A., Mohd Ali, M. R., & Hashim, R. (2023). Application of Multilocus Sequence Typing for the Characterization of Leptospira Strains in Malaysia. Tropical Medicine and Infectious Disease, 8(2), 69. https://doi.org/10.3390/tropicalmed8020069