
Assessment of an Immuno-Diagnostic Method for Hookworm-Related Cutaneous Larva Migrans Using Crude Extracts of Ancylostoma caninum

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antigens
2.2. Serum Samples
2.3. ELISA
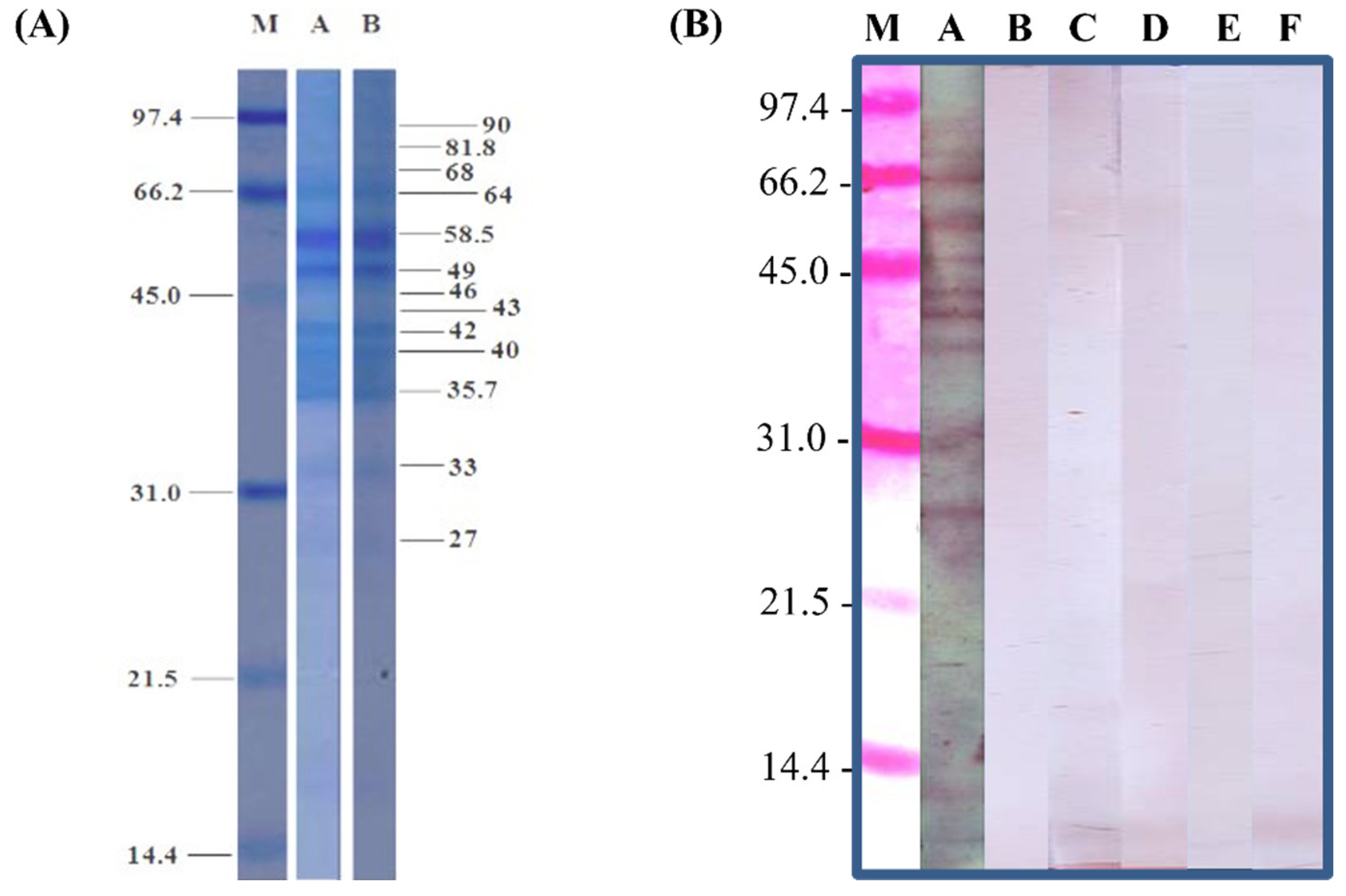
2.4. Observation of Crude Extract Profiles via Staining and Antibodies
2.5. Data Analysis
3. Results
3.1. Analysis of Crude Extract Profiles
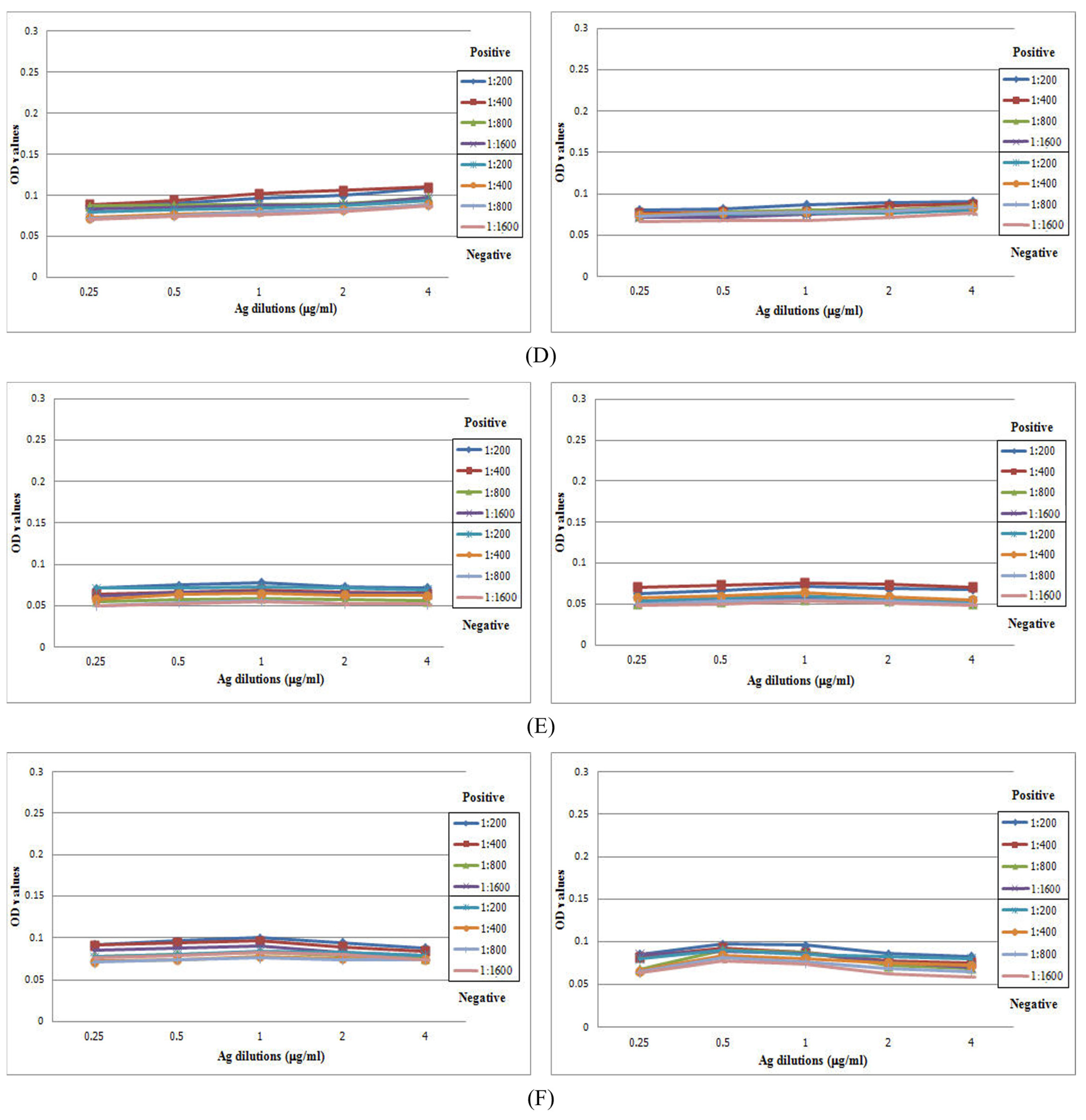
3.2. Correlations between Immunoglobulins and Antigens
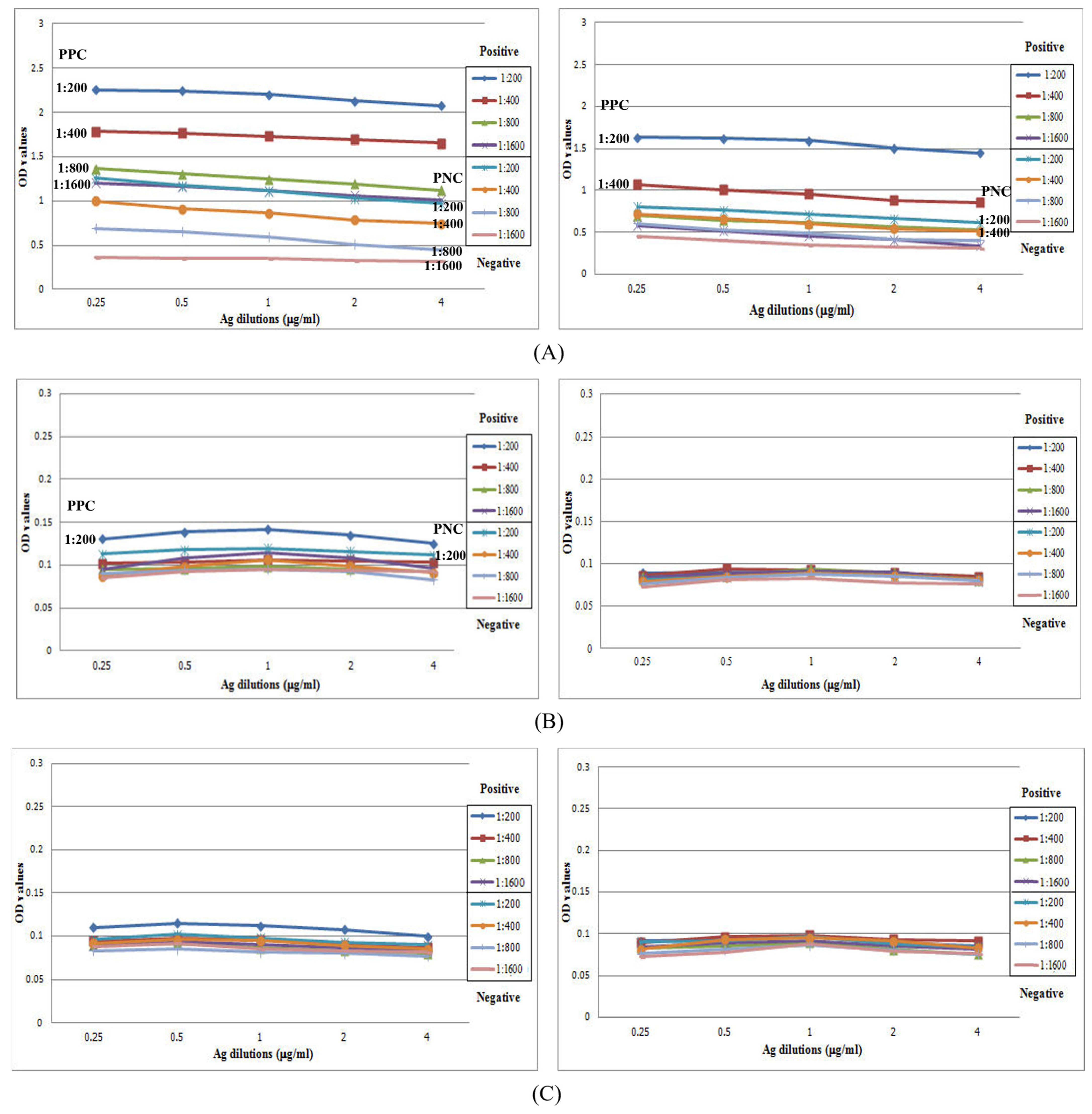
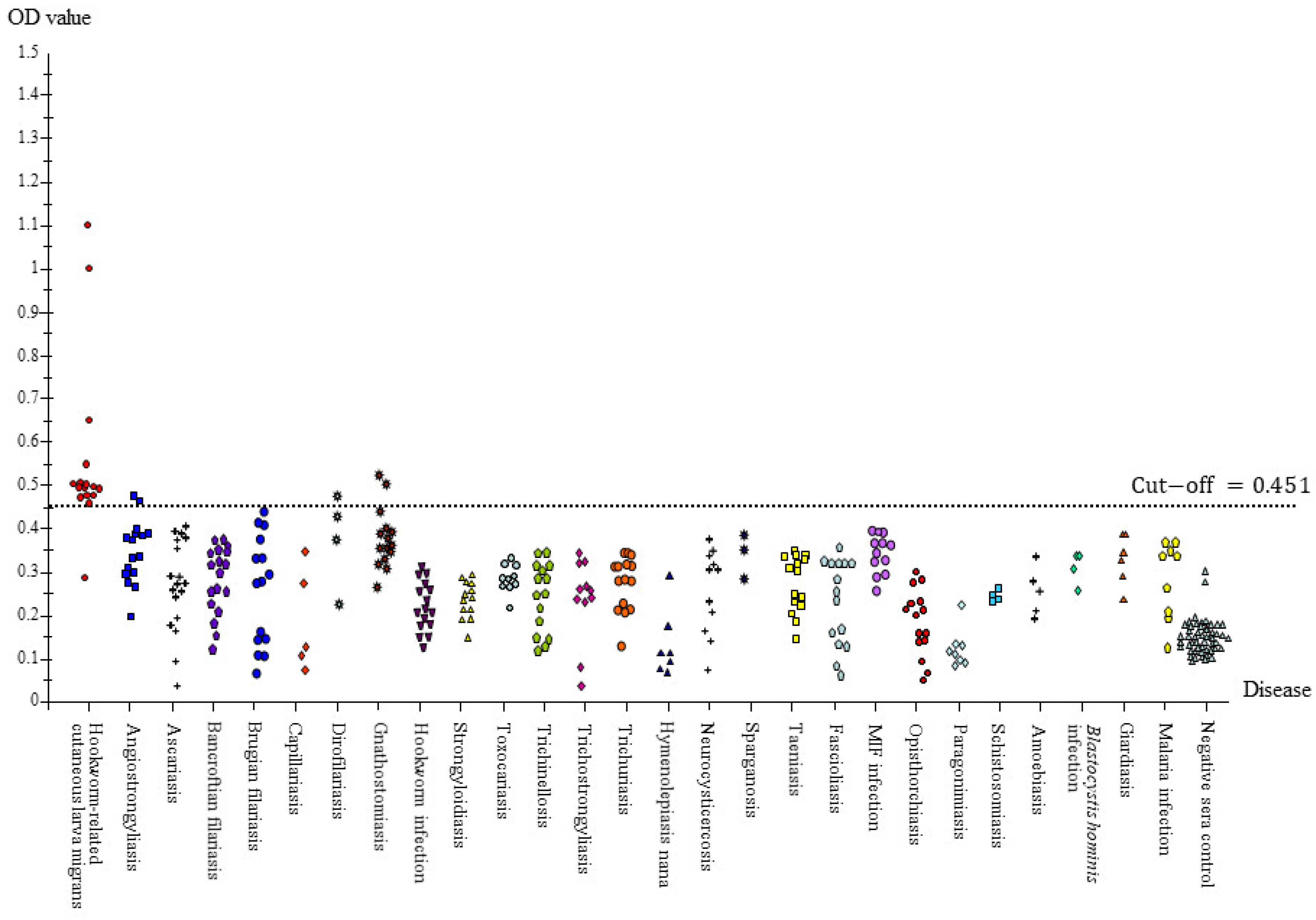
3.3. Full Scale IgG-ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antolová, D.; Miterpáková, M.; Paraličová, Z. Case of human Dirofilaria repens infection manifested by cutaneous larva migrans syndrome. Parasitol. Res. 2015, 114, 2969–2973. [Google Scholar] [CrossRef] [PubMed]
- Brenner, M.A.; Patel, M.B. Cutaneous larva migrans: The creeping eruption. Cutis 2003, 72, 111–115. [Google Scholar] [PubMed]
- Caumes, E. It’s time to distinguish the sign ‘creeping eruption’ from the syndrome ‘cutaneous larva migrans’. Dermatology 2006, 213, 179–181. [Google Scholar] [CrossRef]
- Hochedez, P.; Caumes, E. Hookworm-Related Cutaneous Larva Migrans. J. Trop. Med. 2007, 14, 326–333. [Google Scholar] [CrossRef]
- Xuan, L.T.; Hung, N.T.; Waikagul, J. Cutaneous fascioliasis: A case report in Vietnam. Am. J. Trop. Med. Hyg. 2005, 72, 508–509. [Google Scholar] [PubMed] [Green Version]
- Katz, M.; Despommier, D.D.; Gwadz, R. Parasitic Diseases; Springer: New York, NY, USA, 1988. [Google Scholar]
- Feldmeier, H.; Schuster, A. Mini review: Hookworm-related cutaneous larva migrans. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 915–918. [Google Scholar] [CrossRef]
- Anisuzzaman; Tsuji, N. Schistosomiasis and hookworm infection in humans: Disease burden, pathobiology and anthelmintic vaccines. Parasitol. Int. 2020, 75, 102051. [Google Scholar] [CrossRef]
- Malgor, R.; Oku, Y.; Gallardo, R.; Yarzábal, I. High prevalence of Ancylostoma spp. infection in dogs, associated with endemic focus of human cutaneous larva migrans, in Tacuarembo, Uruguay. Parasite 1996, 3, 131–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnaar, W.N.; Krecek, R.C.; Fourie, L.J. Helminths in dogs from a peri-urban resource-limited community in Free State Province, South Africa. Vet. Parasitol. 2002, 107, 343–349. [Google Scholar] [CrossRef]
- Wang, C.R.; Qiu, J.H.; Zhao, J.P.; Xu, L.M.; Yu, W.C.; Zhu, X.Q. Prevalence of helminthes in adult dogs in Heilongjiang Province, the People’s Republic of China. Parasitol. Res. 2006, 99, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, T.; Maiwald, H.; Nothdurft, H.D.; Löscher, T. Cutaneous larva migrans in travelers: Synopsis of histories, symptoms, and treatment of 98 patients. Clin. Infect. Dis. 1994, 19, 1062–1066. [Google Scholar] [CrossRef] [Green Version]
- Loukas, A.; Opdebeeck, J.; Croese, J.; Prociv, P. Immunologic incrimination of Ancylostoma caninum as a human enteric pathogen. Am. J. Trop. Med. Hyg. 1994, 50, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Morsy, H.; Mogensen, M.; Thomsen, J.; Thrane, L.; Andersen, P.E.; Jemec, G.B. Imaging of cutaneous larva migrans by optical coherence tomography. Travel. Med. Infect. Dis. 2007, 5, 243–246. [Google Scholar] [CrossRef]
- O’Connell, E.; Suarez, J.; Leguen, F.; Zhang, G.; Etienne, M.; Torrecilla, A.; Jimenez, A.; Farahi, F.; Alzugaray, M.; Rodriguez, D. Outbreak of cutaneous larva migrans at a children’s camp-Miami, Florida, 2006. MMWR Morb. Mortal. Wkly. Rep. 2007, 56, 1285–1287. [Google Scholar]
- Tremblay, A.; MacLean, J.D.; Gyorkos, T.; Macpherson, D.W. Outbreak of cutaneous larva migrans in a group of travellers. Trop. Med. Int. Health 2000, 5, 330–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, F.B. Creeping eruption. Int. J. Infect. Dis. 2010, 14, e545. [Google Scholar] [CrossRef] [Green Version]
- Hotez, P.J.; Narasimhan, S.; Haggerty, J.; Milstone, L.; Bhopale, V.; Schad, G.A.; Richards, F.F. Hyaluronidase from infective Ancylostoma hookworm larvae and its possible function as a virulence factor in tissue invasion and in cutaneous larva migrans. Infect. Immun. 1992, 60, 1018–1023. [Google Scholar] [PubMed]
- Muhleisen, J.P. Demonstration of pulmonary migration of the causative organism of creeping eruption. Ann. Intern. Med. 1953, 38, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.K.; Liu, T.T. Cutaneous larva migrans complicated by Löffler syndrome. Arch. Dermatol. 2010, 146, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Soo, J.K.; Vega-Lopez, F.; Stevens, H.P.; Chiodini, P.L. Cutaneous larvae migrans and beyond-a rare association. Travel. Med. Infect. Dis. 2003, 1, 41–43. [Google Scholar] [CrossRef]
- Burrill, D.Y.; Kotcher, E.; Childers, J.K. Nematode infestation of the buccal submucosa. Oral Surg. Oral Med. Oral Pathol. 1957, 10, 612–613. [Google Scholar] [CrossRef]
- Jaeger, R.G.; Araújo, V.C.D.; Marcucci, G.; Araújo, N.S.D. Larva migrans in the oral mucosa. J. Oral Med. 1987, 42, 246–247. [Google Scholar]
- André, J.; Bernard, M.; Ledoux, M.; Achten, G. Larva migrans of the Oral Mucosa. Dermatology 1988, 176, 296–298. [Google Scholar] [CrossRef]
- Horman, R. Larva migrans. In Textbook of Dermatology, 4th ed.; Blackwell: Oxford, UK, 1986; pp. 995–997. [Google Scholar]
- Lopes, M.A.; Zaia, A.A.; de Almeida, O.P.; Scully, C. Larva migrans that affect the mouth. Oral Surg. Oral Med. Oral Pathol. 1994, 77, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Bonne, C. Invasion of the Submucosa of the Human Small Intestine by Ancylostoma Braziliense. Am. J. Trop. Med. 1937, s1–s17, 587–594. [Google Scholar] [CrossRef]
- Gandullia, E.; Lignana, E.; Rabagliati, A.; Penna, R. Visceral larva migrans caused by Ancylostoma caninum. Minerva Pediatr. 1981, 33, 917–923. [Google Scholar] [PubMed]
- Little, M.; Halsey, N.A.; Cline, B.L.; Katz, S.P. Ancylostoma larva in a muscle fiber of man following cutaneous larva migrans. Am. J. Trop. Med. Hyg. 1983, 32, 1285–1288. [Google Scholar]
- Nadbath, R.P.; Lawlor, P.P. Nematode (Ancylostoma) in the cornea; a case report. Am. J. Ophthalmol. 1965, 59, 486–490. [Google Scholar] [CrossRef]
- Beaver, P.C. The nature of visceral larva migrans. J. Parasitol. 1969, 55, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Manalang, C. Studies on Ankylostomiasis in the Philippines. In Proceedings of the Far Eastern Association of Tropical Medicine Transactions Sixth Biennial Congress, Tokyo, Japan, 1925; Kyorinsha Medical Publishing Co.: Tokyo, Japan, 1926; Volume 1, pp. 351–368. [Google Scholar]
- Deane, M. Helminths eliminated from a group of Amazon residents after treatment with hexilresorcinol. Rev. Saude Publica 1950, 3, 443–464. [Google Scholar]
- Komma, M.; Queiroz, D.S.; Silva, A.; Jardim, C. Occurrence and prevalence of hookworms in 14 children in Goiânia. Rev. Goiana Med. 1969, 15, 169–174. [Google Scholar]
- Carrias, V.; de Vargas, P. Infection of the human intestine by Ancylostoma caninum Ercolani 1859. Rev. Soc. Bras. Med. Trop. 1985, 18, 57. [Google Scholar]
- Witenberg, G. Some unusual observations on helminthiasis in Israel. Harefuah 1951, 41, 178–180. [Google Scholar]
- Caumes, E.; Ly, F.; Bricaire, F. Cutaneous larva migrans with folliculitis: Report of seven cases and review of the literature. Br. J. Dermatol. 2002, 146, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Davies, H.D.; Sakuls, P.; Keystone, J.S. Creeping eruption. A review of clinical presentation and management of 60 cases presenting to a tropical disease unit. Arch. Dermatol. 1993, 129, 588–591. [Google Scholar] [CrossRef]
- Kwon, I.H.; Kim, H.S.; Lee, J.H.; Choi, M.H.; Chai, J.Y.; Nakamura-Uchiyama, F.; Nawa, Y.; Cho, K.H. A serologically diagnosed human case of cutaneous larva migrans caused by Ancylostoma caninum. Korean J. Parasitol. 2003, 41, 233–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loukas, A.; Croese, J.; Opdebeeck, J.; Prociv, P. Detection of antibodies to secretions of Ancylostoma caninum in human eosinophilic enteritis. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 650–653. [Google Scholar] [CrossRef]
- Loukas, A.; Opdebeeck, J.; Croese, J.; Prociv, P. Immunoglobulin G subclass antibodies against excretory/secretory antigens of Ancylostoma caninum in human enteric infections. Am. J. Trop. Med. Hyg. 1996, 54, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Croese, J.; Loukas, A.; Opdebeeck, J.; Prociv, P. Occult enteric infection by Ancylostoma caninum: A previously unrecognized zoonosis. Gastroenterology 1994, 106, 3–12. [Google Scholar] [CrossRef]
- Haichou, X.; Yan, W.; Shuhua, X.; Sen, L.; Yong, W.; Guangjin, S.; Weituo, W.; Bin, Z.; Drake, L.; Zheng, F.; et al. Epidemiology of human ancylostomiasis among rural villagers in Nanlin County (Zhongzhou Village), Anhui Province, China: II. Seroepidemiological studies of the age relationships of serum antibody levels and infection status. Southeast Asian J. Trop. Med. Public Health 2000, 31, 736–741. [Google Scholar]
- Pothong, K.; Komalamisra, C.; Kalambaheti, T.; Watthanakulpanich, D.; Yoshino, T.P.; Dekumyoy, P. ELISA based on a recombinant Paragonimus heterotremus protein for serodiagnosis of human paragonimiasis in Thailand. Parasit Vectors 2018, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Ieamsuwan, I.; Watthanakulpanich, D.; Chaisri, U.; Adisakwattana, P.; Dekumyoy, P. Evaluation of immunodiagnostic tests for human gnathostomiasis using different antigen preparations of Gnathostoma spinigerum larvae against IgE, IgM, IgG, IgG1-4 and IgG1 patterns of post-treated patients. Trop. Med. Int. Health 2021, 26, 1634–1644. [Google Scholar] [CrossRef]
- Yoonuan, T.; Nuamtanong, S.; Dekumyoy, P.; Phuphisut, O.; Adisakwattana, P. Molecular and immunological characterization of cathepsin L-like cysteine protease of Paragonimus pseudoheterotremus. Parasitol. Res. 2016, 115, 4457–4470. [Google Scholar]
- Deardorff, T.L.; Throm, R. Commercial blast-freezing of third-stage Anisakis simplex larvae encapsulated in salmon and rockfish. J. Parasitol. 1988, 74, 600–603. [Google Scholar] [PubMed]
- Shikama, K. Effect of Freezing and Thawing on the Stability of Double Helix of DNA. Nature 1965, 207, 529–530. [Google Scholar] [CrossRef]
- Dekumyoy, P.; Insun, D.; Waikagul, J.; Tanantaphruti, M.; Rongsriyam, Y.; Coochote, W. IgG- and IgG4-detected antigens of Dirofilaria immitis adult worms for bancroftian filariasis by enzyme-linked immunoelectrotransfer blot. Southeast Asian J. Trop. Med. Public Health 2000, 31 (Suppl. S1), 58–64. [Google Scholar]
- Florkowski, C.M. Sensitivity, specificity, receiver-operating characteristic (ROC) curves and likelihood ratios: Communicating the performance of diagnostic tests. Clin. Biochem. Rev. 2008, 29 (Suppl. S1), S83–S87. [Google Scholar] [PubMed]
- Alvarez, P.; Melgarejo, C.; Beltran, G.; Santos, R.; Ferrer, K.; Elescano, I.; Victorero, E.; Meza, B.; Ortiz, N.; León, A.; et al. Four Case Reports of Cutaneous Sparganosis From Peruvian Amazon. Am. J. Dermatopathol. 2022, 44, 510–514. [Google Scholar] [CrossRef]
- Dainichi, T.; Nakahara, T.; Moroi, Y.; Urabe, K.; Koga, T.; Tanaka, M.; Nawa, Y.; Furue, M. A case of cutaneous paragonimiasis with pleural effusion. Int. J. Dermatol. 2003, 42, 699–702. [Google Scholar] [CrossRef]
- Singh, T.S.; Devi Kh, R.; Singh, S.R.; Sugiyama, H. A case of cutaneous paragonimiasis presented with minimal pleuritis. Trop. Parasitol. 2012, 2, 142–144. [Google Scholar] [CrossRef]
- Catchpole, B.N.; Snow, D. Human ectopic fascioliasis. Lancet 1952, 2, 711–712. [Google Scholar] [CrossRef]
- Griffin, M.P.; Tompkins, K.J.; Ryan, M.T. Cutaneous sparganosis. Am. J. Dermatopathol. 1996, 18, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Dolpady, J.; Wabl, M.; Curotto de Lafaille, M.A.; Lafaille, J.J. Sequential class switching is required for the generation of high affinity IgE antibodies. J. Exp. Med. 2012, 209, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, T.B.; Bandhauer, M.E.; Bunker, A.M.; Roberts, W.L.; Hill, H.R. New childhood and adult reference intervals for total IgE. J. Allergy Clin. Immunol. 2014, 133, 589–591. [Google Scholar] [CrossRef]
- Neva, F.A.; Kaplan, A.P.; Pacheco, G.; Gray, L.; Danaraj, T.J. Tropical eosinophilia. A human model of parasitic immunopathology, with observations on serum IgE levels before and after treatment. J. Allergy Clin. Immunol. 1975, 55, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, B.M.; Bartlett, A.; Godfrey, R.C.; Turton, J.A.; Worms, M.J.; Yeates, R.A. Antibody responses in self-infections with Necator americanus. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 66–71. [Google Scholar] [CrossRef]
- Phosuk, I.; Intapan, P.M.; Thanchomnang, T.; Sanpool, O.; Janwan, P.; Laummaunwai, P.; Aamnart, W.; Morakote, N.; Maleewong, W. Molecular detection of Ancylostoma duodenale, Ancylostoma ceylanicum, and Necator americanus in humans in northeastern and southern Thailand. Korean J. Parasitol. 2013, 51, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.J.; Inpankaew, T.; Sutthikornchai, C.; Sukthana, Y.; Thompson, R.C. PCR-based coprodiagnostic tools reveal dogs as reservoirs of zoonotic ancylostomiasis caused by Ancylostoma ceylanicum in temple communities in Bangkok. Vet. Parasitol. 2008, 155, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Nuchprayoon, S.; Sanprasert, V.; Suntravat, M.; Kraivichian, K.; Saksirisampant, W.; Nuchprayoon, I. Study of specific IgG subclass antibodies for diagnosis of Gnathostoma spinigerum. Parasitol. Res. 2003, 91, 137–143. [Google Scholar] [CrossRef]
- Laummaunwai, P.; Sawanyawisuth, K.; Intapan, P.M.; Chotmongkol, V.; Wongkham, C.; Maleewong, W. Evaluation of human IgG class and subclass antibodies to a 24 kDa antigenic component of Gnathostoma spinigerum for the serodiagnosis of gnathostomiasis. Parasitol. Res. 2007, 101, 703–708. [Google Scholar] [CrossRef]
- Gill, N.; Somayaji, R.; Vaughan, S. Exploring Tropical Infections: A Focus on Cutaneous Larva Migrans. Adv. Skin. Wound Care 2020, 33, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Firmansyah, Y. New Approachment of Creeping Eruption Management. J. Dermatol. Res. Ther. 2020, 6, 1–5. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Number of Serum Samples | Specific or Proven Diagnosis |
|---|---|---|
| Homologous Sera | ||
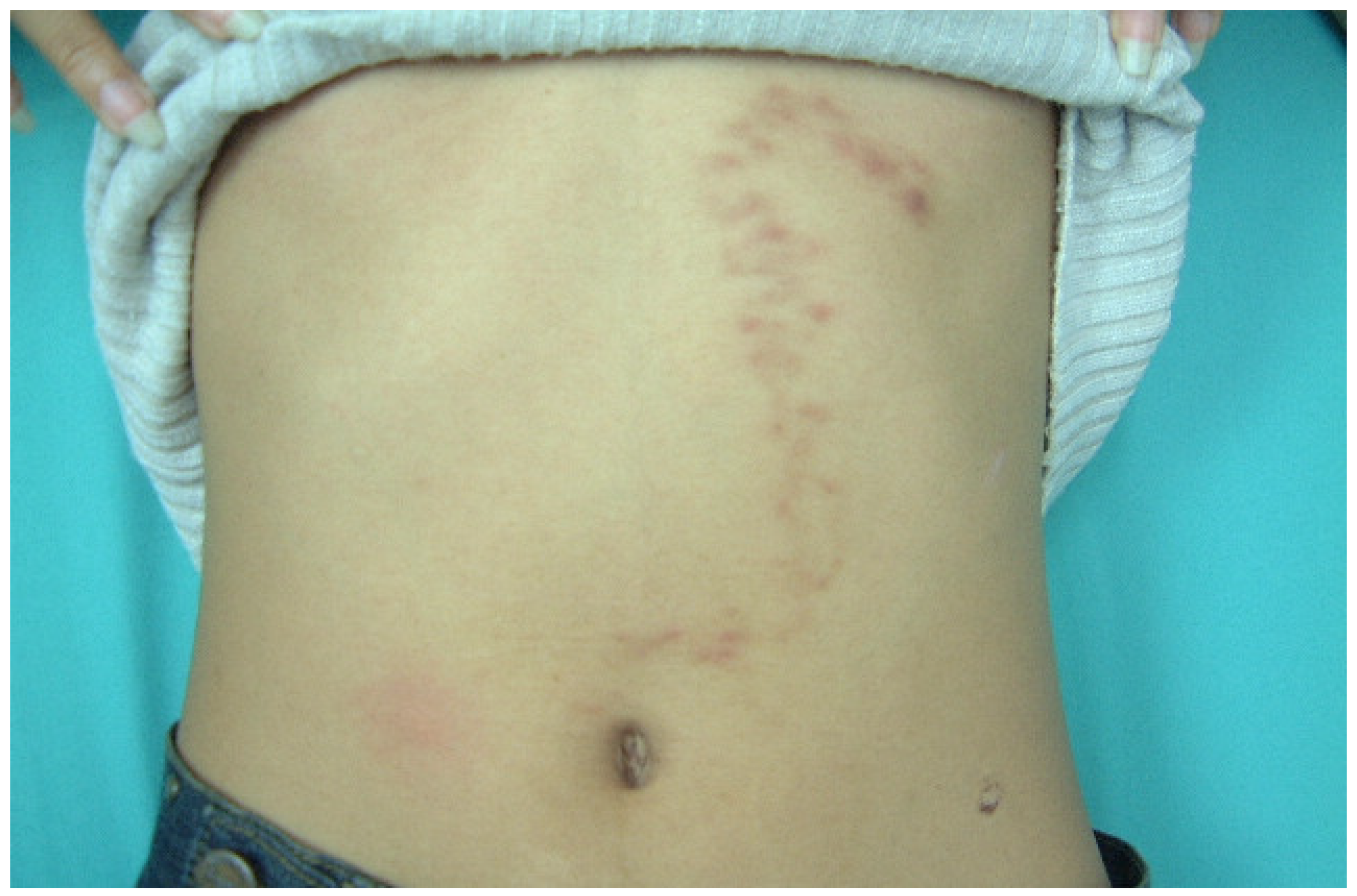
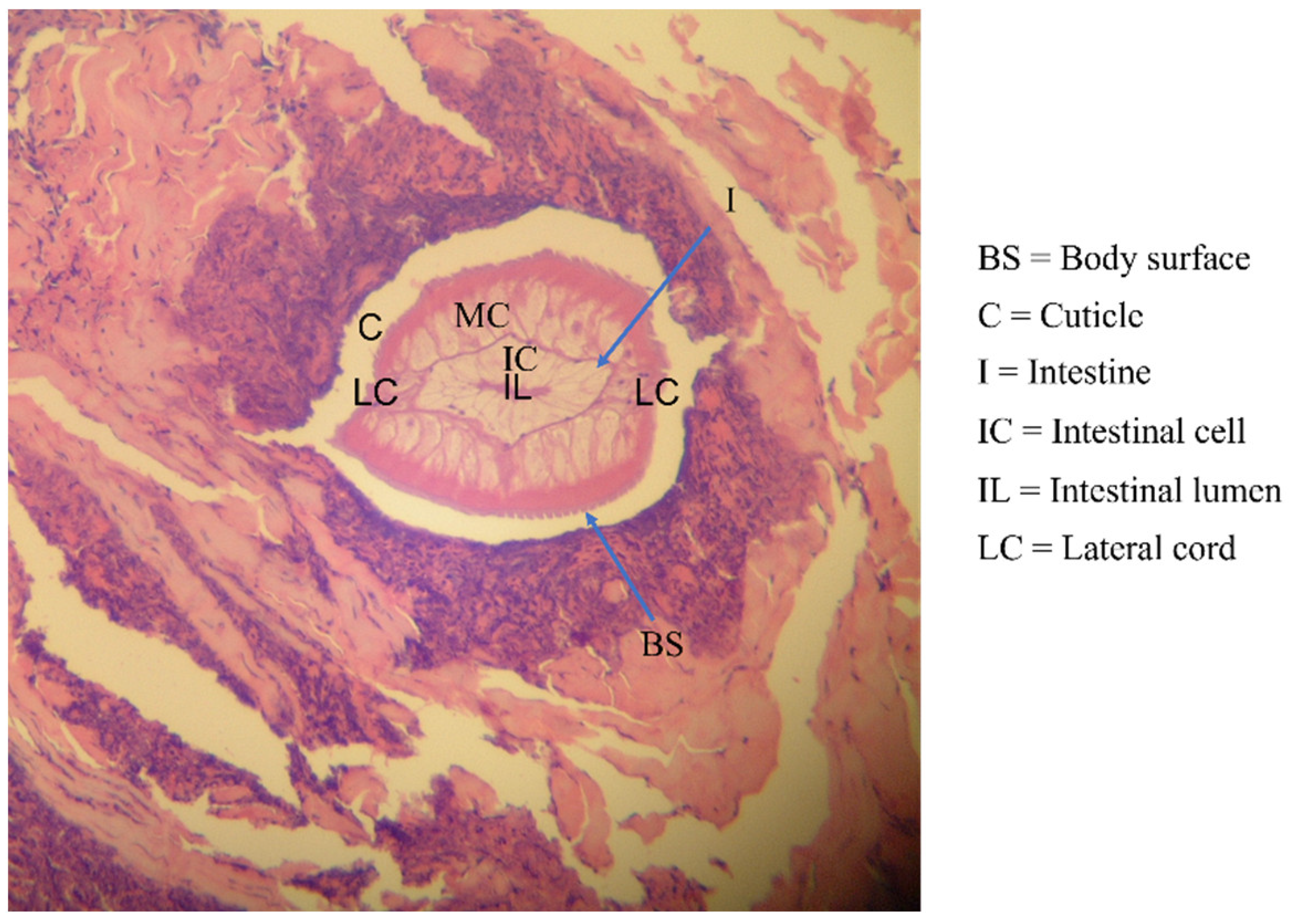
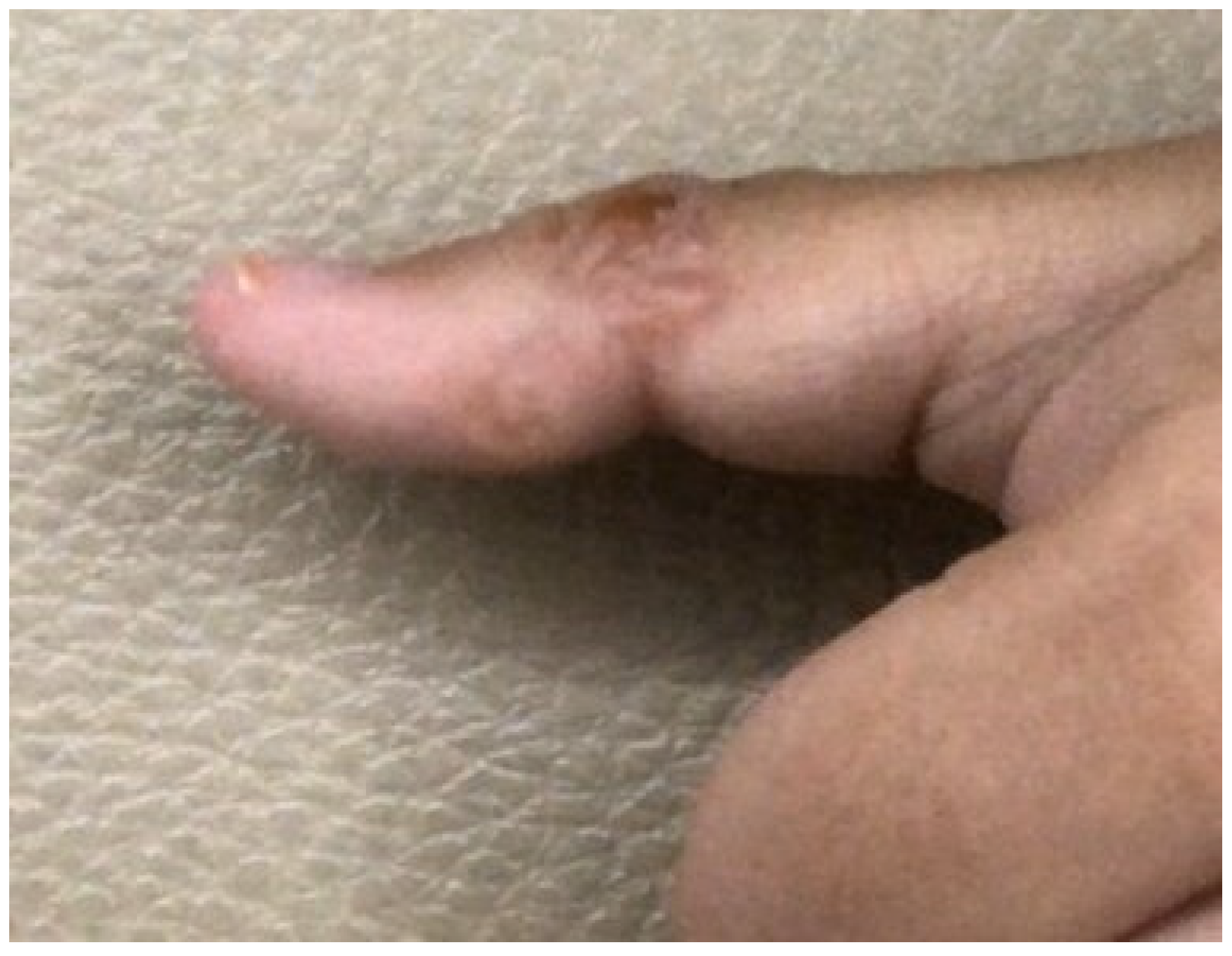
| Human-related cutaneous larva migrans (hwCLM) | 16 | Clinical manifestations related to cutaneous larva migrans or creeping eruption, such as pruritus, swelling pain, and the appearance of an erythematous track on the skin of the trunk, buttock, hand, leg, etc. History of traveling or/and exposure to soil or beach land where dogs were around Tested negative for gnathostomiasis with an IgG-immunoblotting test and for strongyloidiasis with an IgG-ELISA, including a demonstration of two hwCLM cases and a section of larva structure from the patient via tissue biopsy (Figure 1, Figure 2 and Figure 3) |
| Heterologous sera Nematodiasis | ||
| Angiostrongyliasis | 15 | Ocular angiostrongyliasis, larval stage found Larva in CSF sample found Consumption of snail intermediate hosts, stiff neck, and positive immunoblot |
| Ascariasis | 18 | Eggs in feces found by using the Kato thick smear technique |
| Bancroftian filariasis | 17 | Microfilariae on blood films |
| Brugian filariasis | 15 | Microfilariae on blood films and positive ELISA results |
| Capillariasis | 5 | Eggs and adults in feces found by using the simple smear technique and simple sedimentation |
| Dirofilaria infection | 4 | Sectioned worms in lung tissues and worms from the eyes and/or positive immunoblot results |
| Gnathostomiasis | 15 | Detection of worms and positive immunoblot results |
| Hookworm infection | 15 | Eggs in feces found by using the Kato thick smear technique or detection of L3 by using the polyethylene tube culture method |
| Strongyloidiasis | 11 | Larvae found in polyethylene tube culture |
| Toxocariasis | 10 | Clinical symptoms and positive immunoblot results |
| Trichinellosis | 15 | Biopsy and/or history of eating meat of wild pigs, clinical symptom manifestation, and positive immunoblot results |
| Trichostrongyliasis | 11 | Culture technique and larvae confirmation |
| Trichuriasis | 15 | Eggs found by using the Kato thick smear technique |
| Cestodiasis | ||
| Cystic echinocossosis | 6 | Scolices found or positive immunoblot results; 1 indigenous case and 5 imported cases |
| Hymenolepiasis nana | 8 | Eggs in feces found by using the simple smear technique |
| Neurocysticercosis | 11 | Biopsy or clinical manifestation accompanying a computerized axial tomographic (CAT) scan of the brain and positive immunoblot results |
| Sparganosis | 3 | Spargana found |
| Taeniasis | 15 | Taenia saginata segments or Taenia eggs found in feces |
| Trematodiasis | ||
| Fascioliasis | 3 | Eggs found or positive immunoblot results; 1 indigenous case and 2 cases supported by US Naval Medical Research Unit No. 3, Egypt |
| Opisthorchiasis | 15 | Worms detected post-treatment |
| Paragonimiasis heterotremus | 15 | Eggs in feces and sputum found by using the simple smear technique |
| Schistosomiasis | 4 | Schistosoma mansoni eggs found; cases supported by US Naval Medical Research Unit No.3, Egypt |
| Small intestinal fluke infection | 10 | Worms detected post-treatment |
| Protozoa infections | ||
| Amoebiasis | 5 | Cysts in feces found by using the simple smear technique |
| Blastocystis hominis infection | 4 | Cysts in feces found by using the simple smear technique |
| Giardiasis | 6 | Giardia intestinalis cysts in feces found by using the simple smear technique |
| Malaria | 9 | Plasmodium falciparum or P. vivax found in blood smears |
| Negative control sera | 30 | Negative results from stool examinations using the simple smear and formalin-ether concentration techniques for other parasitic infections. Negative screening results from IgG-ELISA using ten antigens, namely crude antigens of Gnathostoma spinigerum larvae, Strongyloides stercolaris larvae, Toxocara canis male and female adult worms, Angiostrongylus cantonensis adult worms, Dirofilaria immitis female adult worms, Trichinella spiralis muscle larvae, Taenia solium metacestode, Paragonimus heterotremus worms, Fasciola gigantica worms, and fluid of hydatid cyst [44,45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adam, S.; Dekumyoy, P.; Nacapunchai, D.; Ketboonlue, T.; Charunwatthana, P.; Dhitavat, J.; Koompapong, K.; Chonsawat, P.; Watthanakulpanich, D. Assessment of an Immuno-Diagnostic Method for Hookworm-Related Cutaneous Larva Migrans Using Crude Extracts of Ancylostoma caninum. Trop. Med. Infect. Dis. 2023, 8, 209. https://doi.org/10.3390/tropicalmed8040209
Adam S, Dekumyoy P, Nacapunchai D, Ketboonlue T, Charunwatthana P, Dhitavat J, Koompapong K, Chonsawat P, Watthanakulpanich D. Assessment of an Immuno-Diagnostic Method for Hookworm-Related Cutaneous Larva Migrans Using Crude Extracts of Ancylostoma caninum. Tropical Medicine and Infectious Disease. 2023; 8(4):209. https://doi.org/10.3390/tropicalmed8040209
Chicago/Turabian StyleAdam, Sitthithana, Paron Dekumyoy, Duangporn Nacapunchai, Thawatchai Ketboonlue, Prakaykaew Charunwatthana, Jittima Dhitavat, Khuanchai Koompapong, Putza Chonsawat, and Dorn Watthanakulpanich. 2023. "Assessment of an Immuno-Diagnostic Method for Hookworm-Related Cutaneous Larva Migrans Using Crude Extracts of Ancylostoma caninum" Tropical Medicine and Infectious Disease 8, no. 4: 209. https://doi.org/10.3390/tropicalmed8040209
APA StyleAdam, S., Dekumyoy, P., Nacapunchai, D., Ketboonlue, T., Charunwatthana, P., Dhitavat, J., Koompapong, K., Chonsawat, P., & Watthanakulpanich, D. (2023). Assessment of an Immuno-Diagnostic Method for Hookworm-Related Cutaneous Larva Migrans Using Crude Extracts of Ancylostoma caninum. Tropical Medicine and Infectious Disease, 8(4), 209. https://doi.org/10.3390/tropicalmed8040209