


Geographic Distribution of the Genus Panstrongylus Berg, 1879 in the Neotropic with Emphasis on Trypanosoma cruzi Vectors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Species Database and Georeferencing
2.3. Modelling the Geographical Distribution, Diversity, and Species Richness of Panstrongylus and Sympatric Mammals
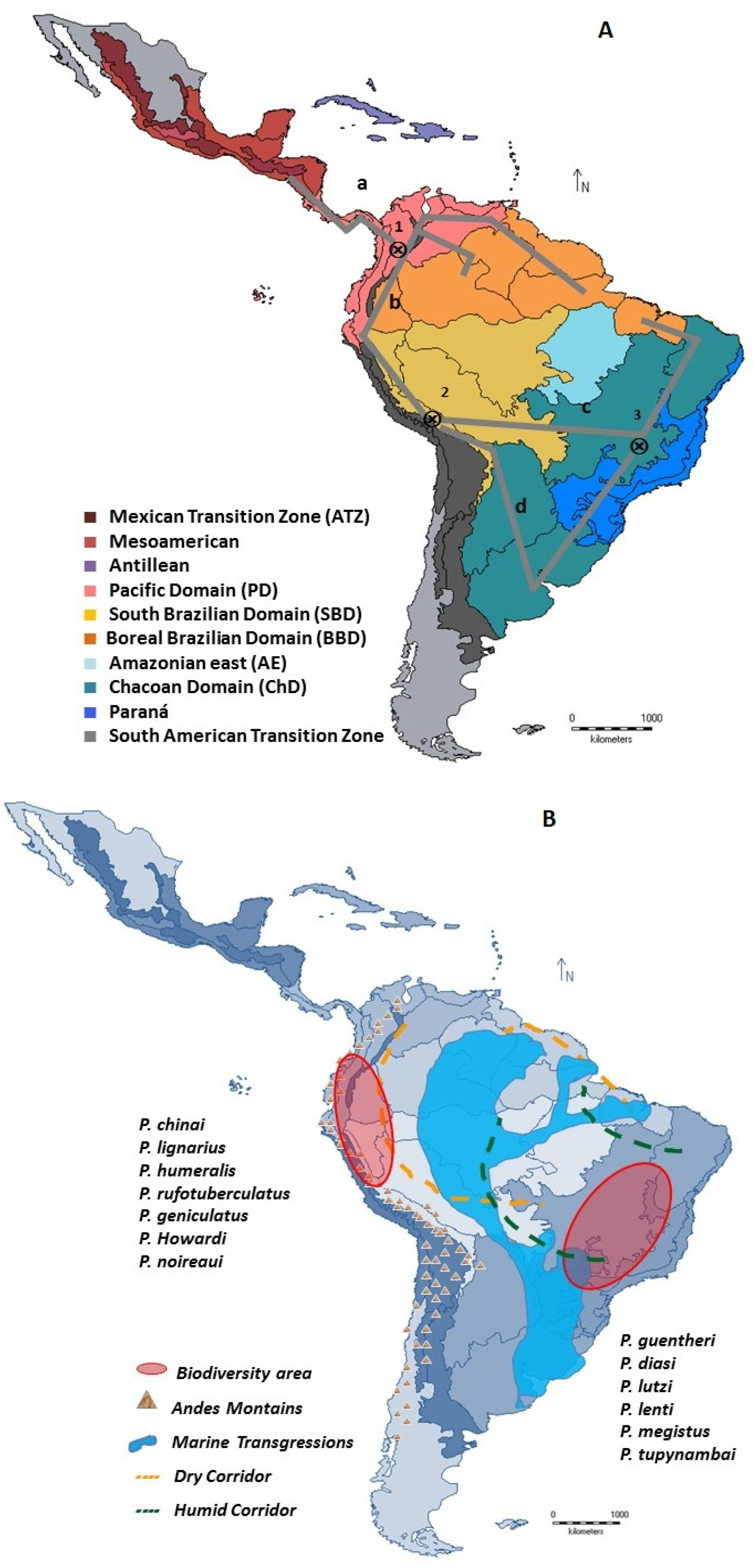
2.4. Pan-Biogeographical Analysis
3. Results and Discussion
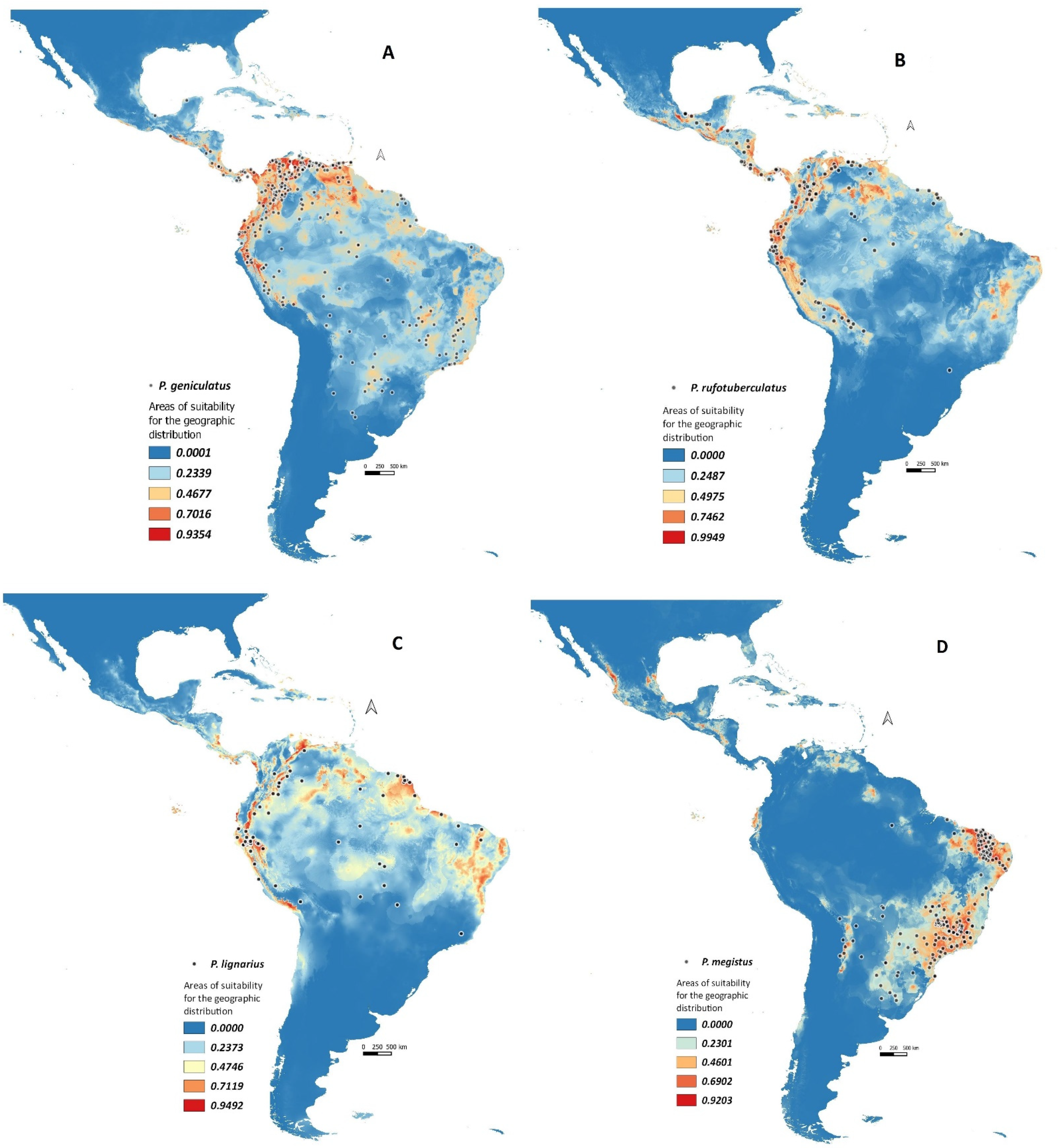
3.1. Geographical Distribution Analysis of Panstrongylus Species as Frequent or Sporadic Vectors of T. cruzi
3.1.1. Panstrongylus geniculatus
3.1.2. Panstrongylus rufotuberculatus
3.1.3. Panstrongylus lignarius
3.1.4. Panstrongylus megistus

3.1.5. Panstrongylus chinai
3.2. Individual and General Traces of the Distribution of Panstrongylus in the Neotropic
- Generalized trace a. Located in the SATZ (Veracruzana and Chiapas provinces; AMD, Mosquito province) and PD (Guatuso, Puntarenas-Chiriquí, Chocó-Darien, and Magdalena provinces), where P. geniculatus, P. humeralis, P. lignarius and P. rufotuberculatus were found.
- Generalized trace b. PD provinces (Venezuelan Guajira and Savanna—east of PD; Cauca, western Ecuador and Ecuadorian zone—west of PD); BBD provinces (Pantepui, Guyana Lowlands); Andean “Paramo” province of SATZ; BD (Napo and Imerí) and BSD (Ucayali, Rondonia, and Yungas) provinces, which include P. chinai, P. geniculatus, P. lignarius, P. howardi, P. martinezorum, and P. rufotuberculatus.
- Generalized trace c. Includes P. diasi, P. geniculatus, P. lignarius, P. lenti, P. lutzi, P. megistus, P. rufotuberculatus, P. noireaui from the BD (Rondônia) through provinces of the Chaco subregion (“Cerrado” and Parana) to areas of the BBD (Para), Chaco domain (ChD) (“Caatinga” and “Cerrado”), and Parana domain (Parana Forest).
- Generalized trace d. Widespread in the Chaco (Chaco and Pampeana departments) and Parana subregions (Parana Forest Department) with P. diasi, P. geniculatus, P. guentheri, P. lignarius, P. megistus, and P. tupynambai.
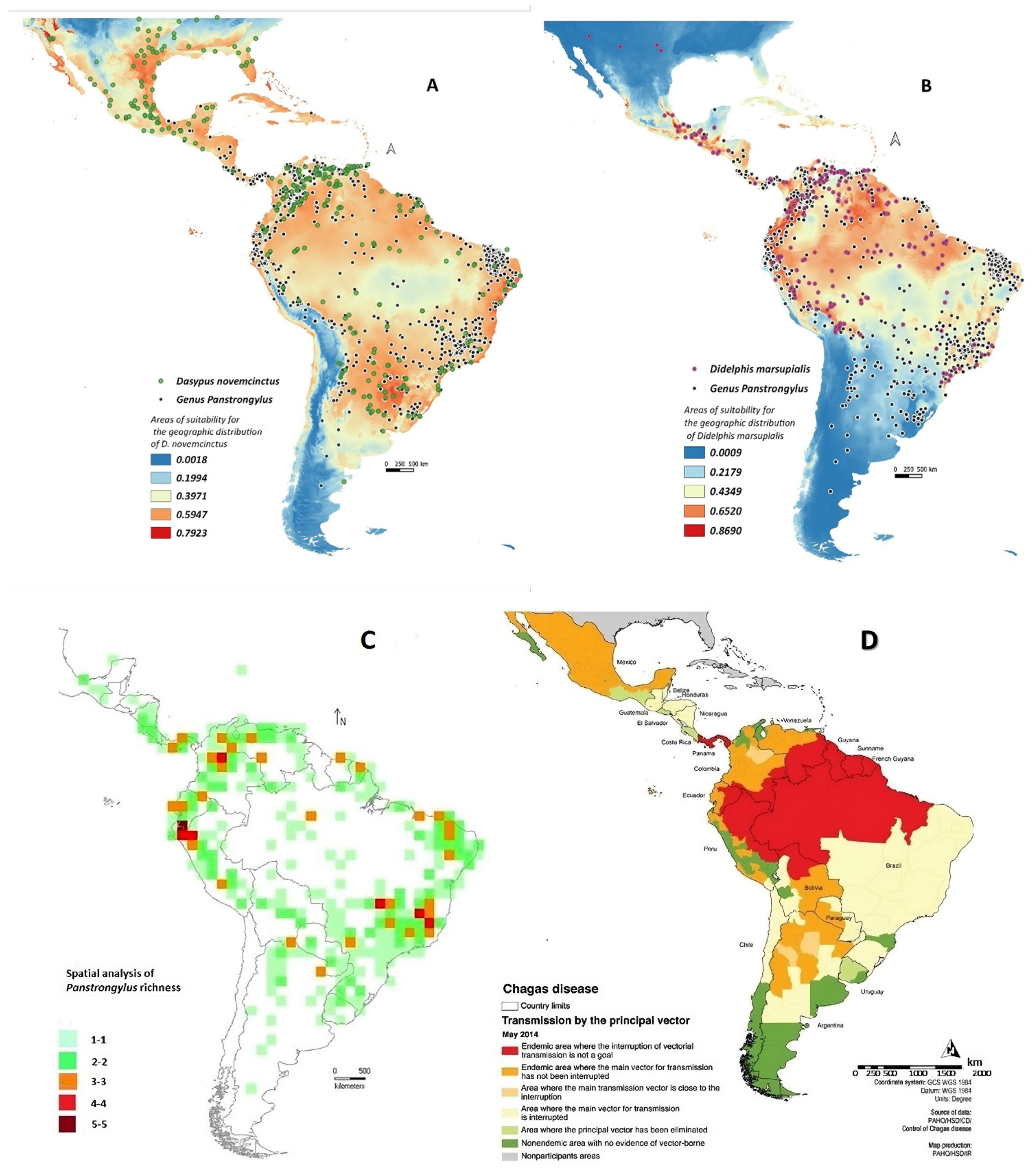
3.3. Species Richness of Panstrongylus, the Associated Mastofauna and the CD Cases in the Neotropical Region
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Otálora-Luna, F.; Pérez-Sánchez, A.J.; Sandoval, C.; Aldana, E. Evolution of hematophagous habit in Triatominae (Heteroptera: Reduviidae). Rev. Chil. Hist. Nat. 2015, 88, 4. [Google Scholar] [CrossRef]
- Ayala, J.M. Una nueva especie de Panstrongylus Berg de Venezuela (Hemiptera: Reduviidae, Triatominae). Entomotropica 2009, 24, 105–109. [Google Scholar]
- Poinar, G., Jr. Panstrongylus hispaniolae sp. n. (Hemiptera: Reduviidae: Triatominae), a new fossil triatomine in Dominican amber, with evidence of gut flagellates. Palaeodiversity 2013, 6, 1–8. [Google Scholar]
- Lent, H.; Wygodzinsky, P. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas disease. Bull. Am. Museum. Nat. Hist. 1979, 163, 123–520. [Google Scholar]
- Carcavallo, R.; Rodriguez, M.; Salvatella, R.; Curto De Casas, S.; Rocha, D.; Sherlock, I.; Galvão, C.; Rocha, D.; Galíndez, I.; Otero, M.; et al. Habitats and related Fauna. In Atlas of Chagas Disease Vectors in the Americas; Carcavallo, R., Galíndez, I., Jurberg, J., Lent, H., Eds.; Editora Fiocruz: Rio Janeiro, Brasil, 1999; Volume 2, pp. 537–560. [Google Scholar]
- Leite, G.R.; Dos Santos, C.B.; Falqueto, A. Insecta, Hemiptera, Reduviidae, Panstrongylus geniculatus: Geographic distribution map. Check List 2007, 3, 147–152. [Google Scholar] [CrossRef]
- Flores-Ferrer, A.; Marcou, O.; Waleckx, E.; Dumontiel, E.; Gourbière, S. Evolutionary ecology of Chagas disease; what do we know and what do we need? Evol. Appl. 2018, 11, 470–487. [Google Scholar] [CrossRef]
- Reyes-Lugo, M.; Rodriguez-Acosta, A. Domiciliation of the sylvatic Chagas disease vector Panstrongylus geniculatus Latreille, 1811 (Triatominae: Reduviidae) in Venezuela. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 508. [Google Scholar] [CrossRef] [PubMed]
- Briceño-León, R. La enfermedad de Chagas en las Américas: Una perspectiva de ecosalud. Cad. Saude Publica 2009, 25 (Suppl. 1), S71–S82. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Bello, Z.; Zavala-Jaspe, R.; Reyes-Lugo, M.; Colmenares, C.; Noya-Alarcón, O.; Noya, O.; Herrera, L.; Alarcón de Noya, B. Urban Trypanosoma cruzi Oral Transmission: From a Zoonotic Founder Focus to the Largest Microepidemic of Chagas Disease. SOJ Microbiol. Infect. Dis. 2016, 4, 1–9. [Google Scholar]
- Herrera, L. Una revisión sobre reservorios de Trypanosoma (Schizotrypanum) cruzi (Chagas, 1909), agente etiológico de la Enfermedad de Chagas. Bol. Mal. Salud Ambient. 2010, 50, 3–15. [Google Scholar]
- Martínez-Hernández, F.; Oria-Martínez, B.; Rendón-Franco, E.; Villalobos, G.; Muñoz-García, C.I. Trypanosoma cruzi, beyond the dogma of non-infection in birds. Infect. Genet. Evol. 2022, 99, 105239. [Google Scholar] [CrossRef]
- Bonilla, A. Origen y distribución de las biotas: La biogeografía como disciplina integradora. Bol. Acad. Ciens. Fis. Mat. Nat. 2012, LXXII, 55–72. [Google Scholar]
- Hijmans, R.J.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant. Genet. Resour. Newsl. 2001, 127, 15–19. [Google Scholar]
- Liria, J. Sistemas de información geográfica y análisis espaciales: Un método combinado para realizar estudios panbiogeográficos. Rev. Mex. Biodivers. 2008, 79, 281–284. [Google Scholar] [CrossRef]
- Liria, J.; Navarro, J. Modelo de nicho ecológico en Haemagogus Williston (Diptera: Culicidae), vectores del virus de la fiebre amarilla. Rev. Biomed. 2010, 21, 149–161. [Google Scholar]
- Galvão, C.; Carcavallo, R.; Rocha, D.D.S.; Jurberg, J. A checklist of the current valid species of the subfamily Triatominae Jeannel, 1919 (Hemiptera, Reduviidae) and their geographical distribution, with nomenclatural and taxonomic notes. Zootaxa 2003, 202, 1–36. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Mechanistic and correlative models of ecological niches. Eur. J. Ecol. 2015, 1, 28–38. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Morocoima, A.; Carrasco, H.J.; Boadas, J.; Chique, J.D.; Herrera, L.; Urdaneta-Morales, S. Trypanosoma cruzi III from armadillos (Dasypus novemcinctus novemcinctus) from Northeastern Venezuela and its biological behavior in murine model. Risk of emergency of Chagas disease. Exp. Parasitol. 2012, 132, 341–347. [Google Scholar] [CrossRef]
- Molinari, J.; Aldana, E.; Nassar, J.M. Panstrongylus geniculatus (Heteroptera: Reduviidae: Triatominae): Natural infection with Trypanosoma cruzi under cavernicolous conditions in Paraguaná Peninsula, Venezuela. J. Cave Karst Stud. 2007, 69, 285–287. [Google Scholar]
- Ruiz-Guevara, R.; Muñoz-Calderón, A.; Alarcón de Noya, B.; Redondo, C.; Pulido, L.; Beitia, Y.; Mauriello, L.; Rojas, D.; Muñoz, J.; Navarro, E.; et al. Familiar outbreak of Chagas disease by oral transmission in Yaguapita, Miranda State, Venezuela. Bol. Malariol. Salud Ambient. 2017, 57, 45–57. [Google Scholar]
- Manel, S.; Ceri Williams, H.; Ormerod, S.J. Evaluating presence-absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Plasencia-Vázquez, A.H.; Escalona-Segura, G.; Esparza-Olguín, L.G. Modelación de la distribución geográfica potencial de dos especies de psitácidos neotropicales utilizando variables climáticas y topográficas. Acta Zool. Mex. 2014, 30, 471–490. [Google Scholar] [CrossRef]
- Galvão, C.; Jurberg, J.; Carcavallo, R.U.; Segura, C.A.; Girón, I.G.; Curto De Casas, S.I. Geographical Distribution and Alti-latitudinal Dispersion of Some Genera and Species of the Tribe Triatomini Jeannel, 1919 (Hemiptera, Reduviidae, Triatominae). Mem. Inst. Oswaldo Cruz 1998, 93, 33–37. [Google Scholar] [CrossRef]
- Villacís, A.G.; Ocaña-Mayorga, S.; Lascano, M.S.; Yumiseva, C.A.; Baus, E.G.; Grijalva, M.J. Abundance, natural infection with trypanosomes, and food source of an endemic species of triatomine, Panstrongylus howardi (Neiva 1911), on the ecuadorian central coast. Am. J. Trop. Med. Hyg. 2015, 92, 187–192. [Google Scholar] [CrossRef]
- Plasencia Vázquez, A.H.; Escalona Segura, G. Área de distribución geográfica potencial de las especies de aves psitácidas de la Península de Yucatán, México. Rev. Biol. Trop. 2014, 62, 1509–1522. [Google Scholar] [CrossRef]
- Morrone, J.J. Biogeographical regionalisation of the neotropical region. Zootaxa 2014, 3782, 1–110. [Google Scholar] [CrossRef]
- Morrone, J.J.; Escalante, T.; Rodriguez-Tapia, G.; Carmona, A.; Arana, M.; Mercado-Gomez, J.D. Biogeographic regionalization of the Neotropical region: New map and shapefile. An. Acad. Bras. Cienc 2022, 94, e20211167. [Google Scholar] [CrossRef]
- Morrone, J.J. Hacia una biogeografía evolutiva. Rev. Chil. Hist. Nat. 2007, 80, 509–520. [Google Scholar] [CrossRef]
- Mariño-Pérez, R.; Brailovsky, H.; Morrone, J.J. Análisis panbiogeográfico de las especies mexicanas de Pselliopus Bergroth (Hemiptera: Heteroptera: Reduviidae: Harpactorinae). Acta Zool. Mex. 2007, 23, 77–88. [Google Scholar] [CrossRef]
- Cáceres, A.; Troyes, L.; González-Pérez, A.; Llontop, E.; Bonilla, C.; Murias, E.; Heredia, N.; Velásquez, C.; Yañez, C. Enfermedad de Chagas en la región nororiental del Perú. I. Triatominos (Hemiptera, Reduviidae) presentes en Cajamarca y Amazonas. Rev. Peru. Med. Exp. Salud Publica 2002, 19, 17–23. [Google Scholar]
- Náquira, C.; Cabrera, R. Breve reseña histórica de la enfermedad de Chagas, a cien años de su descubrimiento y situación actual en el Perú. Rev. Peru. Med. Exp. Salud Publica 2009, 26, 494–504. [Google Scholar]
- Gurgel-Gonçalves, R.; Galvão, C.; Costa, J.; Peterson, A.T. Geographic distribution of Chagas disease vectors in Brazil based on ecological niche modeling. J. Trop. Med. 2012, 2012, 705326. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, C.; Jurberg, J.; Galvão, C.; Da Silva, D.; Rodrigues, J. Estudo Morfometrico do genero Panstrongylus, Berg 1879 (Hemiptera, Reduviidae, Triatominae). Mem. Inst. Oswaldo Cruz 2003, 98, 939–944. [Google Scholar]
- Juberg, J.; Rodrigues, J.; Moreira, F.; Dale, C.; Cordeiro, I.; Lamas, V.; Galvão, C.; Rocha, D. Atlas Iconográfico dos Triatomíneos do Brasil (Vetores da Doença de Chagas); Fiocruz IOC: Rio de Janeiro, Brazil, 2014; 58p. [Google Scholar]
- Patterson, J.; Barbosa, S.; Feliciangeli, D. On the genus Panstrongylus Berg 1879: Evolution, ecology and epidemiological significance. Acta Trop. 2009, 110, 187–199. [Google Scholar] [CrossRef]
- Dias-Lima, A.; Menezes, D.; Sherlock, T.; Noireau, F. Wild Habitat and Related Fauna of Panstrongylus lutzi (Reduviidae, Triatominae). J. Med. Entomol. 2003, 40, 989–990. [Google Scholar] [CrossRef]
- Caranha, L.; Seixas, E.; Da Silva, D.; Jurberg, J.; Galvão, C. Estudo das fontes alimentares de Panstrongylus lutzi (Neiva & Pinto, 1923) (Hemiptera: Reduviidae: Triatominae) no Estado do Ceará. Rev. Soc. Bras. Med. Trop. 2006, 39, 347–351. [Google Scholar]
- Forattini, O. Biogeografia, origen e distribuicao da domiciliacao de Triatomineos no Brasil. Cad. Saúde Pública 1990, 14, 265–299. [Google Scholar] [CrossRef]
- Bérenger, J.M.; Blanchet, D. A new species of the genus Panstrongylus from French Guiana (Heteroptera; Reduviidae; Triatominae). Mem. Inst. Oswaldo Cruz 2007, 102, 733–736. [Google Scholar] [CrossRef]
- Marín, E.; Santillán, R.; Cuba, C.; Jurberg, J.; Galvão, C. Hallazgo de Panstrongylus rufotuberculatus (Champion, 1899) (Hemiptera, Reduviidae, Triatominae) en ambiente domiciliario en la Región Piura, Perú. Cad. Saude Publica 2007, 23, 2235–2238. [Google Scholar] [CrossRef] [PubMed]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. Zookeys 2022, 1104, 203–225. Available online: https://zookeys.pensoft.net/article/81879/ (accessed on 27 March 2023). [CrossRef]
- Curto de Casas, I.; Carcavallo, R.; Galindez Girón, I.; Burgos, J. Bioclimatic factors and zones of Life. In Atlas of Chagas Disease Vectors in the Americas; Carcavallo, R.U., Galindez, I., Jurberg, J., Lent, H., Eds.; Fiocruz: Rio de Janeiro, Brazil, 1998; Volume 3, pp. 793–838. [Google Scholar]
- Pereira, J.M.; de Almeida, P.S.; de Sousa, A.V.; de Paula, A.M.; Bomfim Machado, R.; Gurgel-Gonçalves, R. Climatic factors influencing triatomine occurrence in Central-West Brazil. Mem. Inst. Oswaldo Cruz 2013, 108, 335–341. [Google Scholar] [CrossRef]
- Tineo-González, E.; Contreras-Peña, Y.; Reyes-Lugo, M.; Morocoima, A.; Herrera, L. Modelo de distribución espacial de Panstrongylus geniculatus Latreille 1811 (Hemiptera: Reduviidae: Triatominae) vector del agente de la Enfermedad de Chagas en Venezuela. Rev. Cien. Fac. Ciencs. Vet. 2021, XXXI, 7–15. [Google Scholar]
- Depickère, S.; Durán, P.; López, R.; Chávez, T. Presence of intradomicile colonies of the triatomine bug Panstrongylus rufotuberculatus in Muñecas, La Paz, Bolivia. Acta Trop. 2011, 117, 97–100. [Google Scholar] [CrossRef]
- Coriano, H.; Navas, C.; De Sousa, L.; Ferrer, E.; Herrera, L. Panstrongylus rufotuberculatus (Hemiptera, Reduviidae, Triatominae) infectado con Trypanosoma cruzi en el estado Anzoátegui (Venezuela). Bol. Mal. Salud Ambient. 2012, 52, 135–138. [Google Scholar]
- da Silva Rocha, D.; Lobato, A.; Galvão, C. Panstrongylus rufotuberculatus (Champion, 1899) (Hemiptera, Reduviidae, Triatominae) in Rondônia, Brazil: A novel report. Rev. Soc. Bras. Med. Trop. 2021, 54, e0141-2021. [Google Scholar]
- Brazil, R.P.; da Silva, A.R.; Albarelli, A.; Vale, J.F. Distribuição e infecção de triatomíneos por Trypanosoma do tipo cruzi na Ilha de São Luís-Maranhão. Rev Soc. Bras. Med. Trop. 1985, 18, 257–260. [Google Scholar] [CrossRef]
- Cazorla-Perfetti, D.; Nieves-Blanco, E. Triatominos de Venezuela: Aspectos taxonómicos, biológicos, distribución geográfica e importancia médica. Av. Cardiol. 2010, 30, 347–369. [Google Scholar]
- Cavassin, F.B.; Klisiowicz, D.D.R.; Oliveira, L.G.R.; Kuehn, C.C.; Kopp, R.L.; Thomaz Soccol, V.; Da Rosa, J.A.; Luz, E.; Mas-Coma, S.; Bargués, M.D. Genetic variability and geographical diversity of the main Chagas’ disease vector Panstrongylus megistus (Hemiptera: Triatominae) in Brazil based on ribosomal DNA intergenic sequences. J. Med. Entomol. 2014, 51, 616–628. [Google Scholar] [CrossRef]
- Vendrami, D.P.; Ceretti, W., Jr.; Marrelli, M.T. Analysis of mitochondrial cytochrome B oxidase gene suggests passive dispersal of the Panstrongylus megistus population in Brazil. J. Vector Ecol. 2014, 39, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Abad-Franch, F.; Monteiro, F.A. Biogeography and evolution of Amazonian triatomines (Heteroptera: Reduviidae): Implications for Chagas disease surveillance in humid forest ecoregions. Mem. Inst. Oswaldo Cruz 2007, 102 (Suppl. I), 57–69. [Google Scholar] [CrossRef]
- Bayer, A.M.; Hunter, G.C.; Gilman, R.H.; Del Carpio, J.G.C.; Naquira, C.; Bern, C.; Levy, M.Z. Chagas disease, migration and community settlement patterns in Arequipa, Peru. PLoS Negl. Trop. Dis. 2009, 3, e567. [Google Scholar] [CrossRef] [PubMed]
- Grijalva, M.J.; Terán, D.; Dangles, O. Dynamics of Sylvatic Chagas Disease Vectors in Coastal Ecuador Is Driven by Changes in Land Cover. PLoS Negl. Trop. Dis. 2014, 8, e2960. [Google Scholar] [CrossRef] [PubMed]
- Morrone, J.J. The south american transition zone: Characterization and evolutionary relevance. Acta Ent. Chilena 2004, 28, 41–50. [Google Scholar]
- Hwang, W.S.; Weirauch, C. Evolutionary History of Assassin Bugs (Insecta: Hemiptera: Reduviidae): Insights from Divergence Dating and Ancestral State Reconstruction. PLoS ONE 2012, 7, e45523. [Google Scholar] [CrossRef]
- Delsuc, F.; Vizcaíno, S.F.; Douzery, E.J. Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: A relaxed molecular clock study within xenarthrans. BMC Evol. Biol. 2004, 4, 11–23. [Google Scholar] [CrossRef]
- Justi, S.A.; Galvão, C.; Schrago, C.G. Geological Changes of the Americas and their Influence on the Diversification of the Neotropical Kissing Bugs (Hemiptera: Reduviidae: Triatominae). PLoS Negl. Trop. Dis. 2016, 10, e0004527. [Google Scholar] [CrossRef]
- Morocoima, A.; De Sousa, L.; Herrera, L.; Rojas, L.; Villalobos, M.; Chique, J.; Rogny, B.; Bonoli, S. Simpatría de triatominos (Reduviidae) y escorpiones (Buthidae) en Cocos nucifera y Acrocomia aculeata (Aracaceae) de Anzoátegui, Venezuela. Bol. Malariol. y Salud Ambient. 2011, 51, 187–198. [Google Scholar]
- Abad-Franch, F.; Lima, M.M.; Sarquis, O.; Gurgel-Gonçalves, R.; Sánchez-Martín, M.; Calzada, J.; Saldaña, A.; Monteiro, F.A.; Palomeque, F.S.; Santos, W.S.; et al. On palms, bugs, and Chagas disease in the Americas. Acta Trop. 2015, 151, 126–141. [Google Scholar] [CrossRef]
- Abad-Franch, F.; Monteiro, F.A.; Pavan, M.G.; Patterson, J.S.; Bargues, M.D.; Zuriaga, M.Á.; Aguilar, M.; Beard, C.B.; Mas-Coma, S.; Miles, M.A. Under pressure: Phenotypic divergence and convergence associated with microhabitat adaptations in Triatominae. Parasites Vectors 2021, 14, 195. [Google Scholar] [CrossRef]
- Posadas, P.; Crisci, J.V.; Katinas, L. Spatial methodologies in historical biogeography of islands. In The Biology of Island Floras; Bramwell, D., Caujapé-Castells, J., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 37–56. [Google Scholar]
- Florentin, J.E.; Arana, M.D.; Salas, R.M. Análisis panbiogeográfico del género Galianthe subgénero Ebelia (Rubiaceae). Rodriguesia 2016, 67, 437–444. [Google Scholar] [CrossRef]
- Juárez-Barrera, F.; Espinosa, D.; Morrone, J.J.; Escalante, T.; Bueno-Hernández, A.A. La complejidad biótica de la Zona de Transición Mexicana y la evolución del pensamiento biogeográfico de Gonzalo Halffter. Rev. Mex. Biodivers. 2020, 91, e913402. [Google Scholar] [CrossRef]
- Acosta, N.; López, E. Reservorios mamíferos del Trypanosoma cruzi en Paraguay. Mem. Inst. Investig. Cienc. Salud 2013, 11, 90–96. [Google Scholar]
- Sawyer, C.F.; Brinkman, D.C.; Walker, V.D.; Covington, T.D.; Stienstraw, E.A. The zoogeomorphic characteristics of burrows and burrowing by nine-banded armadillos (Dasypus novemcinctus). Geomorphology 2012, 157–158, 122–130. [Google Scholar] [CrossRef]
- Ginette Pérez Lazo, J.; Mayor, P.; Lescano, G.A. Amazonian Reservoir Hosts of Trypanosoma cruzi. In Biology of Trypanosoma cruzi; De Souza, W., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Cantillo-Barraza, O.; Bedoya, S.C.; Xavier, S.C.C.; Zuluaga, S.; Salazar, B.; Vélez-Mira, A.; Carrillo, L.M.; Triana-Chávez, O. Trypanosoma cruzi infection in domestic and synanthropic mammals such as potential risk of sylvatic transmission in a rural area from north of Antioquia, Colombia. Parasite Epidemiol. Control 2020, 11, e00171. [Google Scholar] [CrossRef]
- Enriquez, G.F.; Bua, J.; Orozco, M.M.; Macchiaverna, N.P.; Otegui, J.A.A.; Argibay, H.D.; Fernández, M.D.P.; Gürtler, R.E.; Cardinal, M.V. Over-dispersed Trypanosoma cruzi parasite load in sylvatic and domestic mammals and humans from northeastern Argentina. Parasites Vectors 2022, 15, 37. [Google Scholar] [CrossRef]
- Pineda, V.J.; González, K.A.; Perea, M.; Rigg, C.; Calzada, J.E.; Chaves, L.F.; Vásquez, V.; Samudio, F.; Gottdenker, N.; Saldaña, A. Show more Surveillance and genotype characterization of zoonotic trypanosomatidae in Didelphis marsupialis in two endemic sites of rural Panama. Int. J. Parasitol. Parasites Wildl. 2022, 17, 20–25. [Google Scholar] [CrossRef]
- Jansen, A.M.; Xavier das Chagas, S.C.; Roque, A.L.R. Landmarks of the Knowledge and Trypanosoma cruzi Biology in the Wild Environment. Front. Cell Infect. Microbiol. 2020, 10, 10. [Google Scholar] [CrossRef]
- Wilson, G.P.; Ekdale, E.G.; Hoganson, J.W.; Calede, J.J.; Vander Linden, A. A large carnivorous mammal from the Late Cretaceous and the North American origin of marsupials. Nat. Commun. 2016, 7, 13734. [Google Scholar] [CrossRef]
- Herrera, L. Trypanosoma cruzi, the Causal Agent of Chagas Disease: Boundaries between Wild and Domestic Cycles in Venezuela. Front. Public Health 2014, 2, 259. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Species | Distribution and Habitat | References |
|---|---|---|
| Panstrongylus chinai (Del Pontei, 1929) * | High jungle in Perú, Ecuador, and localities in the Venezuelan Andes. Found in rocky places and collected in the peridomicile (chicken coops) and inside dwellings. | [4,32,33] |
| Panstrongylus diasi (Pinto y Lent, 1946) | Distributed in “Caatinga” (Brazil), characterized as a semi-arid, warm, and xerophytic area; Santa Cruz (Bolivia), with a warm tropical climate. | [4,17,34] |
| Panstrongylus geniculatus (Latreille, 1811) * | From Mexico and Central America to South America (excluding Chile). Found in caves of armadillos, rodents, reptiles (geckos), and bats; also associated with opossums and bird nests, hollow trunks, under the bark of trees, in bromeliads and among palm leaves (Attalaea, Acromia, and Copernitia). Eurythermic species adapted to dry and humid ecotypes such as the Amazon rainforest and xenomorphic grassland areas. | [4,5,6,8,17,35] |
| Panstrongylus guentheri (Berg, 1879) | Areas of thorny scrub and dry forest in the Chaco region. Found in armadillo and rodent caves; also associated with opossums. Can be found in the peridomicile associated with woodpiles and environments of goats and dogs. | [4,17,36] |
| Panstrongylus hispaniolae (†) (Pionar, 2013) | Species are described from a fossil contained in a piece of amber found in the La Toca mine, located in Puerto Plata, Dominican Republic. | [3] |
| Panstrongylus howardi (Neiva, 1911) | Equatorial rainforest environment (Manabí province). It has been associated with marsupials and rodent nests inserted in the Aiphanes eggersi palm. Found in dwellings and the peridomicile in microhabitats such as piles of bricks or wood coexisting with rodents. | [4,26] |
| Panstrongylus humeralis (Usinger, 1939) | Distributed in humid forest from Panama and Colombia. | [4,5] |
| Panstrongylus lenti (Galvão y Palma, 1968) | Habitat typical of “Cerrado”; shrubby, herbaceous, and dry vegetation in Goiás and Bahia (Brazil). | [17,36] |
| Panstrongylus lignarius (Walker, 1837) * | Described from tropical dry forest habitats in Brazil, Colombia, Ecuador, French Guiana, Perú, Suriname, and Venezuela. Found in trees, hollow trunks, and palm trees; it has been associated with toucan nests and mammals such as porcupines and spiny rats. In Perú, it is considered a frequent species in the home and peridomicile. | [17,37] |
| Panstrongylus lutzi (Neiva y Pinto, 1923) * | Endemic to northeastern Brazil (“Caatinga” area). It colonizes armadillo caves and has been found in dwellings associated with rodents and opossums. | [4,17,38,39,40] |
| Panstrongylus martinezorum (Ayala, 2009) | Described from a male, collected in Puerto Ayacucho (Amazonas, Venezuela) and other males and females located in collections whose records indicate urban and peri-urban areas of capture. | [2] |
| Panstrongylus megistus (Burmeister, 1835) * | Wide distribution in Brazil, with great potential for colonization of artificial environments. In the wild, it is associated with palm trees and animal burrows. The Atlantic forest seems to represent the center of the distribution, although the species is also distributed in humid areas of the “Caatinga” and “Cerrado”. | [17,34,37,41] |
| Panstrongylus mitarakensis (Bérenger y Blanchet, 2007) | Distribution in the Mitarakara Mountains, French Guiana. | [42] |
| Panstrongylus rufotuberculatus (Champion, 1899). | Most widespread after P. geniculatus in Central and South America. Forest habitat, found in palms, hollows and mammal shelters such as bats, monkeys, armadillos. Occasionally in the peridomicile in association with domestic animals. | [4,10,17,37,43] |
| Panstrongylus tupynambai (Lent, 1942) | Atlantic forest region of southern Brazil and Uruguay. Collected in humid rocky habitats, palms, trees, shelters of mammals; human dwellings, and in the peridomicile. | [4,17] |
| Panstrongylus noireaui sp. nov | The specimens were found in the locality of Ayata (Camata), in the department of La Paz, province of Ildefonso de las Muñecas, Bolivia. It is located in the Yunga ecoregion. | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tineo-González, E.; Fermín, R.; Bonilla-Rivero, A.; Herrera, L. Geographic Distribution of the Genus Panstrongylus Berg, 1879 in the Neotropic with Emphasis on Trypanosoma cruzi Vectors. Trop. Med. Infect. Dis. 2023, 8, 272. https://doi.org/10.3390/tropicalmed8050272
Tineo-González E, Fermín R, Bonilla-Rivero A, Herrera L. Geographic Distribution of the Genus Panstrongylus Berg, 1879 in the Neotropic with Emphasis on Trypanosoma cruzi Vectors. Tropical Medicine and Infectious Disease. 2023; 8(5):272. https://doi.org/10.3390/tropicalmed8050272
Chicago/Turabian StyleTineo-González, Evelyn, Rossy Fermín, Ana Bonilla-Rivero, and Leidi Herrera. 2023. "Geographic Distribution of the Genus Panstrongylus Berg, 1879 in the Neotropic with Emphasis on Trypanosoma cruzi Vectors" Tropical Medicine and Infectious Disease 8, no. 5: 272. https://doi.org/10.3390/tropicalmed8050272
APA StyleTineo-González, E., Fermín, R., Bonilla-Rivero, A., & Herrera, L. (2023). Geographic Distribution of the Genus Panstrongylus Berg, 1879 in the Neotropic with Emphasis on Trypanosoma cruzi Vectors. Tropical Medicine and Infectious Disease, 8(5), 272. https://doi.org/10.3390/tropicalmed8050272