
Effect of Temperature and Ionic Substitutions on the Tegumental Potentials of Protoscoleces of Echinococcus granulosus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Collection
2.2. Saline Solutions and In Vitro Incubation Procedures
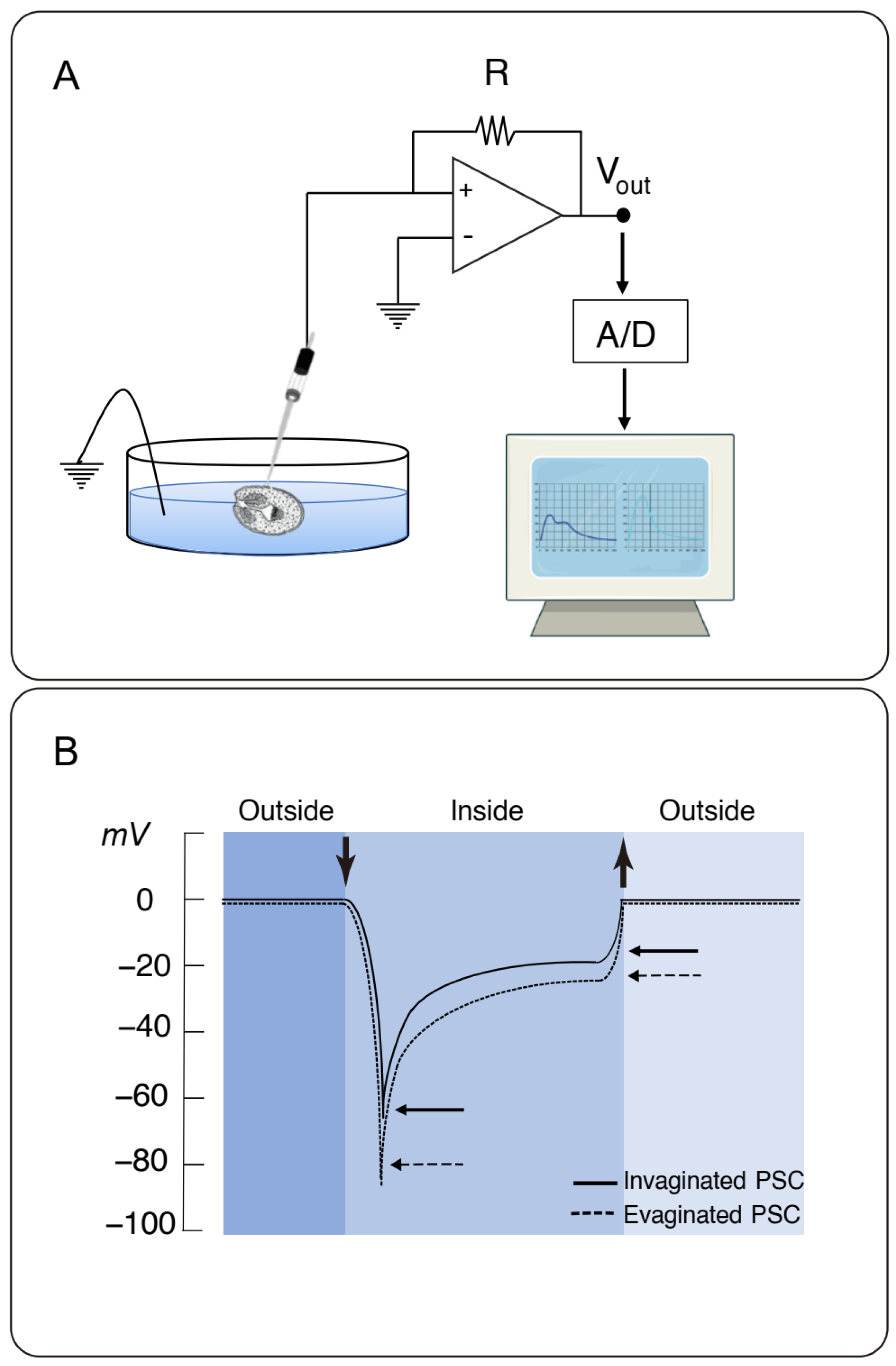
2.3. Microelectrode Measurements
2.4. Data Analyses and Statistics
3. Results
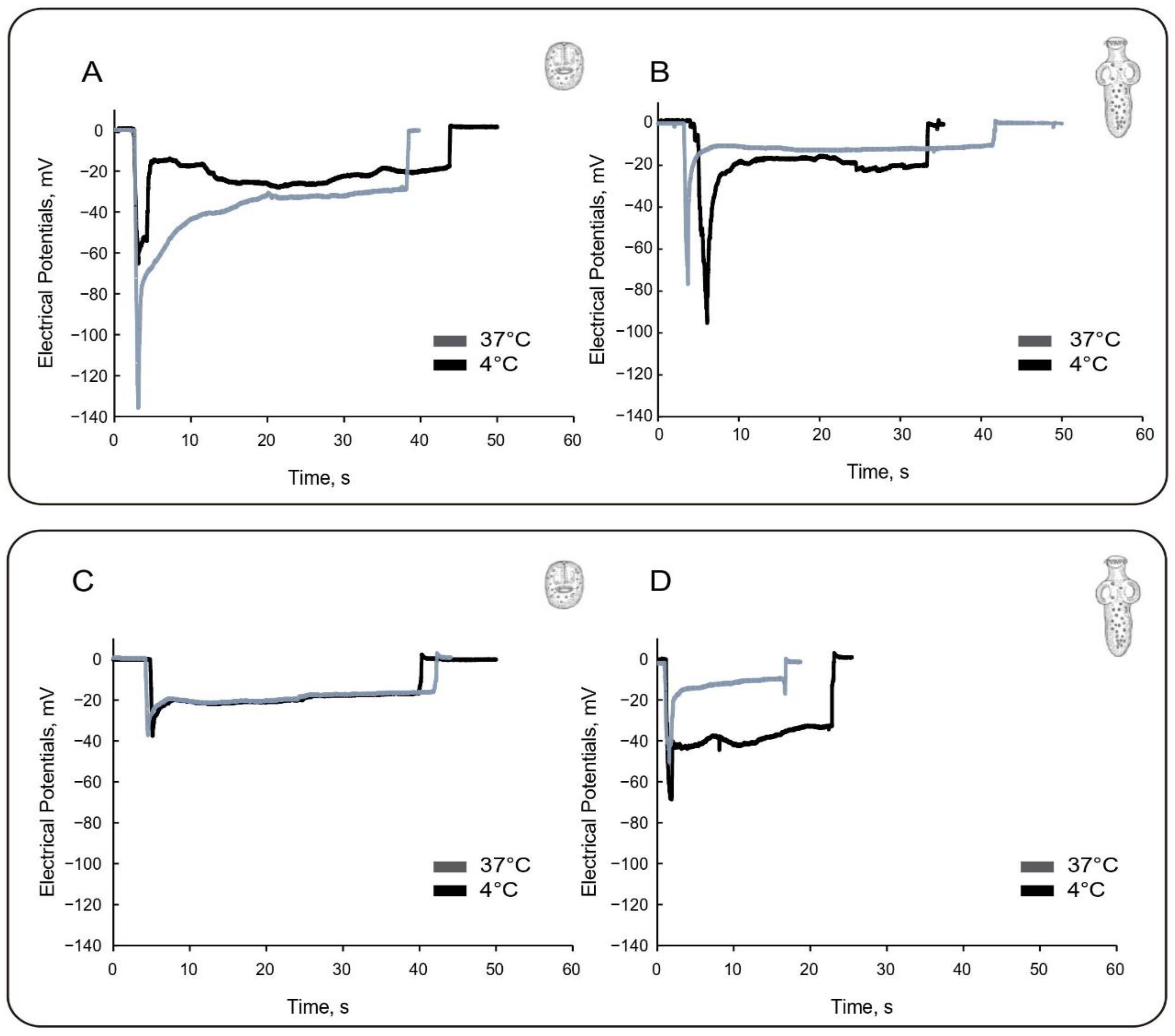
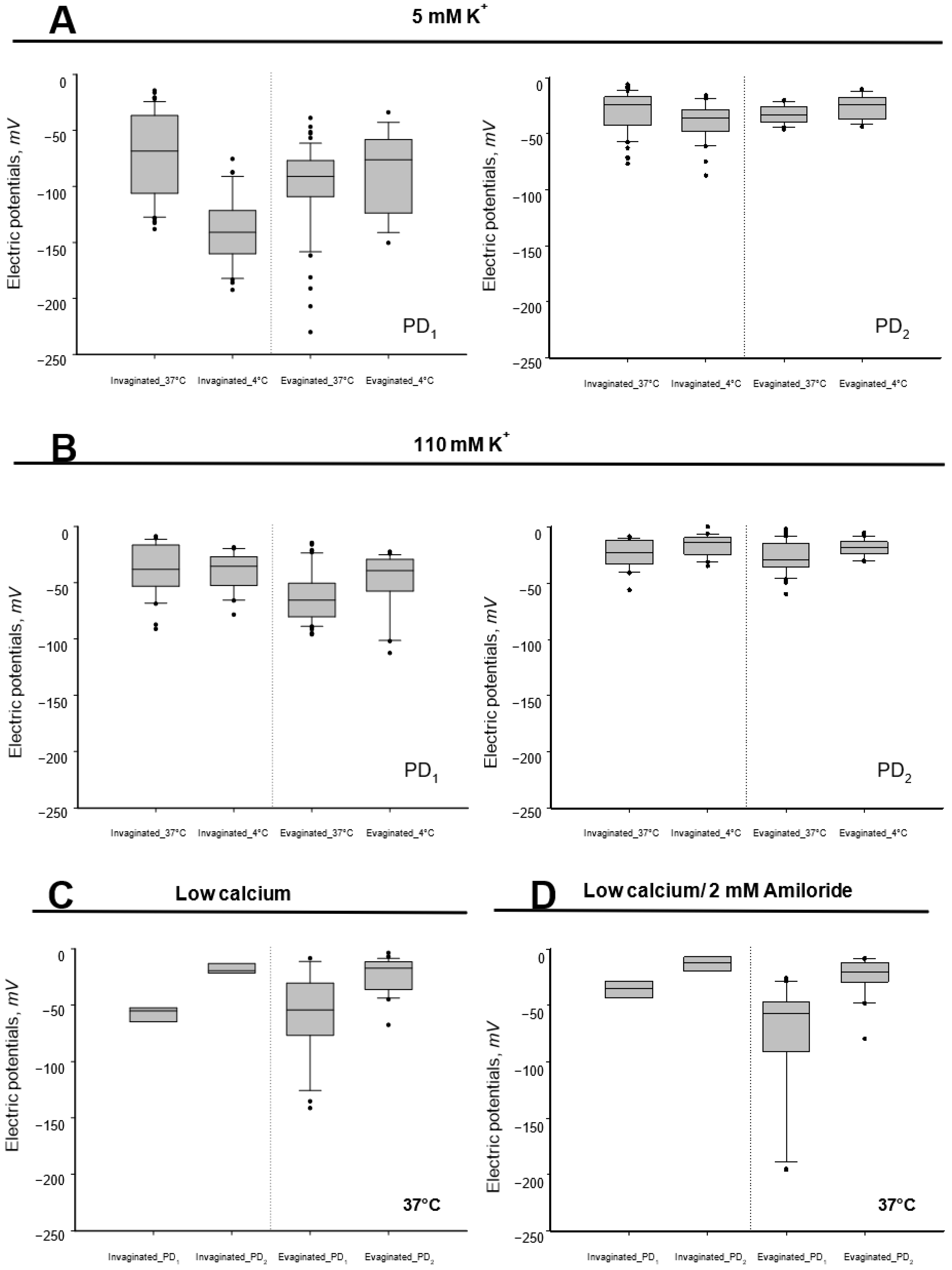
3.1. Effect of Low Temperature on Tegumental Membrane Potentials
3.2. Effect of High External Potassium Concentration
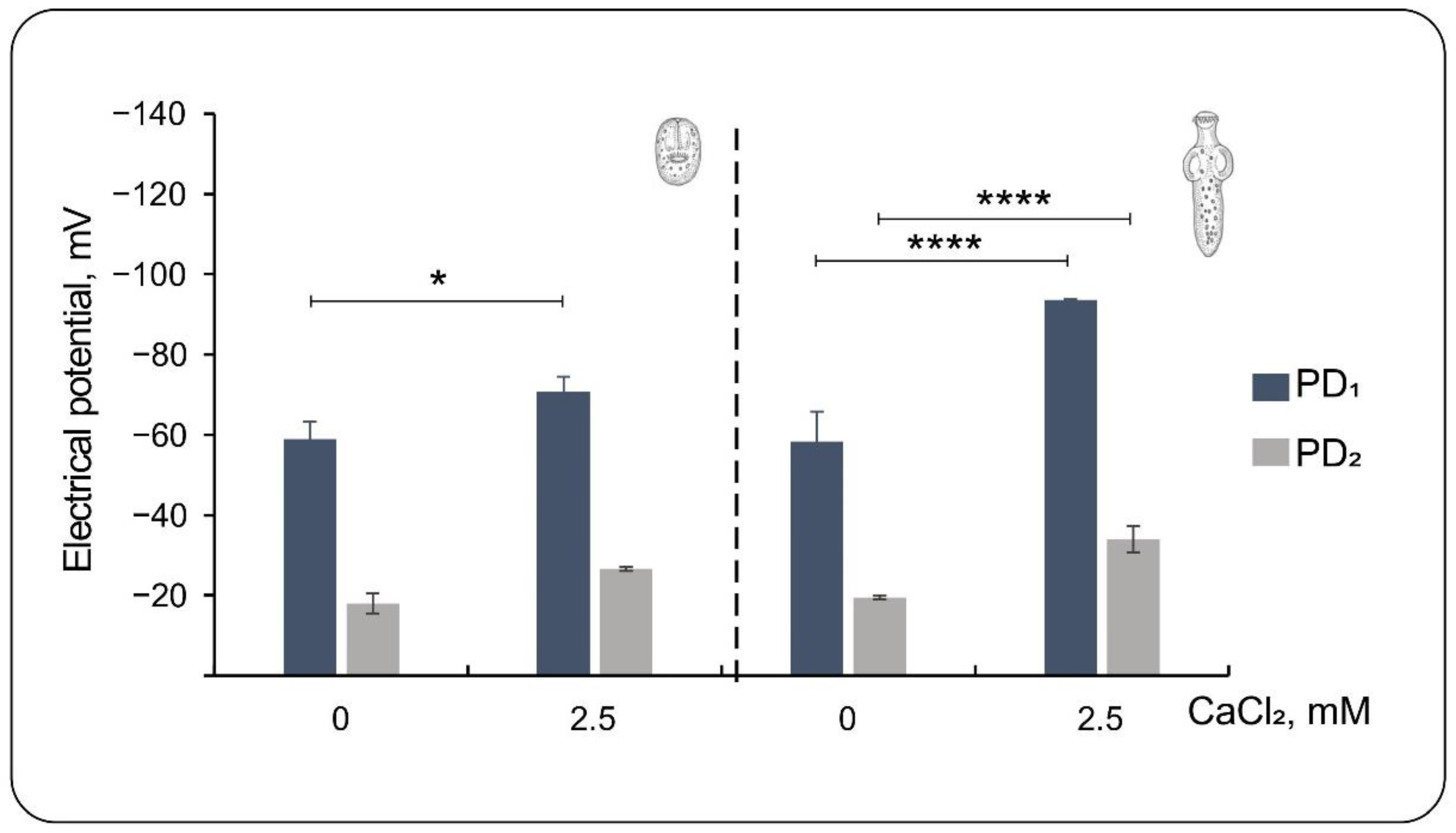
3.3. Effect of Low External Calcium
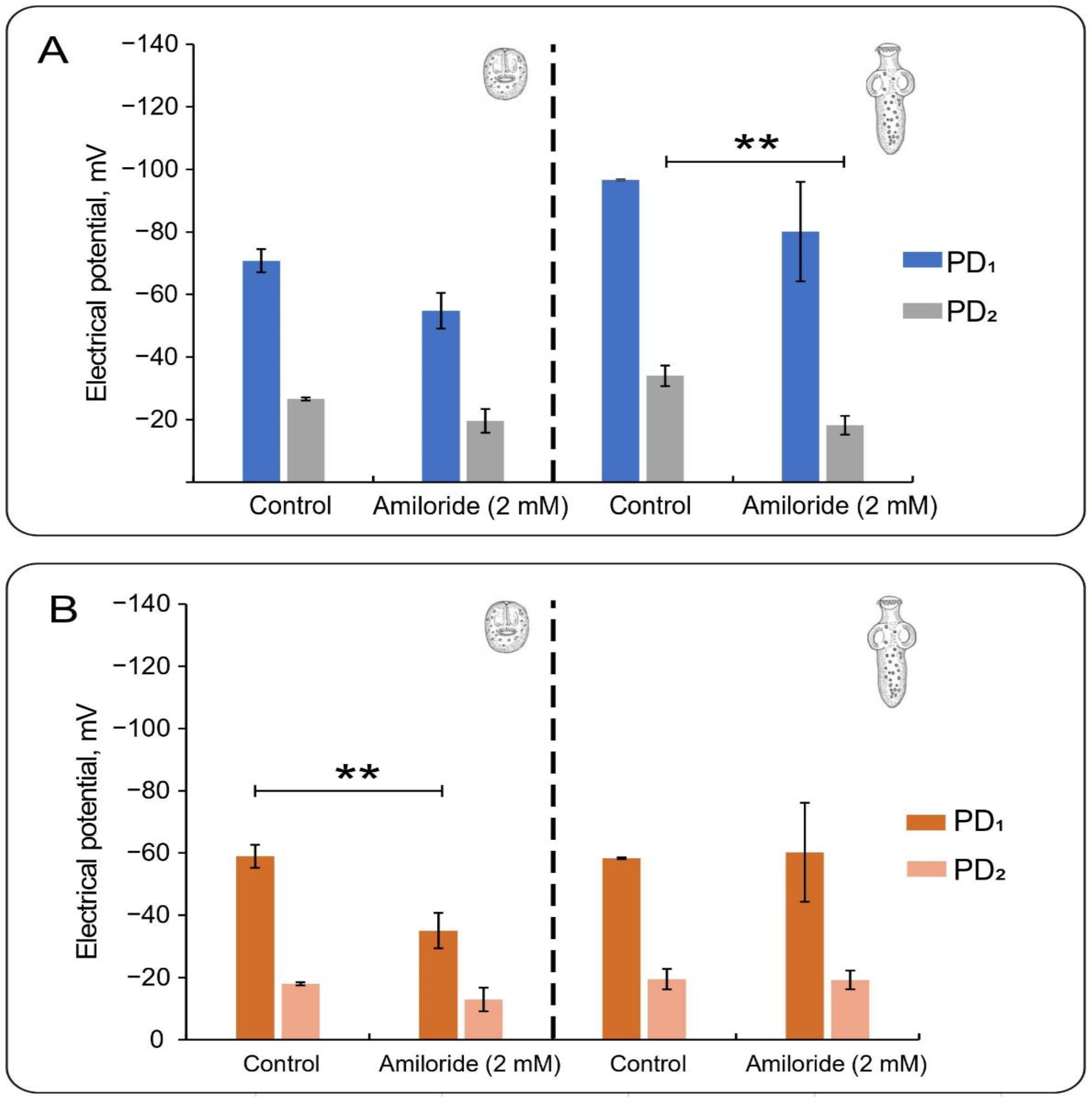
3.4. Effect of Amiloride
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Thompson, R. Biology and systematics of Echinococcus. In Advances in Parasitology; Chapter 2; Thompson, R.C.A., Lymbery, A.J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 65–109. [Google Scholar]
- Moro, P.; Schantz, P.M. Echinococcosis: A review. Int. J. Infect. Dis. 2009, 13, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrieu, E. Prevención y Control de la Hidatidosis en el Nivel Local: Iniciativa Sudamericana para el Control y Vigilancia de la Equinococosis Quística/Hidatidosis; Organización Panamericana de la Salud—OPS/OMS: Rio de Janeiro, Brazil, 2017; Volume 18, p. 56. [Google Scholar]
- Pavletic, C.F.; Larrieu, E.; Guarnera, E.A.; Casas, N.; Irabedra, P.; Ferreira, C.; Sayes, J.; Gavidia, C.M.; Caldas, E.; Lise, M.L.Z.; et al. Cystic echinococcosis in South America: A call for action. Rev. Panam. Salud Pública Pan Am. J. Public Health 2017, 41, e42. [Google Scholar]
- Cucher, M.A.; Macchiaroli, N.; Baldi, G.; Camicia, F.; Prada, L.; Maldonado, L.; Avila, H.G.; Fox, A.; Gutiérrez, A.; Negro, P. Cystic echinococcosis in south America: Systematic review of species and genotypes of Echinococcus granulosus sensu lato in humans and natural domestic hosts. Trop. Med. Int. Health 2016, 21, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Uchiumi, L.; Mujica, G.; Araya, D.; Salvitti, J.C.; Sobrino, M.; Moguillansky, S.; Solari, A.; Blanco, P.; Barrera, F.; Lamunier, J. Prevalence of human cystic echinococcosis in the towns of Ñorquinco and Ramos Mexia in Rio Negro province, Argentina, and direct risk factors for infection. Parasites Vectors 2021, 14, 1–10. [Google Scholar] [CrossRef]
- Menghi, C.I.; Arias, L.E.; Puzzio, C.P.; Gatta, C.L. Pulmonary hydatidosis from southern argentina. Trop. Parasitol. 2015, 5, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, D.P.; Geary, T.G. Helminth surfaces: Structural, molecular and functional properties. In Molecular Medical Parasitology; Academic Press: London, UK, 2003; pp. 297–338. [Google Scholar]
- Pérez-Serrano, J.; Grosman, C.; Urrea-París, M.; Denegri, G.; Casado, N.; Rodríguez-Caabeiro, F. Depolarization of the tegument precedes morphological alterations in Echinococcus granulosus protoscoleces incubated with ivermectin. Parasitol. Res. 2001, 87, 804–807. [Google Scholar] [CrossRef] [PubMed]
- Grosman, C.; Reisin, I.L. Echinococcus granulosus: Partial characterization of the conductive properties of two cation channels from protoscoleces of the ovine strain, reconstituted on planar lipid bilayers. Exp. Parasitol. 1995, 81, 546–555. [Google Scholar] [CrossRef]
- Grosman, C.; Reisin, I. Interconverting gating modes of a nonselective cation channel from the tapeworm Echinococcus granulosus reconstituted on planar lipid bilayers. J. Membr. Biol. 1997, 158, 87–94. [Google Scholar] [CrossRef]
- Zahran, F.; Ezz El-Din, H.M.; Shehata, M.A.S. Study on the effect of an ion channel inhibitor “fluralaner” on Echinococcus granulosus protoscolices and metacestode layers in vitro. J. Parasit. Dis. 2020, 44, 411–419. [Google Scholar] [CrossRef]
- Tsai, I.J.; Zarowiecki, M.; Holroyd, N.; Garciarrubio, A.; Sanchez-Flores, A.; Brooks, K.L.; Tracey, A.; Bobes, R.J.; Fragoso, G.; Sciutto, E.; et al. The genomes of four tapeworm species reveal adaptations to parasitism. Nature 2013, 496, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Kashyap, S.S.; Martin, R.J.; Robertson, A.P. Advances in our understanding of nematode ion channels as potential anthelmintic targets. Int. J. Parasitol. Drugs Drug Resist. 2022, 18, 52–86. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Robertson, A. Electrophysiological investigation of anthelmintic resistance. Parasitology 2000, 120, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Fetterer, R.; Pax, R.; Bennett, J. Schistosoma mansoni: Characterization of the electrical potential from the tegument of adult males. Exp. Parasitol. 1980, 49, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, C.; Reisin, I.L. Echinococcus granulosus: Characterization of the electrical potential of the syncytial tegument of protoscoleces incubated in vitro-effect of inhibitors. Exp. Parasitol. 1994, 78, 400–409. [Google Scholar] [CrossRef]
- Ferreira, A.; Trecu, T.; Reisin, I. Echinococcus granulosus: Study of the in vitro complement activation by protoscoleces by measuring the electric potential difference across the tegumental membrane. Exp. Parasitol. 1992, 75, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Reisin, I.; Ibarra, C.; Cybel, B.; Cantiello, H.F. Mecanismos de acción del mebendazol sobre el Echinococcus granulosus en su estadío larval estudiado in vitro. Bases para la interpretación de los resultados de la quimioterapia experimental y clínica Arch Intl Hidatid. 1988, 28, 442–470. [Google Scholar]
- Carabajal, M.P.A.; Durán, M.A.; Olivera, S.; Fernández Salom, M.J.; Cantiello, H.F. Electrical potentials of protoscoleces of the cestode Echinococcus granulosus from bovine origin. Exp. Parasitol. 2022, 238, 108282. [Google Scholar] [CrossRef]
- Ellory, J.C.; Willis, J.S. Temperature dependence of membrane function. The disparity between active potassium transport and (Na+ + K+) ATPase activity. Biochim. Biophys. Acta (BBA) Biomembr. 1976, 443, 301–305. [Google Scholar] [CrossRef]
- Swann, A.C. Brain (Na+,K+)-ATPase. Opposite effects of ethanol and dimethyl sulfoxide on temperature dependence of enzyme conformation and univalent cation binding. J. Biol. Chem. 1983, 258, 11780–11786. [Google Scholar] [CrossRef]
- Honig, A.; Oppermann, H.; Budweg, C.; Goldbecher, H.; Freyse, E.-J. Demonstration of temperature dependence of Na+-K+ pump activity of human blood cells. Adv. Physiol. Educ. 1994, 266, S10. [Google Scholar] [CrossRef]
- Cornelius, F.; Skou, J.C. The effect of cytoplasmic K+ on the activity of the Na+/K+-ATPase. Biochim. Biophys. Acta (BBA) Biomembr. 1991, 1067, 227–234. [Google Scholar] [CrossRef]
- Lagunes, R.; Ruiz, L.; Frixione, E. Contraction of epithelial (MDCK) cells in response to low extracellular calcium is dependent on extracellular sodium. J. Muscle Res. Cell Motil. 1999, 20, 761–770. [Google Scholar] [CrossRef]
- Williams, M.R.; Riach, R.A.; Collison, D.J.; Duncan, G. Role of the endoplasmic reticulum in shaping calcium dynamics in human lens cells. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1009–1017. [Google Scholar]
- Kleyman, T.R.; Cragoe, E.J., Jr. Amiloride and its analogs as tools in the study of ion transport. J. Membr. Biol. 1988, 105, 1–21. [Google Scholar] [CrossRef]
- Kleyman, T.R.; Cragoe, E.J., Jr. Cation transport probes: The amiloride series. Methods Enzymol. 1990, 191, 739–755. [Google Scholar] [PubMed]
- Greenberg, R.M. Ion channels and drug transporters as targets for anthelmintics. Curr. Clin. Microbiol. Rep. 2014, 1, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Fetterer, R.H.; Pax, R.A.; Bennett, J.L. Schistosoma mansoni: Direct method for simultaneous recording of electrical and motor activity. Exp. Parasitol. 1977, 43, 286–294. [Google Scholar] [CrossRef]
- Fetterer, R.; Pax, R.; Bennett, J. Na+-K+ transport, motility and tegumental membrane potential in adult male Schistosoma mansoni. Parasitology 1981, 82, 97–109. [Google Scholar] [CrossRef]
- Bricker, C.S.; Pax, R.; Bennett, J. Microelectrode studies of the tegument and sub-tegumental compartments of male Schistosoma mansoni: Anatomical location of sources of electrical potentials. Parasitology 1982, 85, 149–161. [Google Scholar] [CrossRef]
- Nor, B.; Young, N.D.; Korhonen, P.K.; Hall, R.S.; Tan, P.; Lonie, A.; Gasser, R.B. Pipeline for the identification and classification of ion channels in parasitic flatworms. Parasites Vectors 2016, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, V.; Docampo, R. Molecular and electrophysiological characterization of a novel cation channel of Trypanosoma cruzi. PLoS Pathog. 2012, 8, e1002750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, D.; Pax, R.; Bennett, J. Microelectrode studies of the tegument and sub-tegumental compartments of male Schistosoma mansoni: An analysis of electrophysiological properties. Parasitology 1982, 85, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Sariban-Sohraby, S.; Benos, D. The amiloride-sensitive sodium channel. Am. J. Physiol. Cell Physiol. 1986, 250, C175–C190. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, J.; Vereecke, J.; Carmeliet, E. Mechanism of cardiac T-type Ca2+ channel blockade by amiloride. J. Pharmacol. Exp. Ther. 1990, 254, 546–551. [Google Scholar]
- Doi, Y.; Marunaka, Y. Amiloride-sensitive and HCO3(-)-dependent ion transport activated by aldosterone and vasotocin in A6 cells. Am. J. Physiol. Cell Physiol. 1995, 268, C762–C770. [Google Scholar] [CrossRef]
- Murata, Y.; Harada, K.; Nakajima, F.; Maruo, J.; Morita, T. Nonselective effects of amiloride and its analogs on ion transport systems and their cytotoxicities in cardiac myocytes. Jpn. J. Pharmacol. 1995, 68, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Stoner, L.; Viggiano, S. Apical nonspecific cation channels in everted collecting tubules of potassium-adapted Ambystoma. J. Membr. Biol. 2000, 177, 109–116. [Google Scholar] [CrossRef]
- Hirsh, A.J. Altering airway surface liquid volume: Inhalation therapy with amiloride and hyperosmotic agents. Adv. Drug Deliv. Rev. 2002, 54, 1445–1462. [Google Scholar] [CrossRef]
- Cantiello, H.; Ibarra, C.; Reisin, I. Flujos de K+ y potenciales eléctricos a través de las membrana de Echinococcus granulosus incubado “in vitro”. In IX Reunión Anual de la Sociedad Argentina de Biofísica; Bermejo: Mendoza, Argentina, 1980. [Google Scholar]
- Koziol, U.; Brehm, K. Recent advances in echinococcus genomics and stem cell research. Vet. Parasitol. 2015, 213, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, P.; Da Silva, A.M.; Akhan, O.; Müllhaupt, B.; Vizcaychipi, K.; Budke, C.; Vuitton, D. The echinococcoses: Diagnosis, clinical management and burden of disease. Adv. Parasitol. 2017, 96, 259–369. [Google Scholar]
- Lacey, E. Mode of action of benzimidazoles. Parasitol. Today 1990, 6, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Doenhoff, M.J.; Cioli, D.; Utzinger, J. Praziquantel: Mechanisms of action, resistance and new derivatives for schistosomiasis. Curr. Opin. Infect. Dis. 2008, 21, 659–667. [Google Scholar] [CrossRef]
- Wu, W.; Wang, W.; Huang, Y.-X. New insight into praziquantel against various developmental stages of schistosomes. Parasitol. Res. 2011, 109, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Prole, D.L.; Taylor, C.W. Identification of intracellular and plasma membrane calcium channel homologs in pathogenic parasites. PLoS ONE 2011, 6, e26218. [Google Scholar] [CrossRef] [PubMed]
- Lader, A.S.; Kwiatkowski, D.J.; Cantiello, H.F. Role of gelsolin in the actin filament regulation of cardiac L-type calcium channels. Am. J. Physiol. Cell Physiol. 1999, 277, C1277–C1283. [Google Scholar] [CrossRef]
- Durán, M.A.; Olivera, S.R.; Cantiello, H.F. Increase in pharmacological potency of praziquantel by cytoskeletal modifiers in protoscoleces of Echinococcus granulosus. In XXXIII Jornadas Nacionales de Hidatidosis; Asociación Parasitológica Argentina: Catamarca, Argentina, 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ionic Composition (mM) | T (°C) | Invaginated PSC | Evaginated PSC | ||
|---|---|---|---|---|---|
| PD1 | PD2 | PD1 | PD2 | ||
| 121 Na+ 5 K+ 2.5 Ca2+ | 37 | −70.8 ± 3.7 (55) | −26.6 ± 0.5 (55) | −93.6 ± 0.3 (55) | −34.0 ± 3.3 (55) |
| 4 | −140.0 ± 5.2 (31) | −39.1 ± 2.9 (31) | −86.2 ± 9.2 (15) | −25.9 ± 2.8 (15) | |
| 15 Na+ 110 K+ 2.5 Ca2+ | 37 | −38.6 ± 4.1 (30) | −21.2 ± 0.4 (30) | −64.1 ± 4.8 (60) | −26.7 ± 1.7 (60) |
| 4 | −40.0 ± 3.8 (20) | −16.5 ± 2.1 (20) | −45.2 ± 0.6 (21) | −18.1 ± 1.6 (21) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carabajal, M.P.A.; Fernández Salom, M.J.; Olivera, S.; Cantiello, H.F. Effect of Temperature and Ionic Substitutions on the Tegumental Potentials of Protoscoleces of Echinococcus granulosus. Trop. Med. Infect. Dis. 2023, 8, 303. https://doi.org/10.3390/tropicalmed8060303
Carabajal MPA, Fernández Salom MJ, Olivera S, Cantiello HF. Effect of Temperature and Ionic Substitutions on the Tegumental Potentials of Protoscoleces of Echinococcus granulosus. Tropical Medicine and Infectious Disease. 2023; 8(6):303. https://doi.org/10.3390/tropicalmed8060303
Chicago/Turabian StyleCarabajal, Mónica Patricia Antonella, María José Fernández Salom, Santiago Olivera, and Horacio F. Cantiello. 2023. "Effect of Temperature and Ionic Substitutions on the Tegumental Potentials of Protoscoleces of Echinococcus granulosus" Tropical Medicine and Infectious Disease 8, no. 6: 303. https://doi.org/10.3390/tropicalmed8060303
APA StyleCarabajal, M. P. A., Fernández Salom, M. J., Olivera, S., & Cantiello, H. F. (2023). Effect of Temperature and Ionic Substitutions on the Tegumental Potentials of Protoscoleces of Echinococcus granulosus. Tropical Medicine and Infectious Disease, 8(6), 303. https://doi.org/10.3390/tropicalmed8060303