1. Introduction
Over recent decades, both the atmosphere and oceans have warmed, sea level has risen, snow cover and ice sheets have diminished, all whilst the atmospheric concentration of greenhouse gases have continued to increase [
1]. Although changes have been observed globally, it is mostly agreed that northern high latitude and alpine regions are amongst the most vulnerable to climate change—currently warming at 0.6 °C per decade; over twice as fast as the global average [
1,
2,
3,
4]. Such regions hold a large portion of global organic carbon with approximately twice as much carbon being stored in frozen Arctic soil or permafrost as is currently in the atmosphere [
5,
6,
7,
8]. There is the potential for this carbon to be released into the atmosphere as greenhouse gases under permafrost thaw, creating a positive feedback loop onto climate [
6,
9,
10,
11,
12].
There are an increasing number of both modelling and experimental studies which are beginning to explore the impact of an interactive nitrogen (N) cycle on the permafrost carbon feedback (for example, [
13,
14,
15,
16]). Generally, Arctic vegetation is strongly nitrogen-limited due to the supply of available soil mineral N being lower than the demands of the plant, reducing net primary productivity (NPP) [
17,
18,
19,
20]. However, with rising temperatures, there is increased ‘greenness’ around the circumpolar Arctic [
21,
22], which is likely a result of increased biomass [
21], changes in plant community composition [
21,
23,
24,
25,
26], and changing plant phenology [
21,
27,
28]. Although there are reports of background increases in tundra vegetation [
21,
25,
29,
30,
31], Arctic shrub expansion is widely reported throughout the literature [
21,
22,
32], with shrub species, such as birch, willow, and alder increasing in abundance, cover and height across the region [
22]. Increases in shrub abundance can be attributed to various processes, such as rising temperatures, permafrost thaw, tundra fires, anthropogenic activity, etc. [
22]. Rising temperatures can promote shrub growth either directly through physiological processes or indirectly by enhancing soil microbial activity such as increased nitrogen mineralisation which supplies nutrients for shrub uptake [
22]. Arctic amplification of shrubs and other Arctic tundra also varies between species. For example, taller shrub species have been observed to have greater nitrogen availability and faster nitrogen cycling compared to shorter, low shrubs [
22,
32,
33]. Whilst shrubs that are able to grow in cover or height can restrict the growth of other species by limiting light [
22,
34], suggesting some species may have a competitive advantage under further future warming. The interactions between this enhanced vegetation growth and the availability of mineral nitrogen is uncertain and will impact the magnitude and timing of the permafrost carbon feedback [
6,
13].
Recent experimental studies at selected sites suggest that nitrogen at depth in the soil (organic and inorganic), previously unavailable to plants due to its frozen state, could be made accessible by warming-induced permafrost thaw and reduce plant nitrogen limitation in the future [
13,
14,
15,
35,
36,
37,
38,
39,
40]. Furthermore, there is evidence that future warming may result in an increased abundance of Arctic plant roots with depth, further increasing plant uptake of previously unavailable nutrients [
40,
41,
42,
43,
44]. For example, certain shrub species such as the dwarf birch
Betula nana is able to take advantage of increased temperatures and subsequent thaw depth, by rapidly elongating its short shoots [
22]. This suggests that increased nitrogen availability could reduce or even outpace nitrogen demand of Arctic vegetation and hence nitrogen limitation [
36].
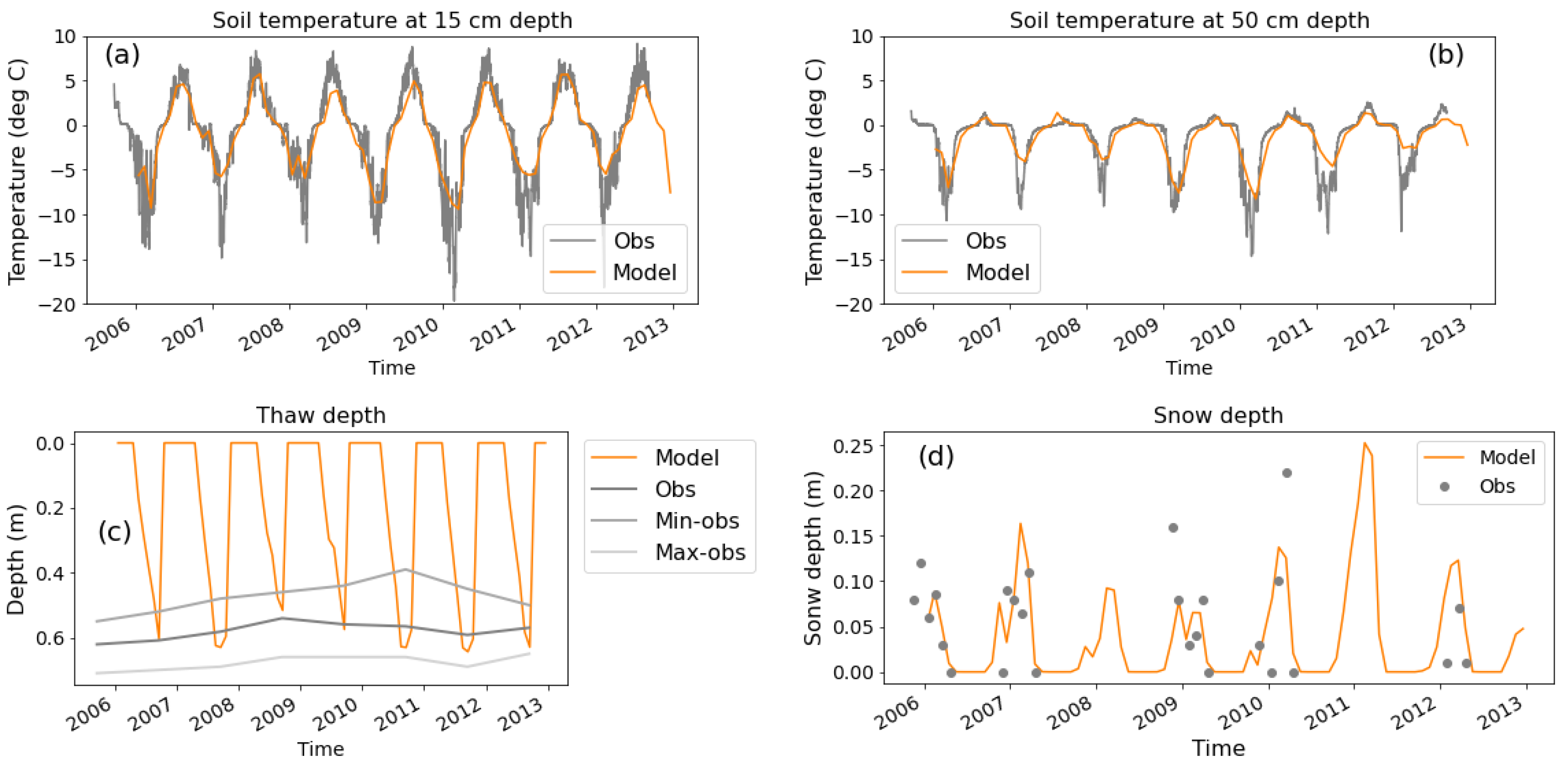
In this study, we run a version of the JULES land surface model for an experimental site in Abisko, Sweden, where near-surface and thaw-front nitrogen addition experiments were conducted, to evaluate the ability of JULES to simulate the response of plants to increased mineral nitrogen availability within the soil. In addition, we ran simulations driven by the projected changes in climate in the future to quantify the potential impact of permafrost thawing on the mineral nitrogen and the subsequent response of vegetation carbon in plausible future climates.
4. Discussion
It has been shown that subarctic permafrost peatlands contain large amounts of plant-available nitrogen which may be released upon future thawing [
13,
35]. Such change in nutrient availability will impact (usually nitrogen-limited) vegetation by altering composition, productivity, and biomass [
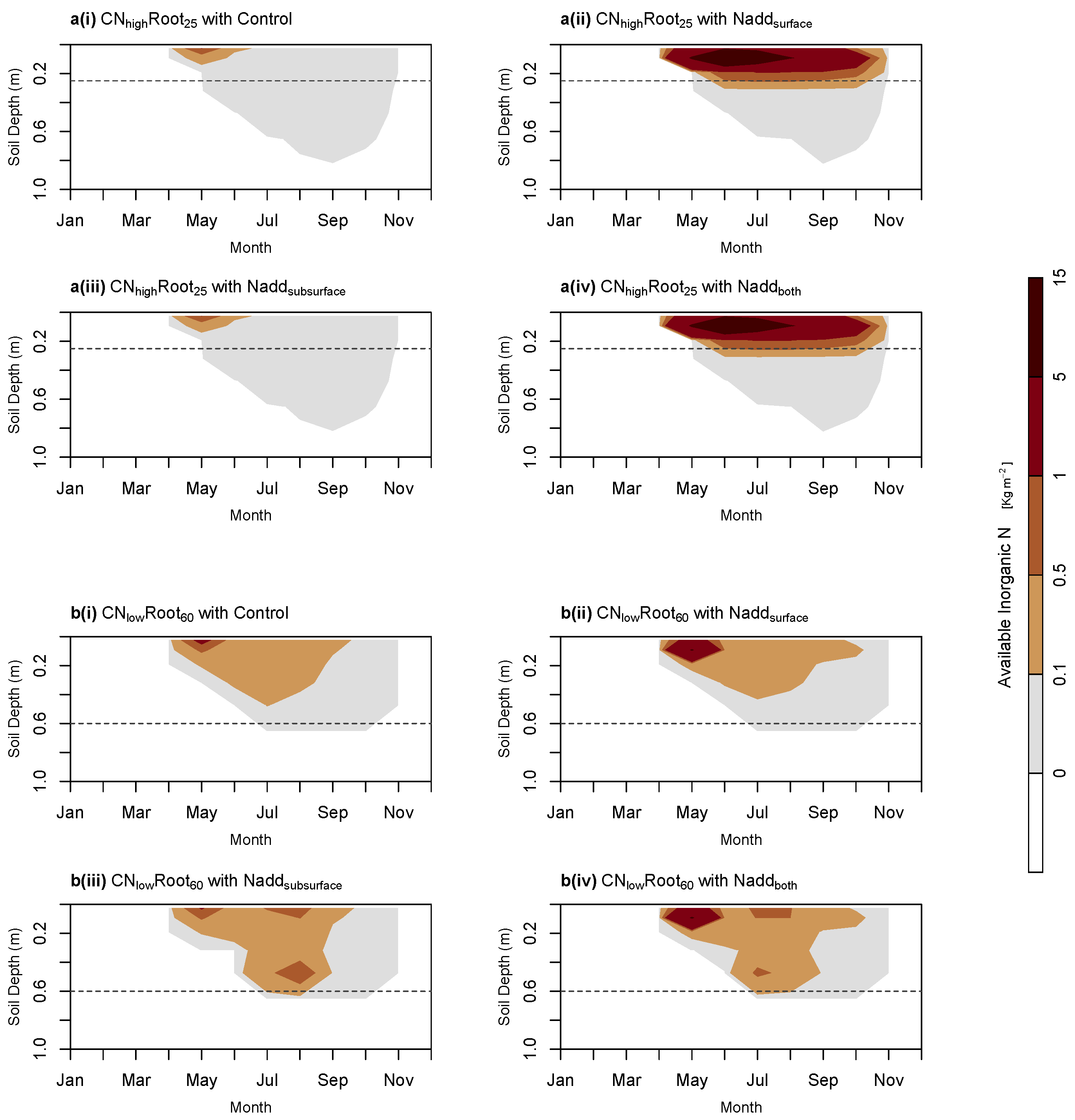
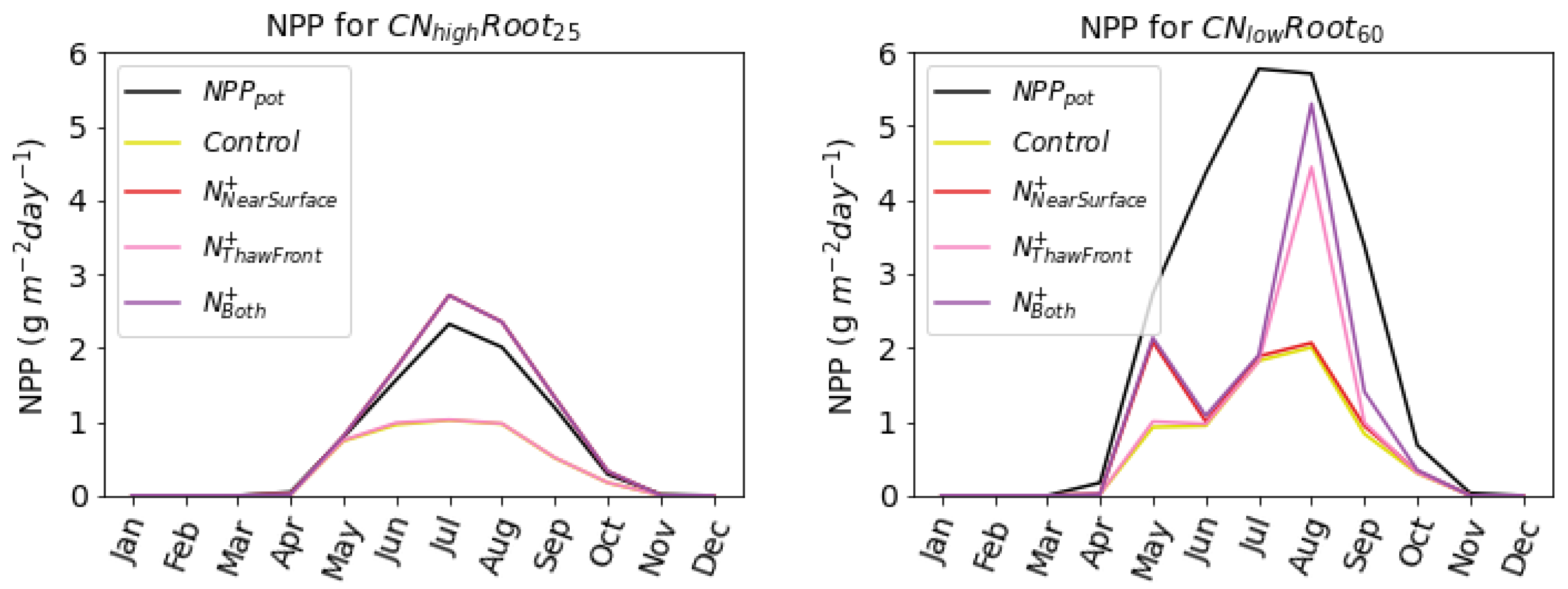
40] and could potentially counteract some of the carbon loss from thawing permafrost. Here, we show that it is possible to simulate belowground fertilisation in JULES by successfully replicating observations seen from previous fertilisation experiments. Our results show that near-surface fertilisation increases nitrogen available to both shallow- and deep-rooting species (
Figure 4). However, only the deep-rooting species benefit from nitrogen applied at the permafrost thaw front. This is consistent with the experimental study conducted by [
40]. In their study, [
40] also found that deep-rooting plants were able to take up nutrients at the end of the growing season at the thaw front. This can be seen in our results by the substantial amount of nitrogen available to the deep-rooting plant species (
) near the thaw front where subsurface fertiliser has been applied (
Figure 4b(iii,iv)) and by the increase in NPP in
Figure 5. Since the largest thaw depth is reached at the end of the growing season, this ability to take up nitrogen late in the season is key to whether or not plant species can benefit from the nitrogen released from thawing permafrost in the future. These results contradict the assumption from both [
63,
64] that seasonal asynchrony between nitrogen-availability at the thaw front and plant nitrogen-demand means that nitrogen released from thawing permafrost will not affect biomass, and our study shows that the impact on biomass can be significant indeed.
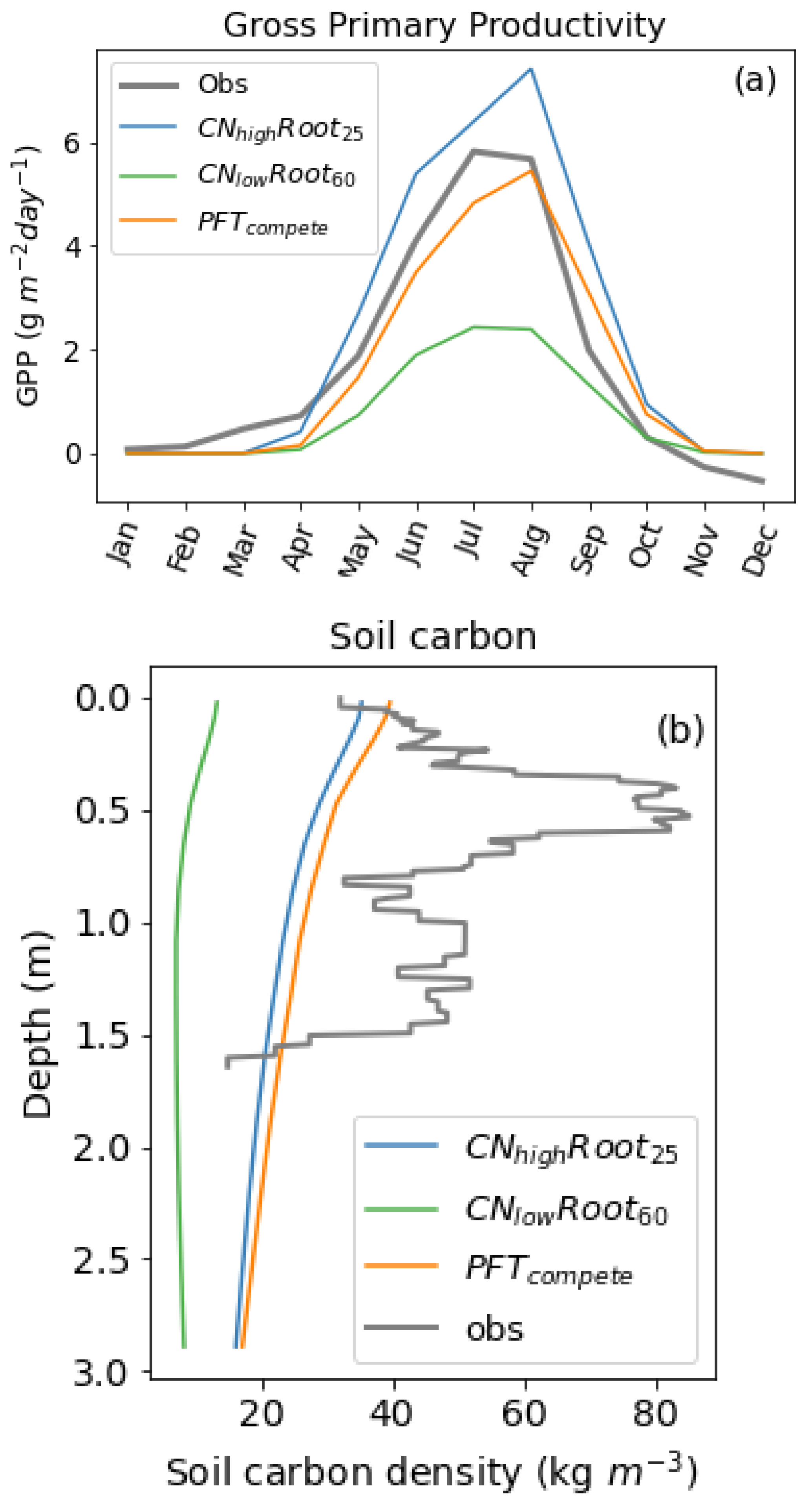
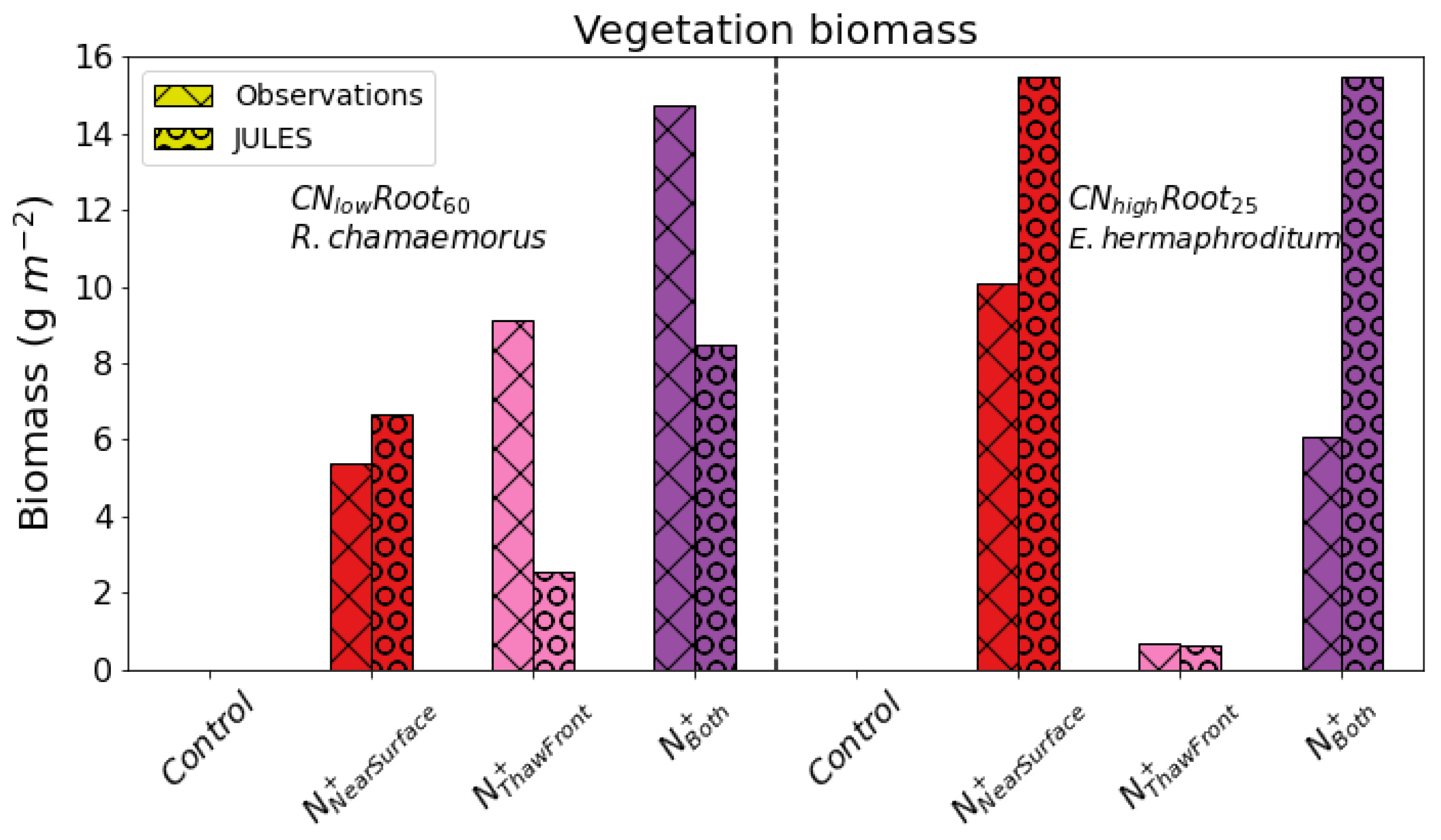
Although broadly our results are similar to [
40], the simulated increase in aboveground biomass in response to fertilisation differs from the values found in the [
40] study. In particular, [
40] showed the fertilisation at the thaw front had a larger affect on the aboveground biomass of the deep-rooting species (
R. chamaerorus) than near-surface fertilisation, whereas our
saw the opposite response. This difference is likely explained by the relative amounts of roots in the soil profile, where larger a larger abundance of roots is observed at the thaw front for deep-rooting species compared to what is assumed in JULES. In addition, the deep-rooting PFT (
) has approximately 10 gm
more aboveground biomass than our shallow-rooting PFT (
), which is contrary to that found by [
40]. There is also an approximately double relative increase in aboveground biomass for
compared to
when near surface fertiliser was added. This is likely caused by differences in nitrogen-limitation, with shallow-rooting species being more nitrogen-limited near the surface (
Figure 5).
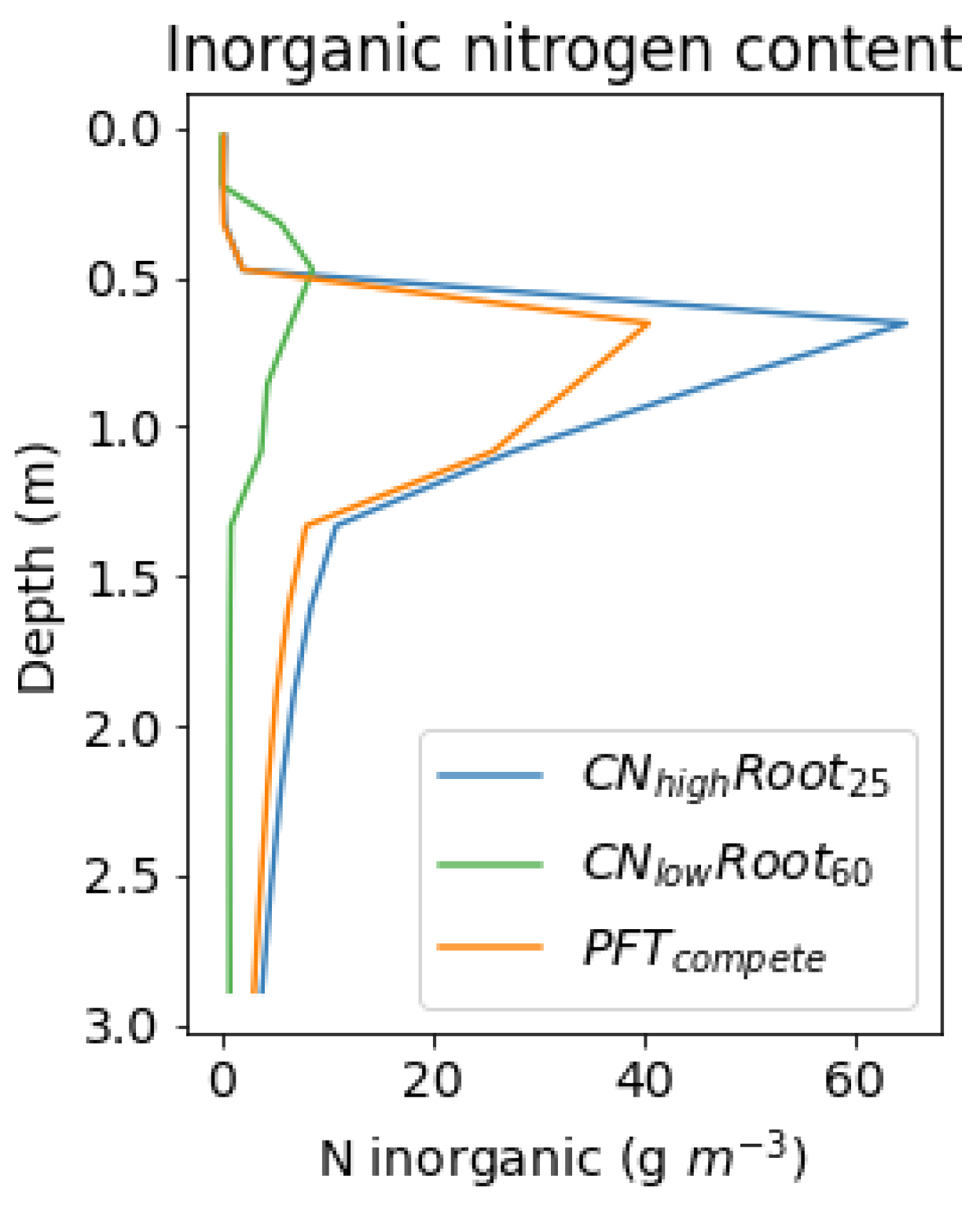
Future projections show that increases in plant-available inorganic nitrogen from present to the end of the century under climate warming are broadly similar to the changes obtained from the fertilisation at the thaw front in the belowground fertilisation simulations (
Table 4). This results in increases of productivity and subsequent changes in vegetation. Nitrogen mineralisation is a key process that provides plant-available inorganic nitrogen at depth and is affected by multiple factors, such as hydrology, biochemistry, oxygen availability, and soil temperatures [
2,
11,
65]. Therefore, under future warming, nitrogen mineralisation at the thaw front could be impacted through the response of such factors. Nitrogen mineralisation is known to be highly sensitive to temperature change [
35,
66,
67,
68]. In shallow soil layers, plant-available nitrogen is enhanced due to increased mineralisation due to air temperatures [
67,
68]. Whilst warmer temperatures in deeper layers of permafrost soils can lead to warming-induced increased microbial activity and increased mineralisation rates of organic matter [
66]. A study conducted by [
35] investigating mineralisation sensitivity to increased temperatures in permafrost soils found a five-fold increase in mineralisation rates over 0.5–11 °C, highlighting the importance that nitrogen mineralisation might play in providing sources of inorganic nitrogen for plant uptake at the thaw front in the future. Thawing permafrost may also provide soil drainage and new oxygen availability which could further enhance mineralisation rates [
66]. However, mineralisation rates may respond differently by different thawing scenarios [
35]. For example, abrupt permafrost thaw can lead to the formation of permafrost ponds and wetland which, in turn, might limit mineralisation through soil moisture [
69].
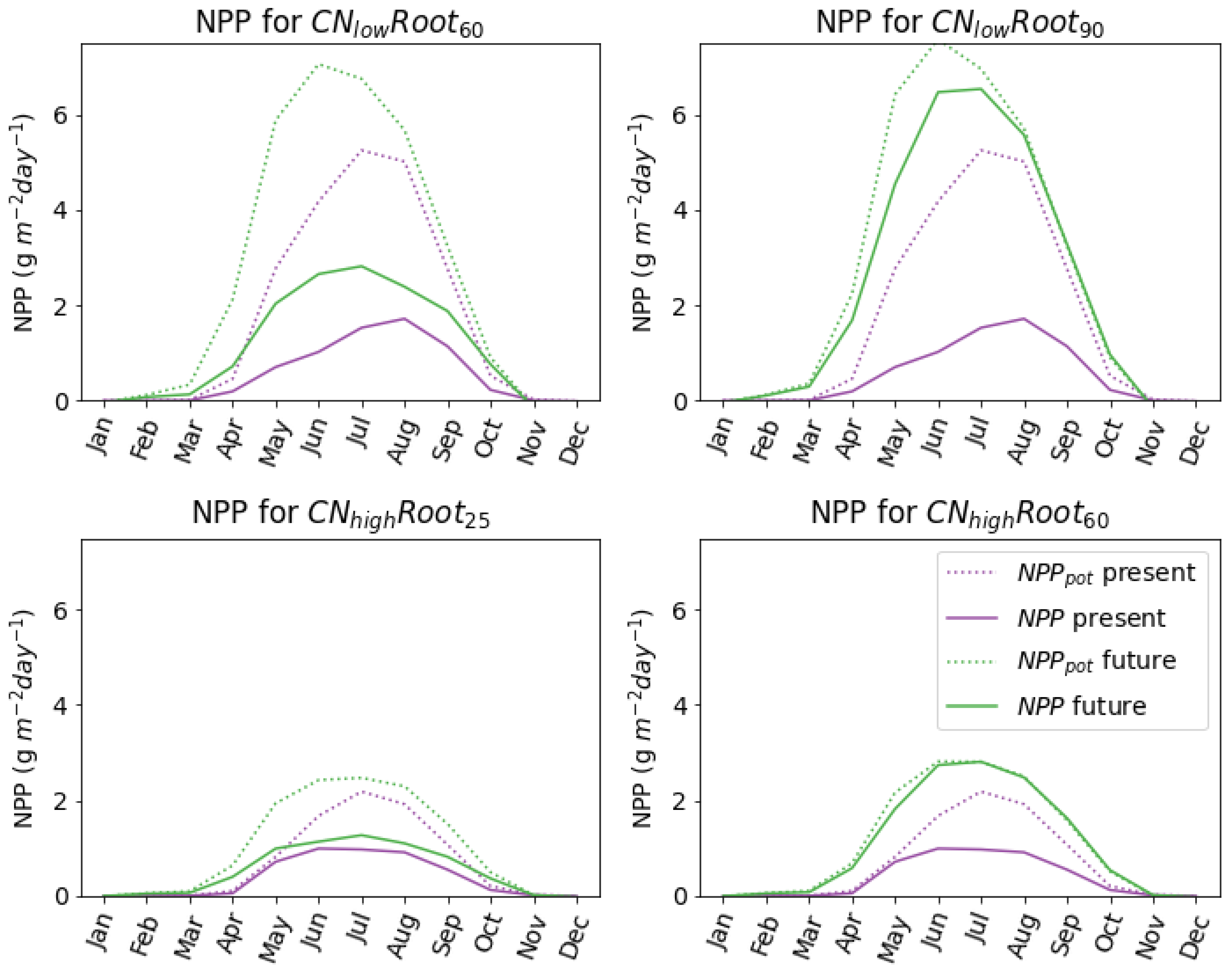
The effects of increased nitrogen availability and uptake are enhanced when the simulated root depth is allowed to increase with increased summer thaw depth. Previous work suggests that a mismatch in time between nutrient demand and nitrogen availability will result in a negligible impact on plant root dynamics with future permafrost thaw [
63,
64,
70]. However, recent research suggests that plant rooting depth can increase in thawing permafrost ecosystems [
40,
42,
43,
44], suggesting that having deep roots could be more advantageous under future warming and could result in vegetation shifts [
71,
72]. In our simulations, the largest pools of plant-available inorganic nitrogen were seen when species roots were extended to account for future root plasticity (
Table 4), resulting in higher uptake and productivity compared to species with no root growth into greater depths. Inorganic nitrogen uptake by the species with no root adaptation increased by the end of the century (54% and 75% for
and
, respectively). However, once the roots were allowed to adapt and grow deeper in the soil, the inorganic nitrogen uptake increased substantially—an increase of up to 300% when compared to present (
Table A3). Increasing inorganic nitrogen uptake reduces nitrogen-limitation, and consequently, simulations with increased root depths had the largest increases in plant productivity and vegetation biomass (
Figure 9). In fact, the
species with adapted roots (
) became no longer nitrogen-limited in the future (
Figure 8) and had an extra 16% increase in aboveground biomass compared with the
species. This suggests that species that are able to extend roots deep into the soil profile will have a competitive advantage under future warming. This has been observed in Arctic permafrost peatlands. For example, [
73] found a change from shrub-dominated hummock sites to graminoid-dominated in Sweden over a 30-year period in response changes in frozen ground.
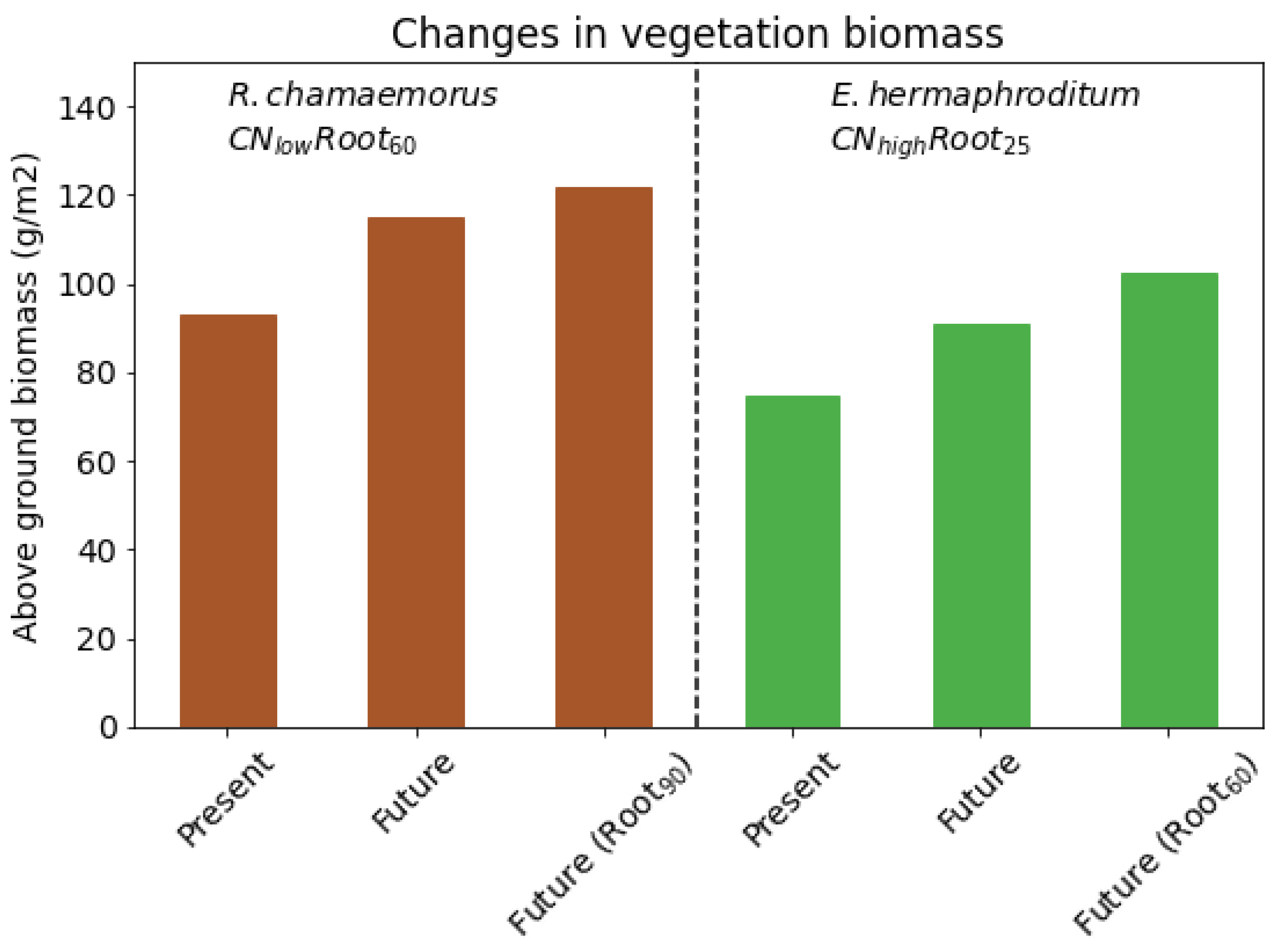
There are around 1.7 million km
of permafrost peatlands [
74]. The shallow-rooting species (
) has about 90 gm
aboveground biomass in the present day, which converts to 0.15 Gt C over the permafrost peatlands. An increase of 22% gives an extra 0.033 Gt C of aboveground biomass for this species over the permafrost peatlands. If this species grows deeper roots, there is an increase of 33% in the aboveground biomass which gives an extra 0.05 Gt C. The extent of the permafrost region is estimated to be 21 million km
with approximately 14 million km
underlain by permafrost [
75]. If the response of the shallow rooting species in JULES is extrapolated to the entire permafrost region, there is an extra 1.2–1.9 Gt C of carbon in the aboveground vegetation. Our results demonstrate an increase in future projections of carbon storage through enhanced biomass when including root adaptation and growth in models. This process has not previously been accounted for in Earth System Models. Therefore, it needs to be quantified for a range of Arctic vegetation types and its impact on climate–carbon feedbacks quantified in future Earth System models.
We assume that the nitrogen cycling represented within JULES has a realistic response to a changing climate. However, it was developed specifically to capture the large-scale interaction of carbon and nitrogen in a simple way [
53] and therefore has limitations. In particular, it assumes fixed plant stoichiometry. Recent analyses of field observations have observed changes in leaf nitrogen content in response to increased nitrogen availability [
76]. The inclusion of flexible stoichiometry into JULES has the potential to alter the carbon and nitrogen cycling and the impact of climate change on the biogeochemical feedbacks in the future. Competition between species was not included in the simulations in this paper. Here, only Arctic
PFT was allowed to grow. However, there is a high dominance of
Sphagnum fuscum moss throughout the site [
40]. This is not accounted for in JULES and will impact both the surface water and energy balance and the biogeochemical cycling, including the soil net mineralisation. By only considering one PFT in this way, we are not considering the complete response of Arctic vegetation to future climate change such as plant species change. For example, although shrub expansion is often considered in tundra ecosystems, abrupt permafrost thaw can result in the formations of thaw ponds which can lead to vegetation shifts towards graminoid-dominated wetland [
69]. Furthermore, shrub growth can be limited by wet soil conditions and nutrient-limitation whilst deep-rooting graminoid species may have the advantage of accessing newly available nutrients such as inorganic nitrogen under future permafrost thaw [
69]. There are also uncertainties surrounding the interactions between Arctic vegetation response to increased nitrogen availability and impacts on nutrient cycling. As an example, as vegetation such as shrubs increase in height and cover as a result of warming, canopy overtopping might reduce the capacity to respond to warming induced changes [
22]. However, such increases can also increase litter input to soils and enhance nitrogen mineralisation rates [
33,
77,
78]. Therefore, any projections of vegetation responses in arctic ecosystems contain uncertainties surrounding overall response until these interactions are better understood and represented in models. Similarly, although an increase of root abundance of Arctic species at the thaw front is expected under future warming [
43,
44], there is relatively little known regarding root distribution with depth. In our simulations, we assumed a linearly decreasing distribution as a function of depth; however, alternatives are suggested and may differ for different species or ecosystems [
41]. Although peat processes have recently been added to JULES [
62], they are not yet included in this model configuration, and will impact the simulation of the soil carbon and nitrogen distribution and hence the available inorganic nitrogen.
Ultimately JULES is a model and we have tested a single process within that model: response to increased soil nitrogen-availability, both to test whether the representation of this process is realistic and to illustrate its potential role under future climate change. The bigger question is what the future of the Arctic system is as a whole, and to evaluate this, there are many interacting processes that must be considered, such as shifts in vegetation composition, hydrological and biogeochemical processes, some of which are included in JULES and some of which still need to be developed. For instance, most terrestrial nitrogen cycling models assume that plants can only utilise inorganic forms of nitrogen, yet it is known that Arctic vegetation can directly access organic nitrogen [
79]. Furthermore, a study by [
80] found that modelled organic nitrogen accounted for 36–87% of total nitrogen uptake by plants in tundra ecosystems with others estimating that nearly 60% of the nitrogen uptake by plants is from free amino acids in the arctic tundra [
81]. Organic nitrogen may have different controls compared to inorganic nitrogen considered in our study and could play a very important role in plant nutrition under future warming. For example, a study by [
35] that measured organic nitrogen (extractable, mineralizable and plant-available) and found consistently higher amounts in deeper layers of the soils. Therefore, future permafrost thaw and vegetation shifts under future climate change could be heavily involved with organic nitrogen; yet, this process is generally not included in land surface models. There are also vegetation processes such as root suberization and re-translocation of nutrients into storage tissues that will affect their response to additional nitrogen availability [
35] and these are generally not included in models either. For example, thaw-depth development synchrony with plant nitrogen demand has the potential to be mismatched, whilst some species such as
R. chamaemorus as well as deep-rooting graminoid species have been found to take up nitrogen by the time a meaningful change in thaw depth has occurred [
35,
82].
It is therefore important to recognise the limitations of using a model as presented in this study in presenting the complete picture of the arctic system under future climate change. However, considering the strong nitrogen limitation in the Arctic overall, we expect that the addition of mineralised nitrogen from permafrost has a role to play in the overall response of the Arctic ecosystem to climate change, and our results support this hypothesis, albeit that they illustrate only one aspect of the response. Similarly, by testing a model’s ability in representing one response such as belowground fertilisation, we can get closer to a more complete representation of the Arctic system and thus closer to getting the whole ecosystem response right.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








