The Use of Plant Macrofossils for Paleoenvironmental Reconstructions in Southern European Peatlands


Abstract
:1. Introduction
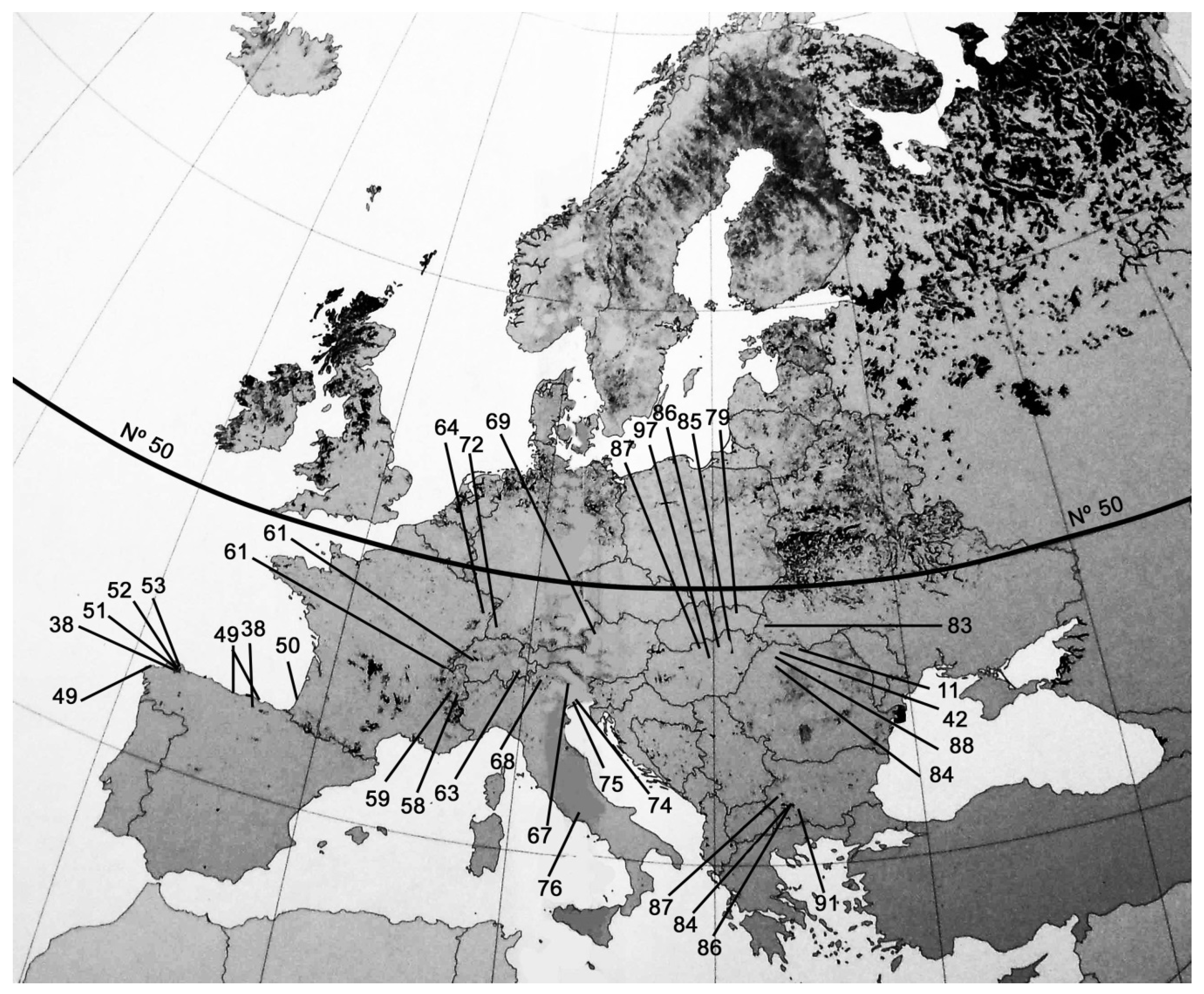
2. Material and Methods
3. Results and Discussion
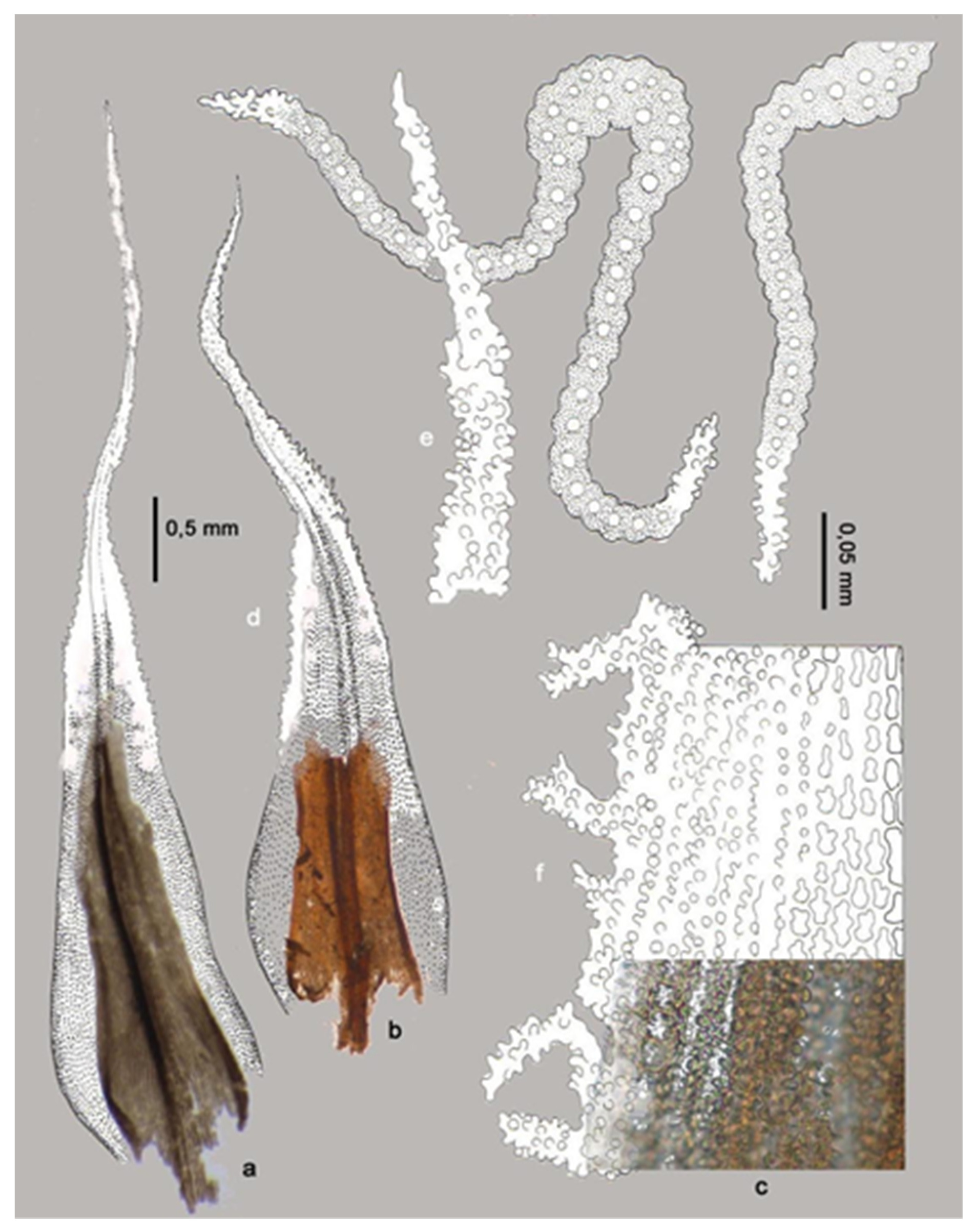
3.1. Problems in the Identification of Plant Macrofossils
3.2. Macrofossils: West Zone (Iberian Peninsula)
3.3. Macrofossils: Central Zone (Alps)
3.4. Macrofossils East Zone
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clymo, R.S. Peat. In Ecosystems of the World. Mires: Swamp, Bog, Fen and Moor; Gore, A.J.P., Ed.; Elsevier: Amsterdam, The Netherlands, 1983; Volume 4A, pp. 159–224. [Google Scholar]
- Joosten, H. Mires in Europe: A preliminary status report. Int. Mire Conserv. Group Memb. Newslett. 1997, 3, 10–13. [Google Scholar]
- Joosten, H. Peat the final frontier: Mires and peatlands outside the tropics. In Peatlands under Pressure. Arctic to Tropical Peatlands; Maltby, E., Maclean, L., Eds.; Royal Holloway Institute for Environmental Research: Egham, UK, 1999; pp. 9–17. [Google Scholar]
- European Commission. European Commission Interpretation Manual of European Union Habitats, Vers. EUR28; European Commission, DG Environment: Brussel, Belgium, 2013. [Google Scholar]
- Barber, K.E.; Chambers, F.M.; Maddy, D.; Stoneman, R.E.; Brew, J.S. A sensitive highresolution record of late Holocene climate change from a raised bog in northern England. Holocene 1994, 4, 198–205. [Google Scholar] [CrossRef]
- Blackford, J. Palaeoclimatic records from peat bogs. Trends Ecol. Evol. 2000, 15, 193–198. [Google Scholar] [CrossRef]
- Birks, H.H.; Birks, H.J.B. Future uses of pollen analysis must include plant macrofossils. J. Biogeogr. 2000, 27, 31–35. [Google Scholar] [CrossRef]
- Birks, H.H.; Birks, H.J.B. Reconstructing Holocene climates from pollen and plant macrofossils. In Global Change in the Holocene; Mackay, A., Battarbee, R.W., Birks, H.J.B., Oldfield, F., Eds.; Arnold: London, UK, 2003; pp. 342–357. [Google Scholar]
- Kołaczek, P.; Gałka, M.; Apolinarska, K.; Gębica, P.; Superson, S.; Michno, A.; Harmata, K.; Szczepanek, K.; Płóciennik, M.; Gąsiorowski, M.; et al. Lost in dating—Problems with the absolute chronologies and sedimentation rates of late glacial and early Holocene oxbow lake deposits in Central Europe. Quat. Geochronol. 2017, 41, 187–201. [Google Scholar] [CrossRef]
- Kołaczek, P.; Gałka, M.; Lamentowiczac, M.; Marcisz, K.; Kajukało-Drygalska, K.; Karpińska-Kołaczeka, M. Increased radiocarbon dating resolution of ombrotrophic peat profiles reveals periods of disturbance which were previously undetected. Quat. Geochronol. 2019, 52, 21–28. [Google Scholar] [CrossRef]
- Feurdean, A.; Gałka, M.; Tanţău, I.; Geantă, A.; Hutchinson, S.M.; Hickler, T. Tree and timberline shifts in the northern Romanian Carpathians during the Holocene and the responses to environmental changes. Quat. Sci. Rev. 2016, 134, 100–113. [Google Scholar] [CrossRef]
- Gałka, M.; Tobolski, K.; Górska, A.; Lamentowicz, M. Resilience of plant and testate amoeba communities after climatic and anthropogenic disturbances in a Baltic bog in Northern Poland: Implications for ecological restoration. Holocene 2017, 27, 130–141. [Google Scholar] [CrossRef]
- Gałka, M.; Tobolski, K.; Lamentowicz, L.; Ersek, V.; Jaseey, V.E.; van der Knaap, W.O.; Lamentowicz, M. Unveiling exceptional Baltic bog ecohydrology, autogenic succession and climate change during the last 2000 years in CE Europe using replicate cores, multi-proxy data and functional traits of testate amoebae. Quat. Sci. Rev. 2017, 156, 90–106. [Google Scholar] [CrossRef] [Green Version]
- Coulson, J.C.; Butterfield, J. An investigation of the biotic factors determining the rates of plant decomposition on blanket bog. J. Ecol. 1978, 66, 631–650. [Google Scholar] [CrossRef]
- Yeloff, D.; Mauquoy, D. The influence of vegetation composition on peat humification: Implications for palaeoclimatic studies. Boreas 2006, 35, 662–673. [Google Scholar] [CrossRef]
- Buurman, P.; Nierop, K.G.J.; Pontevedra-Pombal, X.; Martínez-Cortizas, A. Molecular chemistry by pyrolysis-GC/MS of selected samples of the Penido Vello peat deposit, Galicia, NW Spain. In Peatlands: Basin Evolution and Repositories of Records of Environmental and Climate Changes; Martini, I.P., Martínez, A., Chesworth, W., Eds.; Elsevier: London, UK, 2006; pp. 219–242. [Google Scholar]
- Disnar, J.R.; Jacob, J.; Morched-Issa, M.; Lottier, N.; Arnaud, F. Assessment of peat quality by molecular and bulk geochemical analysis: Application to the Holocene record of the Chautagne marsh (Haute Savoie, France). Chem. Geol. 2008, 254, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.E.; Gallego, J.L.R.; Torres, T.; Díaz-Bautista, A.; Sierra, C. Palaeoenvironmental reconstruction of Northern Spain during the last 8000 cal yr BP based on the biomarker content of the Roñanzas peat bog (Asturias). Org. Geochem. 2010, 41, 454–466. [Google Scholar] [CrossRef]
- Schellekens, J.; Buurman, P.; Fraga, I.; Martínez-Cortizas, A. Holocene vegetation and hydrologic changes inferred from molecular vegetation markers in peat, Penido Vello (Galicia, Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 299, 56–69. [Google Scholar] [CrossRef]
- Schellekens, J.; Buurman, P.; Kuyper, T.W.; Abbott, G.D.; Pontevedra-Pombal, X.; Martínez-Cortizas, A. Influence of source vegetation and redox conditions on lignin-based decomposition proxies in graminoid-dominated ombrotrophic peat (Penido Vello, NWSpain). Geoderma 2015, 237, 270–282. [Google Scholar] [CrossRef]
- Schellekens, J.; Bradley, J.A.; Kuyper, T.W.; Fraga, I.; Pontevedra-Pombal, X.; Vidal-Torrado, P.; Abbott, G.D.; Peter Buurman, P. The use of plant-specific pyrolysis products as biomarkers in peat deposits. Quat. Sci. Rev. 2015, 123, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Moen, A.; Joosten, H.; Tanneberger, F. Mire diversity in Europe: Mire regionality. In Mires and Peatlands of Europe: Status, Distribution, and Nature Conservation; Joosten, H., Tanneberger, F., Moen, A., Eds.; Schweizerbart Science Publishers: Stuttgart, Germany, 2017; pp. 97–149. [Google Scholar]
- Katz, N.J.; Katz, S.V.; Kipiani, M.G. Atlas and Keys of Fruits and Seeds Occurring in the Quaternary Deposits of the USSR; Publishing House Nauka: Moscow, Russia, 1965; pp. 1–367. [Google Scholar]
- Katz, N.J.; Katz, S.V.; Skobeyeva, E.I. Atlas of Plant Remains in Peat Soil; Publishing House Nauka: Moscow, Russia, 1977; p. 736. [Google Scholar]
- Velichkevich, F.U.; Zastawniak, E. Atlas of the Pleistocene Vascular Plant Macrofossils of Central and Eastern Europe. Part 1: Pteridophytes and Monocotylendons; Polish Academy of Sciences, W Szafer Institute of Botany: Krakow, Poland, 2006. [Google Scholar]
- Velichkevich, F.U.; Zastawniak, E. Atlas of the Pleistocene Vascular Plant Macrofossils of Central and Eastern Europe. Part 2: Pteridophytes and Monocotylendons; Polish Academy of Sciences, W Szafer Institute of Botany: Krakow, Poland, 2008. [Google Scholar]
- Godwin, H. The History of the British Flora: A Factual Basis for Phytogeography; Cambridge University Press: Cambridge, UK, 1975. [Google Scholar]
- Grosse-Brauckmann, G. Über pflanzliche Makrofossilien mitteleuropäischer Torfe. I. Gewebereste krautiger Pflanzen und ihre Merkmale. Telma 1972, 2, 19–55. [Google Scholar]
- Grosse-Brauckmann, G. Über pflanzliche Makrofossilien mitteleuropäischer Torfe. II. Weitere Reste (Früchte und Samen, Moose u.a.) und ihre Bestimmungsmöglichkeiten. Telma 1974, 4, 51–117. [Google Scholar]
- Grosse-Brauckmann, G.; Streitz, B. Pflanzliche Makrofossilien mitteleuropäischer Torfe. III. Früchte, Samen undeinige Gewebe (Fotos von fossilen Pflanzenresten). Telma 1992, 22, 53–102. [Google Scholar]
- Mauquoy, D.; Van Geel, B. Plant macrofossil methods and studies: Mire and peat macros. In Encyclopedia of Quaternary Science; Elias, S.A., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2007; pp. 2315–2336. [Google Scholar]
- Berggren, G. Atlas of Seeds and Small Fruits of Northwest-European Plant Species, Part 2. Cyperaceae; Swedish Natural Science Research Council: Stockholm, Sweden, 1969; pp. 1–107. [Google Scholar]
- Körber-Grohne, U. Bestimmungsschlüssel für subfossile Juncus-samen und Gramineen-Früchtchen (Identification keys for subfossil Juncus seeds and grasses fruits). In Probleme der Küstenforschung im Südlichen Nordseegebiet (Problems of Coastal Research in the Southern North Sea Area); Haarnagel, W., Ed.; Lax: Hildesheim, Germany, 1964; Volume 1, pp. 1–47. [Google Scholar]
- Tallantire, P.A. Provisional key for the identification of sub-fossil seeds of Vaccinium spp. Folia Quat. 1976, 47, 39–40. [Google Scholar]
- Truchanowiczówna, J. Fossil seeds of the genus Menyanthes in Eurasia. Acta Paleobot. 1964, 5, 25–69. [Google Scholar]
- Bialobrzeska, M.; Truchanowiczówna, J. Zmiennose ksztaltu owoców i lusek europejskich brzóz (Betula L.) oraz oznaczanie ich w stanie kopalnym. (The variability of shape of fruits and scales of the European birches (Betula L.) and their determination in fossil materials.). Mon. Bot. 1964, 9, 1–93. [Google Scholar]
- Souto, M.; Castro, D.; Pontevedra-Pombal, X.; Garcia-Rodeja, E.; Fraga, M.I. Characterisation of Holocene plant macrofossils from North Spanish ombrotrofic mires: Vascular plants. Mires Peat 2016, 18, 1–21. [Google Scholar]
- Souto, M. Reconstrucción Paleoambiental de Turberas del Norte de la Península Ibérica Mediante Análisis de Macrofósiles Vegetales y Grado de Humificación de la Turba. Ph.D. Thesis, Universidade de Santiago de Compostela, Santiago, Spain, 2018; pp. 1–207. [Google Scholar]
- Birks, H.J.B. Quaternary bryophyte palaeoecology. In Bryophyte Ecology; Smith, A.J.E., Ed.; Chapman & Hall: London, UK, 1982; pp. 473–490. [Google Scholar]
- Barry, T.A.; Synnott, D.M. Further studies into bryophyte occurrence and succession in the Hochmoor peat types of Ireland. Glasra 1987, 10, 1–21. [Google Scholar]
- Tallis, J.H. Climate and erosion signals in British blanket peats: The significance of Racomitrium lanuginosum remains. J. Ecol. 1995, 83, 1021–1030. [Google Scholar] [CrossRef]
- Jakab, G.; Sümegi, P. The role of bryophyte paleoecology in quaternary climate reconstructions. In Bryophyte Ecology and Climate Change; Tuba, Z., Slack, N.G., Stark, Y.R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 335–358. [Google Scholar]
- Souto, M.; Castro, D.; Pontevedra-Pombal, X.; Garcia-Rodeja, E.; Fraga, M.I. Characterisation of Holocene plant macrofossils from North Spanish ombrotrofic mires: Bryophytes. Mires Peat 2017, 19, 1–12. [Google Scholar]
- Maldonado, F.J.; Roch, V.; Rubiales, J.M.; Morla, C.; García-Amorena, I. Caracterización y Significación de los Yacimientos Paleobotánicos (Paleoxilológicos) de la Sierra de Gredos. In Proceedings of the IV Congreso Forestal Nacional, Zaragoza, Spain, 26–30 September 2005. [Google Scholar]
- García-Antón, M.; Franco Múgica, F.; Maldonado Ruiz, J.; Morla Juaristi, C.; Sainz Ollero, H. Una secuencia polínica en Quintana Redonda (Soria). Evolución holocena del tapiz vegetal en el Sistema Ibérico Septentrional. An. Jard. Bot. 1995, 52, 187–195. [Google Scholar]
- Muñoz Sobrino, C.; Ramil Rego, P.; Delibes de Castro, G.; Rojo Guerra, M. Datos paleobotánicos sobre la turbera de la Piedra (Páramo de Tozo, Burgos). In Biogeografía Pleistocena-Holocena de la Península Ibérica; Ramil Rego, P., Fernández Rodríguez, C., Rodríguez Guitián, M., Eds.; Xunta de Galicia: Santiago, Spain, 1996; pp. 149–162. [Google Scholar]
- Roig, S.; Gómez Manzaneque, F.; Masedo, F.; Morla, C.; Sanchez Hernando, L.J. Estudio paleobotánico de estróbilos y maderas subfósiles holocenas en el yacimiento de Cevico Navero (Palencia, España). An. Jard. Bot. 1997, 55, 111–123. [Google Scholar]
- Alcalde, C.; García-Amorena, I.; Gómez Manzaneque, F.; Maldonado, J.; Morla, C.; Postigo Mijarra, J.M. Estudio de los macrorrestos vegetales del yacimiento de Lomilla (Aguilar de Campo, Palencia, España). An. Jard. Bot. 2000, 59, 101–112. [Google Scholar]
- García-Amorena, I.; Morla, C.; Rubiales, J.M.; Gómez Manzaneque, F. Taxonomic composition of the Holocene forests of the northern coast of Spain, as determined from their macroremains. Holocene 2008, 18, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, F. Late-quaternary deposits at le Moura, Biarritz, south-west France. New Phytol. 1964, 63, 374–409. [Google Scholar] [CrossRef]
- Castro, D.; Souto, M.; Garcia-Rodeja, E.; Pontevedra-Pombal, X.; Fraga, M.I. Climate change records between the mid and late Holocene in a peat bog from Serra do Xistral (SW Europe) using plant macrofossils and peat humification analyses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 420, 82–95. [Google Scholar] [CrossRef]
- Castro, D. Reconstrucción Paleoambiental das Turbeiras da Serra do Xistral Mediante Análises Paleobotánicas. Ph.D. Thesis, Universidade de Santiago de Compostela, Santiago, Spain, 2017; p. 207. [Google Scholar]
- Stefanini, B.S.; Oksanen, P.O.; Corcoran, J.P.; Mitchell, F.G. Appraising the cohesion of palaeoenvironmental reconstructions in north-west Spain since the mid-Holocene from a high temporal resolution multi-proxy peat record. Holocene 2018, 28, 681–694. [Google Scholar] [CrossRef]
- Pontevedra-Pombal, X.; Nóvoa-Muñoz, J.C.; García-Rodeja, E.; Martínez-Cortizas, A. Mountain mires from Galicia (NW Spain). In Peatlands: Basin Evolution and Repositories of Records of Environmental and Climate Changes; Martini, I.P., Martínez, A., Chesworth, W., Eds.; Elsevier: London, UK, 2006; pp. 83–108. [Google Scholar]
- Pontevedra-Pombal, X.; Castro, D.; Carballeira, R.; Souto, M.; López-Sáez, J.A.; Pérez-Díaz, S.; Fraga, M.I.; Valcárcel, M.; García-Rodeja, E. Iberian acid peatlands: Types, origin and general trends of development. Mires Peat 2017, 19, 1–19. [Google Scholar]
- Pontevedra-Pombal, X. Turberas de Montaña de Galicia. Génesis, Propiedades y su Aplicación Como Registros Ambientales Geoquímicos. Ph.D. Thesis, Universidade de Santiago de Compostela, Santiago, Spain, 2002; pp. 1–483. [Google Scholar]
- Heras, P.; Infante, M.; Pontevedra-Pombal, X.; Nóvoa-Muñoz, J.C. Mires and peatlands of Spain. In Mires and Peatlands of Europe: Status, Distribution, and Nature Conservation; Joosten, H., Tanneberger, F., Moen, A., Eds.; Schweizerbart Science Publishers: Stuttgart, Germany, 2017; pp. 639–656. [Google Scholar]
- Ponel, P.; de Beaulieu, J.L.; Tobolsk, K. Holocene palaeoenvironments at the timberline in the taillefer massif, French Alps: A study of pollen, plant macrofossils and fossil insects. Holocene 1992, 2, 117–130. [Google Scholar] [CrossRef]
- Ponel, P.; Court-Picon, M.; Badura, M.; Guiter, F.; de Beaulieu, J.L.; Andrieu-Ponel, V.; Djamali, M.; Leydet, M.; Gandouin, E.; Buttler, A. Holocene history of Lac des Lauzons (2180 m a.s.l.), reconstructed from multiproxy analyses of Coleoptera, plant macroremains and pollen (Hautes-Alpes, France). Holocene 2011, 21, 565–582. [Google Scholar] [CrossRef]
- Field, M.H.; de Beaulieu, J.L.; Guiot, J.; Ponel, P. Middle Pleistocene deposits at La Cote, Val-de-Lans, Isère department, France: Plant macrofossil, palynological and fossil insect investigations. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 159, 53–83. [Google Scholar] [CrossRef]
- Tobolski, K.; Ammann, B. Macrofossils as records of plant responses to rapid Late Glacial climatic changes at three sites in the Swiss Alps. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 159, 251–259. [Google Scholar] [CrossRef]
- Finsinger, W.; Tinner, W. Pollen and plant macrofossils at Lac de Fully (2135 m a.s.l.): Holocene forest dynamics on a highland plateau in the Valais, Switzerland. Holocene 2007, 17, 1119–1127. [Google Scholar] [CrossRef]
- Stähli, M.; Finsinger, W.; Tinner, W.; Allgöwer, B. Wildfire history and fire ecology of the Swiss National Park (Central Alps): New evidence from charcoal, polen and plant macrofossils. Holocene 2006, 16, 805–817. [Google Scholar] [CrossRef]
- Kalis, A.J.; van der Knaap, W.O.; Schweizer, A.; Urz, R. Three thousand year succession of plant communities on a valley bottom in the Vosges Mountains, NE France, reconstructed from fossil pollen, plant macrofossils, and modern phytosociological communities. Veget. Hist. Archaeobot. 2006, 15, 377–390. [Google Scholar] [CrossRef]
- Fauvart, N.; Ali, A.; Terral, J.F.; Roiron, P.; Blarquez, O.; Carcaillet, C. Holocene upper tree-limits of Pinus section sylvestris in the Western Alps as evidenced from travertine archives. Rev. Palaeobot. Palynol. 2012, 169, 96–102. [Google Scholar] [CrossRef]
- Ammann, B.; van der Knaap, W.O.; Lang, G.; Gaillard, M.J.; Kaltenrieder, P.; Rösch, M.; Finsinger, W.; Wright, H.E.; Tinner, W. The potential of stomata analysis in conifers to estimate presence of conifer trees: Examples from the Alps. Veget. Hist. Archaeobot. 2014, 23, 249–264. [Google Scholar] [CrossRef]
- Avigliano, R.; Anastasio, G.D.; Improta, S.; Peresani, M.; Ravazzi, C. A new late glacial to early Holocene palaeobotanical and archaeological record in the Eastern Pre-Alps: The Palughetto basin (Cansiglio Plateau, Italy). J. Quat. Sci. 2000, 15, 789–803. [Google Scholar] [CrossRef]
- Heiss, A.G.; Kofler, W.; Oeggl, K. The ulten valley in South Tyrol, Italy: Vegetation and settlement history of the area, and macrofossil record from the Iron Age cult site of St. Walburg. Palyno Bull. 2005, 1, 63–73. [Google Scholar]
- Krisai, R.; van Leeuwen, J.F.; van der Knaap, W.O. Present-day vegetation and the Holocene and recent development of Egelsee-Moor, Salzburg province, Austria. Veget. Hist. Archaeobot. 2016, 25, 555–568. [Google Scholar] [CrossRef]
- Sjögren, P.; Lamentowicz, M. Human and climatic impact on mires: A case study of Les Amburnex mire, Swiss Jura Mountains. Veget. Hist. Archaeobot. 2008, 17, 185–197. [Google Scholar] [CrossRef]
- Krisai, R. Zum rezenten und subfossilen Vorkommen subarktischer Moose im salzburgisch/oberösterreichischen Alpenvorland Vorh. Zool. Bot. Gos. Osterreich. 1985, 123, 143–150. [Google Scholar]
- Hölzer, A.; Hölzer, A. Ein torfprofil vom westabfall der hornisgride im nordschwarzwald mit Messia triquetra Angstr. Carolinea 2000, 58, 139–148. [Google Scholar]
- Mitchell, E.A.D.; Payne, R.J.; van der Knaap, W.O.; Lamentowicz, L.; Gąbka, M.; Lamentowicz, M. The performance of single- and multi-proxy transfer functions (testate amoebae, bryophytes, vascular plants) for reconstructing mire surface wetness and pH. Quat. Res. 2013, 79, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Miola, A.; Bondesan, A.; Corain, L.; Favaretto, S.; Mozzi, P.; Piovan, S.; Sostizzo, I. Wetlands in the Venetian Po Plain (northeastern Italy) during the Last Glacial Maximum: Interplay between vegetation, hydrology and sedimentary environment. Rev. Palaeobot. Palynol. 2006, 141, 53–81. [Google Scholar] [CrossRef]
- Miola, A.; Favaretto, S.; Sostizzo, I.; Valentini, G.; Asioli, A. Holocene salt marsh plant communities in the North Adriatic coastal plain (Italy) as reflected by pollen, non-pollen palynomorphs and plant macrofossil analyses. Veget. Hist. Archaeobot. 2010, 19, 513–529. [Google Scholar] [CrossRef]
- Zaccone, C.; Lobianco, D.; Shotyk, W.; Ciavatta, C.; Appleby, P.; Brugiapaglia, E.; Casella, L.; Miano, T.M.; D’Orazio, V. Highly anomalous accumulation rates of C and N recorded by a relic, free- floating peatland in Central Italy. Sci. Rep. 2017, 7, 43040. [Google Scholar] [CrossRef] [PubMed]
- Hájek, M.; Horsák, M.; Tichy, L.; Hájkova, P.; Díte, D.; Jamrichová, E. Testing a relict distributional pattern of fen plant and terrestrial snail species at the Holocene scale: A null model approach. J. Biogeogr. 2011, 38, 742–755. [Google Scholar] [CrossRef]
- Hájková, P.; Stechová, T.; Soltés, R.; Smerdová, E.; Plesková, Z.; Díte, D.; Bradácová, J.; Mútnanová, M.; Singh, P.; Hájek, M. Using a new database of plant macrofossils of the Czech and Slovak Republics to compare past and present distributions of hypothetically relict fen mosses. Preslia 2018, 90, 367–386. [Google Scholar] [CrossRef]
- Hájková, P.; Grootjans, A.B.; Lamentowicz, M.; Rybnickova, E.; Madaras, M.; Opravilova, V.; Hajkova, T. How a Sphagnum fuscum-dominated bog changed into a calcareous fen: The unique Holocene history of a Slovak spring-fed mire. J. Quat. Sci. 2012, 27, 233–243. [Google Scholar] [CrossRef]
- Peterka, T.; Hájek, M.; Dítě, D.; Hájková, P.; Palpurina, S.; Goia, I.; Grulich, V.; Kalníková, V.; Plesková, Z.; Šímová, A.; et al. Relict occurrences of boreal brown-moss quaking rich fens in the Carpathians and adjacent territories. Folia Geobot. 2018, 53, 265–276. [Google Scholar] [CrossRef]
- Kołaczek, P.; Gałka, M.; Apolinarska, K.; Płóciennik, M.; Gąsiorowski, M.; Brooks, S.J.; Hutchinson, S.M.; Karpińska-Kołaczek, M. A multi-proxy view of exceptionally early postglacial development of riparian woodlands with Ulmus in the Dniester River valley, western Ukraine. Rev. Palaeobot. Palynol. 2018, 250, 27–43. [Google Scholar] [CrossRef]
- Šímová, A.; Pánek, T.; Gałka, M.; Zernitskaya, V.; Hájková, P.; Brodská, H.; Jamrichova, E.; Hájek, M. Landslides increased Holocene habitat diversity on a flysch bedrock in the Western Carpathians. Quat. Sci. Rev. 2019, 219, 68–83. [Google Scholar] [CrossRef]
- Stachowicz-Rybka, R.; Gałka, M.; Alexandrowicz, W.P.; Alexandrowicz, S.W. Plant macrofossils and malacocoenoses of Quaternary mineral-organic sediments at Starunia palaeontological site and vicinity (Carpathian region, Ukraine). Ann. Soc. Geol. Pol. 2009, 79, 297–313. [Google Scholar]
- Gałka, M.; Tantau, I.; Ersek, V.; Feurdean, A. A 9000 year record of cyclic vegetation changes identified in a montane peatland deposit located in the Eastern Carpathians (Central-Eastern Europe): Autogenic succession or regional climatic influences? Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 449, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Magyari, E.; Sümegi, P.; Braun, M.; Jakab, G.; Molnar, M. Retarded wetland succession: Anthropogenic and climatic signals in a Holocene peat bog profile from north-east Hungary. J. Ecol. 2001, 89, 1019–1032. [Google Scholar] [CrossRef]
- Magyari, E.; Jakab, G.; Sümegi, P.; Szoör, G.Y. Holocene vegetation dynamics im the Bereg Plain, NE Hungary—The Báb-tava pollen and plant macrofossil record. Acta GGM Debrecina 2008, 3, 33–50. [Google Scholar]
- Sümegi, P.; Jakab, G.; Majkut, P.; Törocsik, T.; Zatykó, C. Middle age paleoecological and paleoclimatological reconstruction in the Carparthian Basin. Idöjárás 2009, 113, 265–298. [Google Scholar]
- Gałka, M.; Feurdean, A.; Hutchinson, S.; Milecka, K.; Tantau, I.; Apolinarska, K. Response of a spring-fed ecosystem in Central Eastern Europe (NW Romania) to climate changes during the last 4000 years: A high resolution multi-proxy reconstruction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 504, 170–185. [Google Scholar] [CrossRef]
- Bozilova, E.; Tonkov, S. Pollen from Lake Sedmo Rilsko reveals southeast European postglacial vegetation in the highest mountain area of the Balkans. New Phytol. 2000, 148, 315–325. [Google Scholar] [CrossRef]
- Marinova, E.; Tonkov, S. Holocene vegetation history of the northwestern Pirin Mountain (Bulgaria). Plant fossil record from peat-bog Mozgovitsa. C. R. Acad. Bulg. Sci. 2012, 65, 1087–1094. [Google Scholar]
- Tonkov, S.; Possnert, G.; Bazilova, E.; Marinova, E.; Pavlova, D. On the Holocene vegetation history of the Central Rila Mountains, Bulgaria: The palaeoecological record of peat bog Vodniza (2113 m). Rev. Palaeobot. Palynol. 2018, 250, 16–26. [Google Scholar] [CrossRef]
- Tonkov, S.; Marinova, E. Pollen and plant macrofossil analyses of mid-Holocene ra-diocarbon dated profiles from two subalpine lakes in Rila Mountains, Bulgaria. Holocene 2005, 15, 663–671. [Google Scholar] [CrossRef]
- Stefanova, I. New data on the late Holocene vegetative succession in the Northern Pirin Mts: Pollen and macrofossil analysis of depositions from peat bogs Goce Delchev and Visokata Ela. Phytol. Balc. 1997, 3, 53–63. [Google Scholar]
- Stefanova, I.; Atanassova, J.; Delcheva, M.; Wright, H.E. Chronological framework for the Lateglacial pollen and macrofossil sequence in the Pirin Mountains, Bulgaria: Lake Besbog and Lake Kremensko-5. Holocene 2006, 16, 877–892. [Google Scholar] [CrossRef]
- Lazarova, M.; Marinova, E.; Tonkov, S.; Snowball, I. A 5000-year pollen and plant macrofossil record from the Osogovo Mountain, Southwestern Bulgaria: Vegetation history and human impact. Rev. Palaeobot. Palynol. 2015, 223, 1–9. [Google Scholar] [CrossRef]
- Stefanova, I.; Ammann, B. Lateglacial and Holocene vegetation belts in the Pirin Mountains (southwestern Bulgaria). Holocene 2003, 13, 97–107. [Google Scholar] [CrossRef]
- Finsinger, W.; Morales-Molino, C.; Gałka, M.; Valsecchi, V.; Bojovic, S.; Tinner, W. Holocene vegetation and fire dynanics at Crveni Potok, a small mire in the Dinaric Alps (Tara National Park, Serbia). Quat. Sci. Rev. 2017, 167, 63–77. [Google Scholar] [CrossRef]
- Markgraf, V. Pollen dispersal in a mountain area. Grana 1989, 19, 127–146. [Google Scholar] [CrossRef]
- Chytrý, M.; Hennekens, S.M.; Jiménez-Alfaro, B.; Knollová, I.; Dengler, J.; Schaminée, J.H.J.; Aćić, S.; Agrillo, E.; Ambarlı, D.; Angelini, P.; et al. European vegetation archive (EVA): An integrated database of European vegetation plots. Appl. Veg. Sci. 2016, 19, 173–180. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/avsc.12191 (accessed on 8 October 2019). [CrossRef]
- Mucina, L.; B€ultmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Carni, A.; Sumberova, K.; Willner, W.; Dengler, J.; Garcia, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Pokorná, A.; Dreslerová, D.; Křivánková, D. Archaeobotanical Database of the Czech Republic, an Interim Report. IANSA 2011, 1, 49–53. [Google Scholar] [CrossRef]
- Rochefort, L.; Lode, E. Restoration of degraded boreal peatlands. In Boreal Peatland Ecosystems; Ecological Studies (Analysis and Synthesis); Wieder, R.K., Vitt, D.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 188. [Google Scholar]
- Schouten, M.G.C. (Ed.) Conservation and Restoration of Raised Bogs; Geological, Hydrological and Ecological Studies; Department of the Environment and Local Government Ireland, Geological Survey of Ireland: Staatsbosbeheer, The Netherlands, 2002.




{kind=link}
{kind=link}
{kind=link}
| SITE LOCATION | DEPTH (cm) | Yrs. Cal. BP | MATERIAL | ALT (m a.s.l.) | REF | AUTOR |
|---|---|---|---|---|---|---|
| WEST ZONE | ||||||
| Zalama, De Ordunte Mts. (ES) | 226 | 8000 | Peat | 1330 | 38 | Souto, 2018 |
| Borralleiras, Cabaleiros Mts. (ES) | 230 | 5500 | Peat | 600 | 38 | Souto, 2018 |
| Pena da Cadela, Serra do Xistral (ES) | 183 | 5500 | Peat | 970 | 51 | Castro et al., 2015 |
| Chan de Veiga Mol,Serra do Xistral (ES) | 845 | 8000 | Peat | 695 | 52 | Castro, 2017 |
| Pedrido, Serra do Xistral (ES) | 250 | 4750 | Peat | 770 | 53 | Stefanini et al., 2018 |
| Noja, Cantabric Coast (ES) | 100 | 3200–4600 | Coast peat deposit | 50 | 49 | García et al., 2008 |
| Merón, Cantabric Coast (ES) | 20 | 6000–7000 | Coast peat deposit | 50 | 49 | García et al., 2008 |
| Baldaio, Cantabric Coast (ES) | 30 | 870 | Coast peat deposit | 25 | 49 | García et al., 2008 |
| Le Moura, Cantabric Coast (FR) | 600 | 10,000 | Peat | 50 | Oldfield, 1964 | |
| CENTRAL ZONE | ||||||
| Grande Basse, Vosges Mts. (FR) | 200 | 3000 | Peat | 945 | 64 | Kalis et al., 2006 |
| Canard Ib, Taillefer Massif (FR) | 125 | 10,000 | Peat | 2200 | 58 | Ponel et al., 1992 |
| Lac Lauzons, Hautes-Alpes (FR) | 170 | 10,000 | Peat | 2180 | 59 | Ponel et al., 2011 |
| Egelsee-Moor Mire, Salzburg (AT) | 600 | 10,000 | Peat | 700 | 69 | Krisai et al., 2016 |
| Fuorn Mire, Fuorn Valley (CH) | 255 | 8500 | Peat-Gyttja | 1805 | 63 | Stahii et al., 2006 |
| Lac de Fully, Rhône Valley (CH) | 270 | 11,000 | Gyttja | 2135 | 62 | Finsinger and Tinner, 2007 |
| Gerzensee (CH) | 190–200 | 11,500 | Gyttja | 630 | 61 | Tobolski and Ammann, 2000 |
| Leysin (CH) | 320–350 | 10,850–12,050 | Gyttja | 1230 | 61 | Tobolski and Ammann, 2000 |
| Northern Black Forest (GR) | 80 | 2000 | Peat | 430 | 72 | Hölzer and Hólzer, 2000 |
| Palughetto, Lapisina Valley (IT) | 50 | 12,200 | Paleo peat | 1040 | 67 | Avigliano et al., 2000 |
| Totenmoos, South Tyrol (IT) | 800 | 15,000 | Peat | 1718 | 68 | Heiss et al., 2005 |
| Paludetto S2, Venetian plain (IT) | 500–1050 | 8700 | Peat | 50 | 75 | Miola et al., 2010 |
| Fiorentina, Venetian plain (IT) | 1100–1800 | 3000–17,000 | Peat-clay-silt | 50 | 74 | Miola et al., 2006 |
| La Rota, Posta Fibreno (IT) | 400 | 600 | Peat | 76 | Zaccone et al., 2017 | |
| EAST ZONE | ||||||
| Begbunar, Osogovo Mts (BG) | 105 | 5000 | Peat | 1800 | 95 | Lazarova et al., 2015 |
| Lake Ostrezko, Rila Mts (BG) | 200 | 6000 | Peat | 2340 | 92 | Tonkov and Marinova, 2005 |
| Lake Besbog,Pirin Mts (BG) | 380 | 15,000 | Peat-Gyttja | 2200 | 94 | Stefanova et al., 2006 |
| Vodniza, Rila Mts (BG) | 500 | 9500 | Peat-Gyttja | 2113 | 91 | Tonkov et al., 2018 |
| Tara, Mts. (RS) | 270 | 9000 | Peat | 1600 | 97 | Fisinger et al., 2017 |
| Nagymohos (HU) | 140–280 | 4000–7500 | Peat | 300 | 85 | Magyari et al., 2001 |
| Báb-tava, Bereg Plain, (HU) | 142–176 | 1000–2300 | Peat | 100 | 86 | Magyari et al., 2008 |
| Sirok Nyírjes-to, Mátra Mts, (HU) | 400 | 9000 | Peat | 250 m | 42 | Jakab & Sümegi, 2010 |
| Gärgäläu fen, Rodna Mts. (RO) | 150 | 9500 | Peat | 1810 | 11 | Feurdean et al., 2016 |
| Valea Morii, North western (RO) | 100 | 1100 | Peat | 640 | 88 | Galka et al., 2018 |
| Tăul Muced, Rodna Mts. (RO) | 500 | 9000 | Peat | 1360 | 84 | Galka et al., 2016 |
| Belanské Lú́ky, Tatra Mts. (SK) | 230 | 10,000 | Peat | 700 | 79 | Hájková et al., 2012 |
| Nádas Lake, Cserhát Mts. (HU) | 340 | 8000 | Peat-Clay | 360 | 87 | Sümegi et al., 2009 |
| Starunia, Carpathos (UA) | 300–500 | 12,000 | Peat-Clay | 400 | 83 | Stachowicz-Rybka et al., 2009 |
| REF. BIBLIOGRAPHY | [53] | [52] | [38] | [51] | [38] |
|---|---|---|---|---|---|
| MACROFOSSIL TAXA | |||||
| Betula pubescens | S | ||||
| Ericaceae | LF, F, W | LF, F, W | |||
| Erica tetralix | LF | LF, S | |||
| Erica mackaiana | LF, S | LF, S | LF, S | LF, S | |
| Calluna vulgaris | S | LF, S | LF, S | LF, S | LF, S |
| Andromeda polifolia | LF, S | ||||
| Daboecia cantabrica | S | S | |||
| Vaccinium myrtillus | LF | LF, W | |||
| Poaceae | LF | ||||
| Agrostis curtisii | LF,R | LF,R | |||
| Molinia caerulea | LF,R,F | LF,R,F | LF,R,F | LF,R,F | |
| Potentilla erecta | LF, F | F | LF, F | ||
| Drosera intermedia | S | ||||
| Drosera rotundifolia | S | S | S | S | |
| Narthecium ossifragum | S | S | S | ||
| Caltha palustris | S | S | |||
| Cyperaceae | LF | LF, R | LF, R | LF, R | |
| Eriophorum sp. | S, LF | LF, R, S | LF, R, S | LF, R, S | S, LF, R |
| Carex echinata | F | ||||
| Carex demissa | F | ||||
| Carex binervis | F | ||||
| Carex durieui | F | F | |||
| Rhynchospora alba | F | ||||
| Trichophorum sp | LF | ||||
| Luzula multiflora | S | ||||
| Juncus bulbosus | S | S | S | ||
| Juncus squarrosus | S | S | S | ||
| BRYOPHYTA | |||||
| Aulacomnium palustre | VP | VP | |||
| Calliergonella cuspidata | VP | ||||
| Campylopus sp | VP | VP | |||
| Dicranum scoparium | VP | ||||
| Hypnum cupressiforme | VP | VP | VP | VP | VP |
| Leucobryum juniperoideum | VP | VP | VP | ||
| Polytrichum sp. | VP | ||||
| Racomitrium lanuginosum | VP | VP | VP | ||
| Spagnum capillifolium | VP | ||||
| Spagnum tenellum | VP | VP | VP | ||
| Sphagnum acutifolia | VP | ||||
| Sphagnum compactum | VP | VP | |||
| Sphagnum cuspidatum | VP | ||||
| Sphagnum molle | VP | ||||
| Sphagnum papillosum | VP | VP | |||
| Sphagnum sec. acutifolia | VP | VP | VP | VP | |
| Sphagnum sec. cuspidata | VP | VP | |||
| Thuidium tamariscinum | VP | ||||
| Calypogeia sphagnicola | VP | ||||
| Odontochisma sphagni | VP | ||||
| FUNGI | |||||
| Cenoccocum geophilum | SCL | SCL | SCL |
| REF. BIBLIOGRAPHY | [50] | [64] | [58] | [69] | [63] | [62] | [61] | [61] |
|---|---|---|---|---|---|---|---|---|
| MACROFOSSIL TAXA | ||||||||
| Pinus sp. | ST | S | ||||||
| Pinus cembra | BS, NE | NE | NE, S, W | |||||
| Pinus mugo | W, NE | |||||||
| Pinus uncinata | NE, S | |||||||
| Pinus sylvestris | NE, BS,S | NE,BS,S | ||||||
| Picea abies | NE, S | NE, W, S | NE, ST | S | ||||
| Abies alba | NE | NE | ||||||
| Juniperus sp. | W | NE | ||||||
| Juniperus communis | NE, S | NE, W | ||||||
| Larix sp. | Ne | BS,BK | ||||||
| Larix decidua | NE, ST | NE, S, W | ||||||
| Salix sp. | BS | BS | ||||||
| Populus tremula | BS,F | BS,F | ||||||
| Alnus glutinosa | S, BS | S, W | ||||||
| Betula sp. | F, W | BS | S, W | |||||
| Betula alba | F | F, BS | F, BS | F | ||||
| Betula carpatica | F | |||||||
| Betula pendula | S, BS | F | ||||||
| Betula pubescens | F, BS | F | ||||||
| Betula nana | F | |||||||
| Sambucus racemosa | S | |||||||
| Fagus sylvatica | BS, W, F |
| COD. SITE LOCATION | [79] | [85] | [97] | [87] | [86] | [84] | [88] |
|---|---|---|---|---|---|---|---|
| MACROFOSSIL TAXA | |||||||
| Amblystegium kochii | VP | ||||||
| Amblystegium serpens | VP | ||||||
| Anomodon sp. | VP | ||||||
| Aulacomnium palustre | VP | VP | |||||
| Brachythecium mildeanum | VP | ||||||
| Bryum pseudotriquetum | VP | VP | |||||
| Calliergon giganteum | VP | ||||||
| Calliergonella cuspidate | VP | ||||||
| Campylium stellatum | VP | VP | |||||
| Campylopus sp. | VP | ||||||
| Depranocladus | VP | ||||||
| Depranocladus aduncus | VP | VP | |||||
| Herzogiella seligeri | VP | ||||||
| Leucodon sciuroides | VP | ||||||
| Meesia cf hexasticha | VP | ||||||
| Meesia longiseta | VP | VP | VP | VP | |||
| Meesia trinquetra | VP | ||||||
| Philonotis calcarean | VP | ||||||
| Polytrichum strictum | VP | VP | |||||
| Scorpidium cossonii | VP | ||||||
| Scorpidium revolvens | VP | ||||||
| Sphagnum sp. | VP | VP | VP | ||||
| Sphagnum acutifolia | VP | VP | |||||
| Sphagnum angustifolium | VP | ||||||
| Sphagnum cuspidatum | VP | ||||||
| Sphagnum fuscum | VP | ||||||
| Sphagnum magellanicum | VP | VP | |||||
| Sphagnum obtusum | VP | ||||||
| Sphagnum palustre | VP | VP | VP | VP | VP | ||
| Sphagnum sec. cuspidate | VP | VP | VP | VP | |||
| Sphagnum squarrosum | VP | VP | |||||
| Sphagnum subsecundum | VP | ||||||
| Thuidium recognitum | VP | ||||||
| Warnstorfia exannulata | VP | VP | VP | ||||
| Warnstorfia fluitans | VP |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souto, M.; Castro, D.; García-Rodeja, E.; Pontevedra-Pombal, X. The Use of Plant Macrofossils for Paleoenvironmental Reconstructions in Southern European Peatlands. Quaternary 2019, 2, 34. https://doi.org/10.3390/quat2040034
Souto M, Castro D, García-Rodeja E, Pontevedra-Pombal X. The Use of Plant Macrofossils for Paleoenvironmental Reconstructions in Southern European Peatlands. Quaternary. 2019; 2(4):34. https://doi.org/10.3390/quat2040034
Chicago/Turabian StyleSouto, Martin, Daniel Castro, Eduardo García-Rodeja, and Xabier Pontevedra-Pombal. 2019. "The Use of Plant Macrofossils for Paleoenvironmental Reconstructions in Southern European Peatlands" Quaternary 2, no. 4: 34. https://doi.org/10.3390/quat2040034
APA StyleSouto, M., Castro, D., García-Rodeja, E., & Pontevedra-Pombal, X. (2019). The Use of Plant Macrofossils for Paleoenvironmental Reconstructions in Southern European Peatlands. Quaternary, 2(4), 34. https://doi.org/10.3390/quat2040034