
Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications


Abstract
:1. Introduction
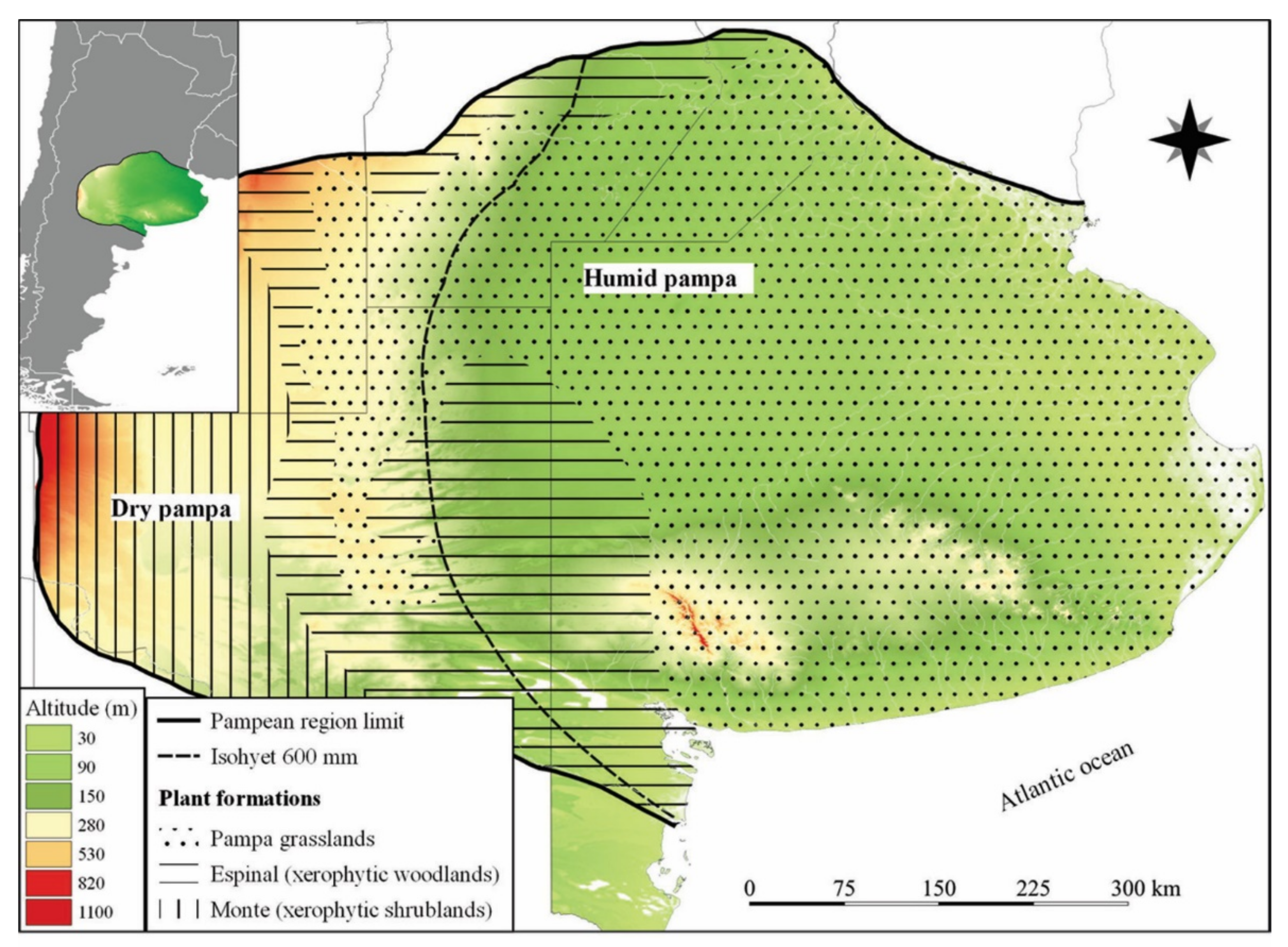
2. Characteristics of the Pampean Region
3. Geology
4. Paleoclimatic History and Mammal Diversity
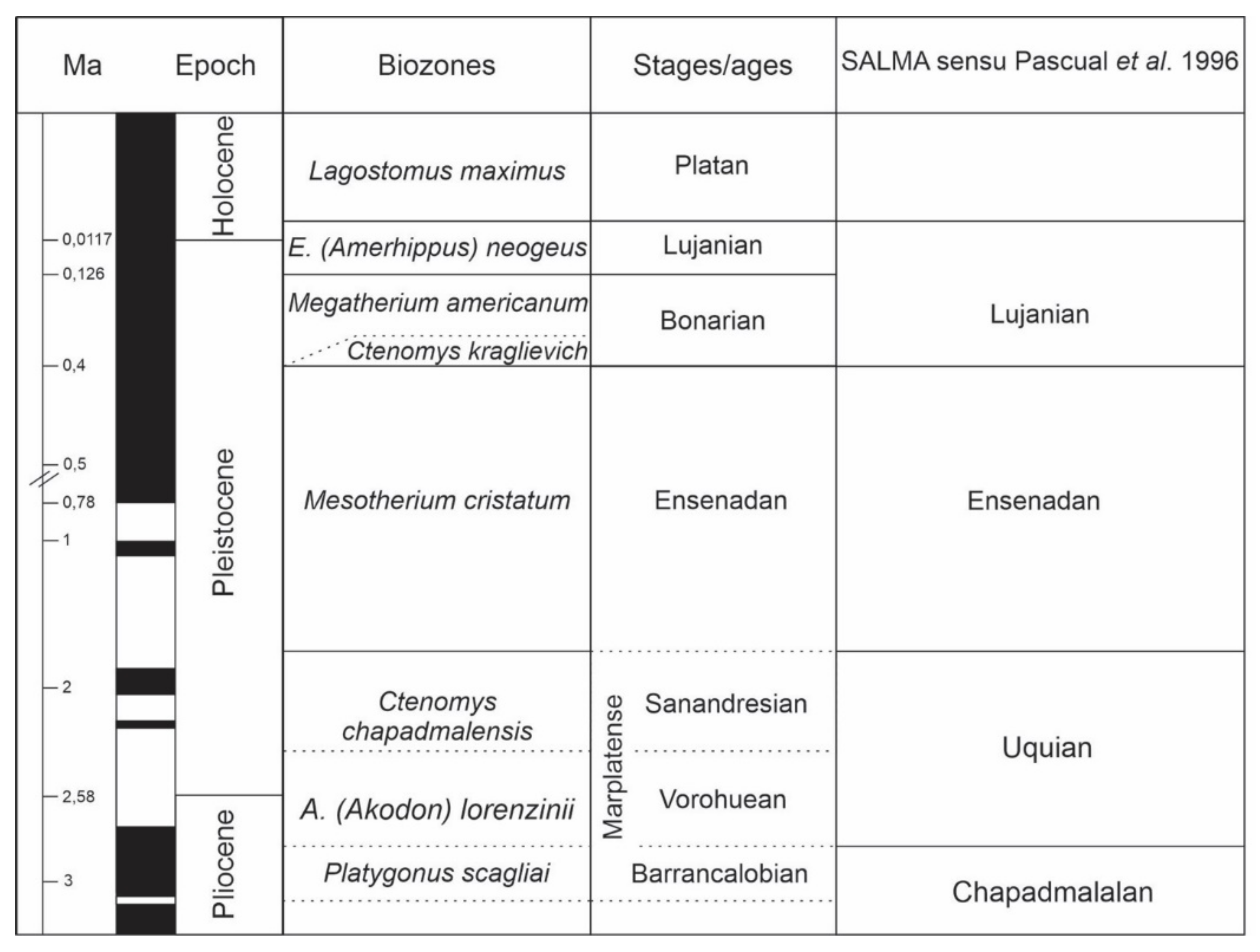
5. The Land Mammals Ages and Biostratigraphy
6. Mammal Biozone Scheme for the Pampas Region
6.1. Ctenomys chapalmalensis Biozone
6.2. Mesotherium cristatum Biozone
6.3. Megatherium americanum Biozone
6.4. Equus neogeus Biozone
7. Great American Biotic Interchange
8. Quaternary Megafauna Extinction
9. Final Remark
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- d’Orbigny, A. Voyage dans l’Amérique Méridionale (le Brésil, la République Orientale del’Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la République du Pérou), exécuté pendant les années 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833. Tome Troisième, 4° Partie: Paléontologie. Paris: P. Bertrand; Strasbourg: V. Levrault. 1842, p. 340. Available online: https://www.biodiversitylibrary.org/item/180098#page/7/mode/1up (accessed on 1 May 2021).
- Zárate, M.A.; Folguera, A.; Folguera, A. On the formations of the Pampas in the footsteps of Darwin: South of the Salado. Rev. Asoc. Geológica Argent. 2009, 64, 124–136. [Google Scholar]
- Ameghino, F. La Antigüedad del Hombre en el Plata 2; G. Masson: Paris, France; Igon Hermanos: Buenos Aires, Argentina, 1881; p. 640. [Google Scholar]
- Ameghino, F. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapalmalán. An. Mus. Nac. Buenos Aires 1908, 3, 343–428. [Google Scholar]
- Darwin, C.R. Journal of Researches into the Natural History and Geology of the Countries Visited during the Voyage of H.M.S. Beagle Round the World, under the Command of Capt. Fitz Roy, 2nd ed.; R.N. John Murray: London, UK, 1845; p. 520. [Google Scholar]
- Bravard, A. Geología de las Pampas; Registro Estadístico del Estado de Buenos Aires: Buenos Aires, Argentina, 1857; pp. 1–22. [Google Scholar]
- Soibelzon, E.; Soibelzon, L.H.; Gasparini, G.M.; Tonni, E.P. El Pleistoceno de la provincia de Buenos Aires y sus mamíferos. In Mioceno al Pleistoceno del centro y norte de Argentina; Nasif, N., Esteban, G., Chiesa, J., Zurita, A., Georgieff, S., Eds.; Opera Lilloana: Tucumán, Argentina, 2019; pp. 606–637. [Google Scholar]
- Pascual, R.; Ortega Hinojosa, E.J.; Gondar, D.; Tonni, E.P. Las edades del Cenozoico mamalífero de Argentina con especial atención a aquellas del territorio bonaerense. An. Com. Investig. Científica 1965, 6, 165–193. [Google Scholar]
- Tonni, E.P.; Alberdi, M.T.; Prado, J.L.; Bargo, M.S.; Cione, A.L. Changes of mammal assemblages in the pampean region (Argentina) and their relation with the Plio-Pleistocene boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1992, 95, 179–194. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P. Chronostratigraphy and ‘land mammal-ages’: The Uquian problem. J. Paleontol. 1995, 69, 135–159. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P. Inchasi, a Chapadmalalan (Pliocene) locality in Bolivia. Comments on the Pliocene-Pleistocene continental scale of southern South America. J. South. Am. Earth Sci. 1996, 9, 221–236. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P. Biostratigraphy and chronological scale of upper-most Cenozoic in the Pampean area, Argentina. In Quaternary Vertebrate Palaeontology in South America. Quaternary in South America and Antarctic Peninsula; Tonni, E.P., Cione, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 1999; Volume 12, pp. 23–52. [Google Scholar]
- Pardiñas, U.F.J.; Gelfo, J.; San Cristóbal, J.; Cione, A.L.; Tonni, E.P. Una asociación de organismos marinos y continentales en el Pleistoceno superior en el sur de la provincia de Buenos Aires, Argentina. XIII Congr. Geológico Argent. III Congr. Explor. Hidrocarb. 1996, 5, 95–112. [Google Scholar]
- Verzi, D.H.; Lezcano, M. Status sistemático y antigüedad de Megactenomys kraglievichi Rusconi, 1930 (Rodentia, Octodontidae). Rev. Mus. La Plata 1996, 9, 239–246. [Google Scholar]
- Verzi, D.H.; Deschamps, C.M.; Tonni, E.P. Biostratigraphic and paleoclimatic meaning of the Middle Pleistocene South American rodent Ctenomys kraglievichi (Caviomorpha, Octodontidae). Palaeogeogr. Palaeoclimatol. Palaeocology 2004, 212, 315–329. [Google Scholar] [CrossRef]
- Deschamps, C.M.; Tomassini, R.L. Late Cenozoic vertebrates from the southern Pampean Region: Systematic and bio-chronostratigraphic update. Publicación Electrónica Asoc. Paleontológica Argent. 2016, 16, 202–225. [Google Scholar] [CrossRef] [Green Version]
- Soibelzon, E.; Prevosti, F.J.; Bidegain, J.C.; Rico, Y.; Tonni, E.P.; Verzi, D. Correlation of cenozoic sequences of southeast Buenos Aires Province. Biostratigraphy and magnetostratigraphy. Quat. Int. 2009, 210, 51–56. [Google Scholar] [CrossRef]
- Burgos, J.J. El clima de la provincia de Buenos Aires en relación con la vegetación y el suelo. Flora de la provincia de Buenos Aires, parte 1. In Colección Científica; INTA: Buenos Aires, Argentina, 1969; pp. 33–100. [Google Scholar]
- Mancini, M.V.; Paez, M.M.; Prieto, A.R.; Stutz, S.; Tonello, M.; Vilanova, I. Mid-Holocene climatic variability reconstruction from pollen records (32°–52°S, Argentina). Quat. Int. 2005, 132, 47–59. [Google Scholar] [CrossRef]
- Prado, J.L.; Martinez-Maza, C.; Alberdi, M.T. Megafauna extinction in South America: A new chronology for the Argentine Pampas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 425, 41–49. [Google Scholar] [CrossRef]
- Ringuelet, R.A. Rasgos fundamentales de la zoogeografía de la Argentina. Phycis 1961, 22, 151–170. [Google Scholar]
- Cabrera, A.L. Fitogeografía de la República Argentina. Boletín Soc. Argent. Botánica 1971, 14, 1–42. [Google Scholar]
- Morrone, J.J. Biogeografía de América Latina y el Caribe. Man. Tesis Soc. Entomológica Aragonesa 2001, 3, 1–144. [Google Scholar]
- Cabrera, A.L. Regiones fitogeográficas argentinas. In Enciclopedia Argentina de Agricultura y Jardinería; T II. Editorial; ACME SACI: Buenos Aires, Argentina, 1976; p. 85. [Google Scholar]
- Zárate, M.A.; Tripaldi, A. The aeolian system of central Argentina. Aeolian Res. 2012, 3, 401–417. [Google Scholar] [CrossRef]
- Tonni, E.P.; Fidalgo, F. Consideraciones sobre los cambios climáticos durante el Pleistoceno tardio-reciente en la provincia de Buenos Aires. Aspectos ecológicos y zoogeográficos relacionados. Ameghiniana 1978, 15, 235–253. [Google Scholar]
- Iriondo, M.; Garcia, M.O. Climatic variations in the Argentine Plains during the last 18,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1993, 101, 201–220. [Google Scholar] [CrossRef]
- Irigoyen, M.R. Geología del Subsuelo y Plataforma Continental. In Proceedings of the Geología del Subsuelo y Plataforma Continental, Bahía Blanca, Argentina, 21–27 September 1975; pp. 139–169. [Google Scholar]
- Fidalgo, F.; de Francesco, O.; Colado, U. Geología superficial en las Hojas Castelli, J.M. Cobo y Monasterio (provincia de Buenos Aires). Relat. Congr. Geológico Argent. 1973, 4, 27–39. [Google Scholar]
- Fidalgo, F.; De Francesco, F.; Pacual, R. Geología superficial de la llanura bonaerense. Relat. Geol. Prov. Buenos Aires Sexto Congr. Geológico Argent. 1975, 103–138. [Google Scholar]
- Riggi, J.; Hidalgo, F.; Martínez, O.; Porro, N. Geología de los ‘‘Sedimentos Pampeanos’’ en el Partido de La Plata. Rev. Asoc. Geológica Argent. 1986, 41, 316–333. [Google Scholar]
- Tonni, E.P.; Nabel, P.; Cione, A.L.; Etchichury, M.; Tófalo, R.; Scillato Yané, G.; San Cristóbal, J.; Carlini, A.; Vargas, D. The Ensenada and Buenos Aires formations (Pleistocene) in a quarry near La Plata, Argentina. J. S. Am. Earth Sci. 1999, 12, 273–291. [Google Scholar] [CrossRef]
- Navel, P.; Cione, A.; Tonni, P. Environmental changes in the Pampean area of Argentina at the Matuyama–Brunhes (C1r–C1n) Chrons boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 162, 403–412. [Google Scholar]
- Zárate, M. El Cenozoico Tardío continental de la provincia de Buenos Aires. In 16 Congreso Geológico Argentino (La Plata); Relatorio: La Plata, Argentina, 2005; pp. 139–158. [Google Scholar]
- Kraglievich, J.L. El perfil geológico de Chapadmalal y Miramar, Provincia de Buenos Aires. Rev. Mus. Munic. Cienc. Nat. Tradic. 1952, 1, 1–8. [Google Scholar]
- González Bonorino, F. Mineralogía de las fracciones arcilla y limo del Pampeano en el área de la ciudad de Buenos Aires y su significado estratigráfico y sedimentológico. Rev. Asoc. Geológica Argent. 1965, 20, 67–148. [Google Scholar]
- Zárate, M.A. Loess of southern South America. Quat. Sci. Rev. 2003, 22, 1987–2006. [Google Scholar] [CrossRef]
- Rabassa, J.; Coronato, A.M.; Salemme, M.C. Chronology of the Late Cenozoic Patagonia glaciations and their correlation with biostratigraphic units of the pampean region (Argentina). J. S. Am. Earth Sci. 2005, 20, 81–103. [Google Scholar] [CrossRef]
- Fidalgo, F.; Riggi, J.C.; Gentile, R.; Correa, H.; Porro, N. Los ‘sedimentos post-pampeanos’ continentales en el ámbito sur bonaerense. Rev. Asoc. Geológica Argent. 1991, 46, 239–256. [Google Scholar]
- Cione, A.L.; Tonni, E.P. Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. Geología y Recursos Minerales de la Provincia de Buenos Aires. 16 Congreso Geológico Argentino. Relatorio 2005, 11, 183–200. [Google Scholar]
- Zárate, M.; Blasi, A. Late Pleistocene–Holocene Eolian deposits of the Southern Buenos Aires province, Argentina: A preliminary model. Quat. Int. 1993, 17, 15–20. [Google Scholar] [CrossRef]
- Zárate, M.; Blasi, A. Late Pleistocene and Holocene loess deposits of the southereastern Buenos Aires province, Argentina. Geojournal 1991, 24, 211–220. [Google Scholar] [CrossRef]
- Iriondo, M. Map of the South American plains. Its present state. Quat. S. Am. Antarct. Penins. 1988, 6, 297–308. [Google Scholar]
- García, N.O. South American climatology. Quat. Int. 1994, 21, 7–28. [Google Scholar] [CrossRef]
- Salazar Lea Plaza, J.C. Inventario Integrado de los Recursos Naturales de la provincia de La Pampa. Clima, Geomorfología, Suelo y Vegetación. INTA, provincia de La Pampa-UNL Pampa. Reg. Fisiográficas 1980, 98, 1–444. [Google Scholar]
- Gardenal, M. Geomorfología del partido de Saliquelló, provincia de Buenos Aires. CIC Unpubl. Rep. 1986, 60, 1. [Google Scholar]
- Prado, J.L.; Menegaz, A.; Tonni, E.P.; Salemme, M. Los mamíferos de la fauna local de Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectos paleoambientales y biostratigráficos. Ameghiniana 1987, 24, 217–233. [Google Scholar]
- Dillon, A.; Rabassa, J. Miembro La Chumbiada, Formación Luján (Pleistoceno, Provincia de BuenosAires), una nueva unidad estratigráfica del valle del río Salado. I J. Geol. Bonaer. 1985, 27, 1. [Google Scholar]
- Zárate, M.; Kemp, R.A.; Espinosa, M.; Ferrero, L. Pedosedimentary and palaeoenvironmental significance of a Holocene alluvial sequence in the southern Pampas, Argentina. Holocene 2000, 10, 481–488. [Google Scholar] [CrossRef]
- Isla, F.; Rutter, N.; Schnack, E.; Zárate, M. La transgresión Belgranense en Buenos Aires. Una revisión a cien años de su definición. Asoc. Geológica Argent. Serie D Publicación Espec. 2000, 4, 3–14. [Google Scholar]
- Aguirre, M.L.; Whatley, R.C. Late Quaternary marginal marine deposits and palaeoenvironments from northeastern Buenos Aires province, Argentina: A review. Quat. Sci. Rev. 1995, 14, 223–254. [Google Scholar] [CrossRef]
- Colado, U.; Figini, A.; Fidalgo, R.; Fucks, E. Los depósitos marinos del Cenozoico superior aflorantes en la zona comprendida entre Punta Indio y el Río Samborombón, provincia de Buenos Aires. Actas IV Jorn. Geol. Bonaer. 1995, 1, 151–158. [Google Scholar]
- Cione, A.L.; Tonni, E.P.; San Cristóbal, J.; Hernández, P.; Benítez, A.; Bordignon, F.; Perí, J. Putative meteoritic craters in Rıo Cuarto, Central Argentina interpreted as eolian landforms. Earth, Moon, and Planets. Int. J. Solar Syst. Sci. 2002, 91, 9–24. [Google Scholar]
- Martínez, S.; Ubilla, M.; Verde, M.; Perea, D.; Rojas, A.; Guérèquiz, R.; Pinñeiro, G. Paleoecology and Geochronology of Uruguayan Coastal Marine Pleistocene deposits. Quat. Res. 2001, 55, 246–254. [Google Scholar] [CrossRef]
- Suguio, K.; Martin, L. Quaternary marine formations of the State of Sao Paulo and Southern Rio de Janeiro. Int. Symp. Coast. Evol. Quat. 1978, 1, 1–55. [Google Scholar]
- Haq, B.U.; Hardenbol, J.; Vail, P.R. Chronology of fluctuation sea levels since the Triasic. Science 1987, 235, 1156–1167. [Google Scholar] [CrossRef] [Green Version]
- Shackleton, N.J. New Data on the Evolution of Pliocene Climatic Variability. In Paleoclimate and Evolution with Emphasis on Human Origins; Vrba, E.S., Denton, G.H., Partridge, T.C., Burckle, L.H., Eds.; Yale University Press: New Haven, CT, USA; London, UK, 1995; pp. 242–248. [Google Scholar]
- Rabassa, J. Cuaternario de la cordillera patagónica y Tierra del Fuego. In Anales del Instituto de Geología y Recursos Minerales; Caminos, R., Ed.; Geología Argentina: Buenos Aires, Argentina, 1999; pp. 710–714. [Google Scholar]
- Pascual, R.; Ortiz Jaureguizar, E. Evolving climates and mammal faunas in Cenozoic South America. In The Platyrrhine Fossil Record; Fleagle, J.G., Rosenberg, A.L., Eds.; Academic Press: London, UK, 1990; pp. 23–60. [Google Scholar]
- Pascual, R.; Ortiz Jaureguizar, E.; Prado, J.L. Land mammals: Paradigm for Cenozoic South American Geobiotic evolution. Münchner Geowissenchaftliche Abh. A 1996, 30, 265–320. [Google Scholar]
- Ortiz Jaureguizar, E.; Cladera, G. Paleoenvironmental evolution of southern South America during the Cenozoic. J. Arid Environ. 2006, 66, 489–532. [Google Scholar] [CrossRef]
- Marshall, L.G.; Cifelli, R. Analysis of changing diversity patterns in Cenozoic Land mammal age faunas, South America. Palaeovertebrata 1990, 19, 169–210. [Google Scholar]
- Strömberg, C.A.; Dunn, R.E.; Madden, R.H.; Kohn, M.J.; Carlini, A.A. Decoupling the spread of grasslands from the evolution of grazer-type herbivores in South America. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Pascual, R. Late Tertiary mammals of southern South America as indicators of climatic deterioration. Quat. S. Am. Antarct. Penins. 1984, 2, 1–30. [Google Scholar]
- Tonni, E.P.; Fidalgo, F. Geología y paleontología de los sedimentos del Pleistoceno en el área de Punta Hermengo (Miramar, Prov. de Buenos Aires, Rep. Argentina): Aspectos paleoclimáticos. Ameghiniana 1982, 19, 79–108. [Google Scholar]
- Fariña, R.A. Limb bone strength and habits in large glyptodonts. Lethaia 1995, 28, 189–196. [Google Scholar] [CrossRef]
- Croft, D.A. Cenozoic environmental change in South America as indicated by mammalian body size distributions (cenograms). Diversity Distrib. 2001, 7, 271–287. [Google Scholar] [CrossRef]
- Sánchez, B.; Prado, J.L.; Alberdi, M.T. Ancient feeding, ecology, and extinction of Pleistocene horses from the Pampean Region, Argentina. Ameghiniana 2006, 43, 427–436. [Google Scholar]
- Vucetich, M.G.; Verzi, D.H. First record of Dasyproctidae (Rodentia) in the Pleistocene of Argentina: Paleoclimatic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 178, 67–73. [Google Scholar] [CrossRef]
- Quattrocchio, M.E.; Borromei, A.M.; Deschampsc, C.M.; Grilla, S.C.; Zavala, C.A. Landscape evolution and climate changes in the Late Pleistocene–Holocene, southern Pampa (Argentina): Evidence from palynology, mammals and sedi- mentology. Quat. Int. 2008, 181, 123–138. [Google Scholar] [CrossRef]
- Vrba, E.S. Environment and evolution: Alternative causes of the temporal distribution of evolutionary events. S. Afr. J. Sci. 1985, 81, 229–236. [Google Scholar]
- Pardiñas, U.F.J.; Cione, A.L.; San Cristóbal, J.; Verzi, D.H.; Tonni, E.P. A new interglacial continental vertebrate assemblage in central-eastern Argentina. Curr. Res. Pleistocene 2004, 21, 111–112. [Google Scholar]
- Tonni, E.P.; Huarte, R.A.; Carbonari, J.E.; Figini, A.J. New radiocarbon chronology for the Guerrero Member of the Luján Formation (Buenos Aires, Argentina): Palaeoclimatic significance. Quat. Int. 2003, 109–110, 45–48. [Google Scholar]
- Bonadonna, F.P.; Leone, G.; Zanchetta, G. Composición isotópica de los fósiles de gasterópodos continentales de la provincia de Buenos Aires. Indicaciones paleoclimáticas. In Evolución Biológica y Climática de la Región Pampeana Durante los Últimos Cinco Millones de Años; Alberdi, M.T., Leone, G., Tonni, E.P., Eds.; Monografías del Museo Nacional de Madrid: Madrid, Spain, 1995; Volume 12, pp. 77–104. [Google Scholar]
- Ameghino, F. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas Acad. Nac. Cienc. 1889, VI, 1027. [Google Scholar]
- Ameghino, F. Le Diprothomo platensis: Un precurseur de l’homme du Pliocene inférieur de Buenos Aires. An. Mus. Nac. Hist. Nat. Buenos Aires 1909, 19, 107–209. [Google Scholar] [CrossRef]
- Frenguelli, J. Rasgos generales de la morfología y la geología de la Provincia de Buenos Aires. Publ. Lab. Ens. Mater. Investig. Tecnológicas 1950, 33, 1–72. [Google Scholar]
- Frenguelli, J. Argentina de Estudios geográficos GAEA. Neozoico 1957, 2–3, 218. [Google Scholar]
- Simpson, G.G. Review of the mammal-bearing tertiary of South America. Proc. Am. Philos. Soc. 1940, 83, 649–710. [Google Scholar]
- Wood, H.E., 2nd; Chaney, R.W.; Clark, J.; Colbert, E.H.; Jepsen, G.L.; Reeside, J.B., Jr. Committee. Nomenclature and correlation of the North American continental Tertiary. Bull. Geol. Soc. Am. 1941, 52, 1–48. [Google Scholar]
- Woodburne, M.O. Definition and characterization in mammalian chronostratigraphy. J. Paleontol. 1977, 51, 220–234. [Google Scholar]
- Archibald, J.D.; Clemens, W.A.; Gingerich, P.D.; Krause, D.K.; Lindsay, E.H.; Rose, K.D. First North American land-mammal ages of the Cenozoic Era. In Cenozoic Mammals of North America; Woodburne, M.O., Ed.; University of California Press: Berkeley, CA, USA, 1987; pp. 24–76. [Google Scholar]
- Pascual, R.; Ortega Hinojosa, E.J.; Gondar, D.; Tonni, E.P. Las edades del Cenozoico mamalífero de la provincia de Buenos Aires. In Paleontología Bonaerense; Borrello, A.V., Ed.; Comisión de Investigaciones Científicas de la Provincia de Buenos Aires: Buenos Aires, Argentina, 1966; pp. 3–12. [Google Scholar]
- Savage, D.E. Cenozoic geochronology of the fossil mammals of the Western Hemisphere. Rev. Mus. Argent. Cienc. Nat. 1962, 8, 53–67. [Google Scholar]
- Simpson, G.G. The evolution of marsupials in South America. Annais Acad. Bras. Ciências 1971, 43, 103–118. [Google Scholar]
- Cione, A.L.; Gasparini, G.M.; Soibelzon, E.; Soibelzon, L.H.; Tonni, E.P. The Great American Biotic Interchange. A South. American Perspective; Springer Brief Monographies in Earth System Sciences. South America and the Southern Hemisphere; Springer International Publishing AG: Cham, Switzerland, 2015; p. 97. [Google Scholar]
- Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.-X. The ICS International Chronostratigraphic Chart. Episodes 2013, 36, 199–204. [Google Scholar]
- Tonni, E.P. Los mamíferos del Cuaternario de la región pampeana de Buenos Aires, Argentina. In Quaternario do Rio Grande do Sul: Integrando Conhecimentos; Ribeiro, A.M., Girardi Bauermann, S., Saldanha Scherer, C., Eds.; Monografías da Sociedades Brasileira de Paleontologia: Porto Alegre, Brazil, 2009; pp. 207–216. [Google Scholar]
- Olivares, A.I.; Verzi, D.H.; Vucetich, M.G. Definición del género Eumysops Ameghino, 1888 (Rodentia, Echimyidae) y revisión de las especies del Plioce- no temprano de Argentina central. Ameghiniana 2012, 49, 198–216. [Google Scholar] [CrossRef]
- Merino, M.; Lutz, A.; Verzi, D.; Tonni, E.P. The fishing bat Noctilio (Mam- malia, Chiroptera) in the middle Pleistocene of central Argentina. Acta Chiropterologica 2007, 9, 401–407. [Google Scholar] [CrossRef]
- Deschamps, C.M. Late Cenozoic mammal bio-chronostratigraphy in south western Buenos Aires Province, Argentina. Ameghiniana 2005, 42, 733–750. [Google Scholar]
- Cione, A.L.; Tonni, E.P. Correlation of Pliocene to Holocene southern South American and European vertebrate-bearing units. Neogene and Quaternary Continental Stratigraphy and Mammal Evolution. Boll. Soc. Paleontol. Ital. 2001, 40, 167–173. [Google Scholar]
- Tonni, E.P.; Carbonari, J.E.; Huarte, R. Marine sediments attributed to marine isotope stage 3 in the southeastern Buenos Aires province, Argentina. Curr. Res. Pleistocene 2010, 27, 154–156. [Google Scholar]
- Gasparini, G.M.; Tonni, E.P. La fauna y los ambientes en el Cuaternario de la región Pampeana. Historia Evolutiva y Paleobiogeografía de los Vertebrados de América del Sur. XXX Jornadas Argentinas de Paleontología de Vertebrados, Volumen Especial. Contrib. Mus. Argent. Cienc. Nat. Bernardino Rivadavia 2016, 6, 395–402. [Google Scholar]
- Simpson, G.G. History of the fauna of Latin America. Am. Sci. 1950, 38, 261–389. [Google Scholar]
- Patterson, B.; Pascual, R. The Fossil Mammal Fauna of South America. In Evolution, Mammals and Southern Continents; Keast, A., Erk, F.C., Glass, B., Eds.; University of New York Press: Albany, NY, USA, 1972; pp. 247–309. [Google Scholar]
- Simpson, G.G. The Curious History of South American Mammals. In Splendid Isolation; Yale University Press: New Haven, CT, USA; London, UK, 1980; pp. 1–266. [Google Scholar]
- Webb, S.D. Late Cenozoic mammal dispersals between the Americas. In The Great American Biotic Interchange; Stehli, F.G., Webb, S.D., Eds.; Plenum Press: New York, NY, USA; London, UK, 1985; pp. 357–386. [Google Scholar]
- O’Dea, A.; Lessios, H.A.; Coates, A.G.; Eytan, R.I.; Restrepo-Moreno, S.A.; Cione, A.L.; Collins, L.S.; de Queiroz, A.; Farris, D.W.; Norris, R.D.; et al. Formation of the Isthmus of Panama. Sci. Adv. 2016, 2, e1600883. [Google Scholar] [CrossRef] [Green Version]
- Domingo, L.; Tomassini, R.L.; Montalvo, C.I.; Sanz-Pérez, D.; Alberdi, M.T. The Great American Biotic Interchange revisited: A new perspective from the stable isotope record of Argentine Pampas fossil mammals. Sci. Rep. 2020, 10, 1–10. [Google Scholar]
- Webb, S.D. Ecogeography and the Great American Interchange. Paleobiology 1991, 17, 266–280. [Google Scholar] [CrossRef]
- Webb, S.D. A history of savanna vertebrates in the New World. Part II: South America and the great interchange. Ann. Rev. Ecol. Syst. 1978, 9, 393–426. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Leone, G.; Tonni, E.P. Evolución Biológica y Climática de la Región Pampeana Durante los Últimos cinco Millones de años; Monografías del Museo Nacional de Madrid: Madrid, Spain, 1995; Volume 12, pp. 1–423. [Google Scholar]
- Prado, J.L.; Alberdi, M.T. Quaternary mammalian faunas of the Pampean Region. Quat. Int. 2010, 212, 176–186. [Google Scholar] [CrossRef]
- Reguero, M.A.; Candela, A.M.; Alonso, R.N. Biochronology and biostratigraphy of the Uquía Formation (Pliocene–early Pleistocene, NW Argentina) and its significance in the Great American Biotic Interchange. J. S. Am. Earth Sci. 2007, 23, 1–16. [Google Scholar] [CrossRef]
- Woodburne, M.O.; Cione, A.L.; Tonni, E.P. Central American provincialism and the Great American Biotic Interchange. In Advances in Late Tertiary Vertebrate Paleontology in Mexico and The Great American Biotic Interchange; Carranza-Castañeda, O., Lindsay, E.H., Eds.; Publicación Especial del Instituto de Geología y Centro de Geociencias de la Universidad Nacional Autónoma de México: Mexcio City, Mexico, 2006; Volume 4, pp. 73–101. [Google Scholar]
- Nieto, M.; Hortal, J.; Martínez-Maza, C.; Morales, J.; Ortiz-Jaureguizar, E.; Peláez-Campomanes, P.; Pickford, M.; Prado, J.L.; Rodríguez, J.; Senut, B.; et al. Historical determinants of mammal diversity in Africa: Evolution of mammalian body mass distribution in Africa and South America during Neogene and Quaternary times. In African Biodiversity. Molecules, Organisms, Ecosystems; Huber, B.A., Sinclair, B.J., Lampe, K.H., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 287–295. [Google Scholar]
- Cione, A.L.; Tonni, E.P.; Soibelzon, L.H. Did humans cause large mam- mal late Pleistocene-Holocene extinction in South America in a context of shrinking open areas. In American megafaunal extinctions at the end of the Pleistocene; Vertebrate Paleobiology and Paleontology Series; Haynes, G., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 125–144. [Google Scholar]
- Barnosky, A.D.; Lindsey, E.L.; Villavicencio, N.A.; Bostelmann, E.; Hadly, E.A.; Wanket, J.; Marshall, C.R. Variable impact of late-Quaternary megafaunal extinction in causing ecological state shifts in North and South America. Proc. Natl. Acad. Sci. USA 2016, 113, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnosky, A.D.; Lindsey, E.L. Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quat. Int. 2010, 217, 10–29. [Google Scholar] [CrossRef]
- Cione, A.L.; Tonni, E.P.; Soibelzon, L. The broken zig-zag: Late Cenozoic large mammal and turtle extinction in South America. Rev. Mus. Argent. Cienc. Nat. Bernardino Rivadavia 2003, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.S. Prehistorc Overkill: The Global Model. In Quaternary Extinction: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 354–403. [Google Scholar]
- Firestone, R.B.; West, A.; Kennett, J.P.; Becker, L.; Bunch, T.E.; Revay, Z.S.; Schultz, P.H.; Belgya, T.; Kennett, D.J.; Erlandson, J.M.; et al. Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and the Younger Dryas cooling. Proc. Natl. Acad. Sci. USA 2007, 104, 16016–16021. [Google Scholar] [CrossRef] [Green Version]
- Barnosky, A.D.; Bell, C.J.; Emslie, S.D.; Goodwin, H.T.; Mead, J.I.; Repenning, C.A.; Scott, E.; Shabel, A.B. Exceptional record of mid-Pleistocene vertebrates helps differentiate climatic from anthropogenic ecosystem perturbations. Proc. Natl. Acad. Sci. USA 2004, 101, 9297–9302. [Google Scholar] [CrossRef] [Green Version]
- Villavicencio, N.A.; Lindsey, E.L.; Martin, F.M.; Borrero, L.A.; Moreno, P.I.; Marshall, C.R.; Barnosky, A.D. Combination of humans, climate, and vegetation change triggered Late Quaternary megafauna extinction in the Última Esperanza region, southern Patagonia, Chile. Ecography 2016, 39, 125–140. [Google Scholar] [CrossRef]
- Steele, J.; Politis, G. AMS 14C dating of early human occupation of southern South America. J. Archaeol. Sci. 2009, 36, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Borrero, L.A. The elusive evidence: The archeological record of the South American extinct megafauna. In American Megafaunal Extinctions at the End of the Pleistocene; Haynes, G., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 145–168. [Google Scholar]
- Politis, G.G.; Prado, J.L.; Beukens, R.P. The human impact in Pleistocene–Holocene extinctions in South America. The Pampean case. In Ancient Peoples and Landscapes; Johnson, E., Ed.; Museum of Texas Tech University: Lubbock, TX, USA, 1995; pp. 187–205. [Google Scholar]
- Gutiérrez, M.; Martínez, G. Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean Region (Argentina). Quat. Int. 2008, 191, 53–68. [Google Scholar] [CrossRef]
- Prates, L.; Politis, G.; Steele, J. Radiocarbon chronology of the early human occupation of Argentina. Quat. Int. 2013, 301, 104–122. [Google Scholar] [CrossRef]
- Prates, L.; Politis, G.G.; Perez, S.I. Rapid radiation of humans in South America after the last glacial maximum: A radiocarbon-based study. PLoS ONE 2020, 15, e0236023. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.D. Megafauna biomass tradeoff as a driver of Quaternary and future extinctions. Proc. Natl. Acad. Sci. USA 2008, 105, 11543–11548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen-Smith, R.N. Pleistocene extinctions: The pivotal role of megaherbivores. Paleobiology 1987, 13, 351–362. [Google Scholar] [CrossRef]
- Graham, R.W.; Lundelius, E.L. Coevolutionary disequilibrium and Pleistocene extinctions. In Quaternary Extinction: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 223–249. [Google Scholar]
- Guthrie, R.D. Mosaics, allelochemics and nutrients. An ecological theory of late Pleistocene Megafaunal Extinction. In Quaternary Extinction: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; University of Arizona Press: Tucson, AZ, USA, 1984; pp. 259–298. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Doering (1882) | Ameghino (1908) | Kraglievich (1952) | Frenguelli (1957) | Fidalgo et al. (1973) | Riggi et al. (1986) | |
|---|---|---|---|---|---|---|
| Fm Querandina | Post pampeano | Platense | Fm de Loberia | Serie postpampeano | Fm La Postrera/Fm Luján | |
| Fm Tehuelche | Lujanense | Fm Buenos Aires | ||||
| Bonaerense | Fm de Arroyo Seco | Bonaerense | ||||
| Fm Pampeana | Ensenadense | Fm de Miramar | ||||
| Fm de San Andrés | Ensenadense | |||||
| Fm Pampeano | Fm Ensenada | |||||
| Fm de Vorohué | Fm Pampiano | |||||
| Fm de Barranca de los Lobos | ||||||
| Fm Araucano | Chapadmalense | Fm de Chapadmalal | Chapadmalense | |||
| Monte-hermosense | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prado, J.L.; Alberdi, M.T.; Bellinzoni, J. Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications. Quaternary 2021, 4, 15. https://doi.org/10.3390/quat4020015
Prado JL, Alberdi MT, Bellinzoni J. Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications. Quaternary. 2021; 4(2):15. https://doi.org/10.3390/quat4020015
Chicago/Turabian StylePrado, José Luis, María Teresa Alberdi, and Jonathan Bellinzoni. 2021. "Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications" Quaternary 4, no. 2: 15. https://doi.org/10.3390/quat4020015
APA StylePrado, J. L., Alberdi, M. T., & Bellinzoni, J. (2021). Pleistocene Mammals from Pampean Region (Argentina). Biostratigraphic, Biogeographic, and Environmental Implications. Quaternary, 4(2), 15. https://doi.org/10.3390/quat4020015