
‘Moving South’: Late Pleistocene Plant Exploitation and the Importance of Palm in the Colombian Amazon

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Theoretical Background
2. Geographical and Archaeological Background
2.1. Geographical Setting
2.2. Archaeological Setting
2.3. Study Region
3. Materials and Methods
4. Results
4.1. Chronology
4.2. Lithics
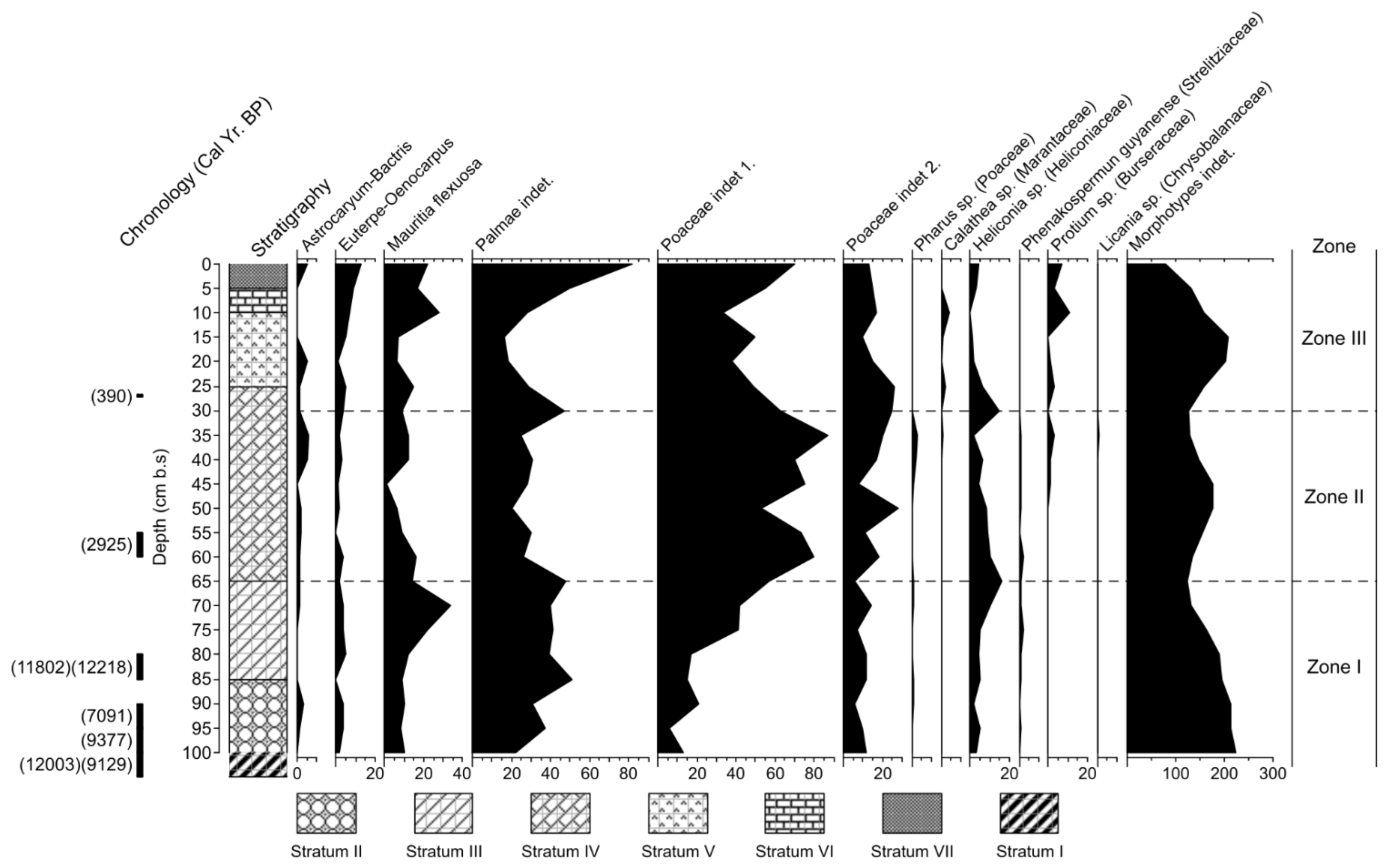
4.3. Plant Remains
4.4. Faunal Remains
5. Discussion
5.1. Diet
5.2. Palms
5.3. Ecological Knowledge
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ambrose, S.H. Paleolithic technology and human evolution. Science 2001, 291, 1748–1753. [Google Scholar] [CrossRef]
- Langley, M.C.; Amano, N.; Wedage, O.; Deraniyagala, S.; Pathmalal, M.; Perera, N.; Boivin, N.; Petraglia, M.D.; Roberts, P. Bows and arrows and complex symbolic displays 48,000 years ago in the South Asian tropics. Sci. Adv. 2020, 6, eaba3831. [Google Scholar] [CrossRef]
- Shea, J.J.; Sisk, M.L. Complex projectile technology and Homo sapiens dispersal into western Eurasia. PaleoAnthropology 2010, 2010, 100–122. [Google Scholar]
- Kennett, D.J.; Winterhalder, B. Behavioral Ecology and the Transition to Agriculture; University of California Press: Berkeley, CA, USA, 2019. [Google Scholar]
- Lombardo, U.; Iriarte, J.; Hilbert, L.; Ruiz-Pérez, J.; Capriles, J.M.; Veit, H. Early Holocene crop cultivation and landscape modification in Amazonia. Nature 2020, 581, 190–193. [Google Scholar] [CrossRef]
- Piperno, D.R. The origins of plant cultivation and domestication in the New World tropics: Patterns, process, and new developments. Curr. Anthropol. 2011, 52, S453–S470. [Google Scholar] [CrossRef]
- Piperno, D.R.; Pearsall, D.M. The Origins of Agriculture in the Lowland Neotropics; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Bailey, R.C.; Head, G.; Jenike, M.; Owen, B.; Rechtman, R.; Zechenter, E. Hunting and gathering in tropical rain forest: Is it possible? Am. Anthropol. 1989, 91, 59–82. [Google Scholar] [CrossRef]
- Headland, T.N. The wild yam question: How well could independent hunter-gatherers live in a tropical rain forest ecosystem? Hum. Ecol. 1987, 15, 463–491. [Google Scholar] [CrossRef]
- Jones, M. Moving north: Archaeobotanical evidence for plant diet in Middle and Upper Paleolithic Europe. Evol. Hominin Diets 2009, 171–180. [Google Scholar] [CrossRef]
- Erlandson, J.M.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Estes, J.A.; Steneck, R.S. The kelp highway hypothesis: Marine ecology, the coastal migration theory, and the peopling of the Americas. J. Isl. Coast. Archaeol. 2007, 2, 161–174. [Google Scholar] [CrossRef]
- Mellars, P.; Gori, K.C.; Carr, M.; Soares, P.A.; Richards, M.B. Genetic and archaeological perspectives on the initial modern human colonization of southern Asia. Proc. Natl. Acad. Sci. USA 2013, 110, 10699–10704. [Google Scholar] [CrossRef] [Green Version]
- Aceituno, F.J.; Loaiza, N. Early and middle Holocene evidence for plant use and cultivation in the middle Cauca river basin, Cordillera central (Colombia). Quat. Sci. Rev. 2014, 86, 49–62. [Google Scholar] [CrossRef]
- Aceituno, F.J.; Loaiza, N. The role of plants in the early human settlement of Northwest South America. Quat. Int. 2015, 363, 20–27. [Google Scholar] [CrossRef]
- Aceituno, F.J.; Loaiza, N. The origins and early development of plant food production and farming in Colombian tropical forests. J. Anthropol. Archaeol. 2018, 49, 161–172. [Google Scholar] [CrossRef]
- Gnecco, C. Against ecological reductionism: Late Pleistocene hunter-gatherers in the tropical forests of northern South America. Quat. Int. 2003, 109, 13–21. [Google Scholar] [CrossRef]
- Gnecco, C.; Mora, S. Late pleistocene/early holocene tropical forest occupations at San Isidro and Pena Roja, Colombia. Antiquity 1997, 71, 683. [Google Scholar] [CrossRef]
- Morcote-Ríos, G.; Aceituno, F.J.; Iriarte, J.; Robinson, M.; Chaparro-Cárdenas, J.L. Colonisation and early peopling of the Colombian Amazon during the late pleistocene and the early holocene: New evidence from La serranía La Lindosa. Quat. Int. 2021, 578, 5–19. [Google Scholar] [CrossRef]
- Bryan, A.L.; Casamiquela, R.M.; Cruxent, J.M.; Gruhn, R.; Ochsenius, C. An El Jobo Mastodon Kill at Taima-Taima, Venezuela. Science 1978, 200, 1275–1277. [Google Scholar] [CrossRef]
- Correal-Urrego, G. Exploraciones Arqueológicas en la costa Atlántica y valle del Magdalena: Sitios precerámicos y tipologías líticas. Caldasia 1977, 11, 33–128. [Google Scholar]
- Dickau, R.; Aceituno, F.J.; Loaiza, N.; López, C.; Cano, M.; Herrera, L.; Restrepo, C.; Ranere, A.J. Radiocarbon chronology of terminal Pleistocene to middle Holocene human occupation in the Middle Cauca Valley, Colombia. Quat. Int. 2015, 363, 43–54. [Google Scholar] [CrossRef]
- Hurt, W.R.; Van Der Hammen, T.; Urrego, G.C. Preceramic sequences in the El Abra rock-shelters, Colombia. Science 1972, 175, 1106–1108. [Google Scholar] [CrossRef]
- Mora, S.; Camargo, S.M. Early Inhabitants of the Amazonian Tropical Rain Forest: A Study of Humans and Environmental Dynamics; Center for Comparative Arch: Pittsburgh, PA, USA, 2003; Volume 3. [Google Scholar]
- Ranere, A.J.; López, C.E. Cultural diversity in late Pleistocene/early Holocene populations in northwest South America and lower Central America. Int. J. S. Am. Archaeol. 2007, 1, 25–31. [Google Scholar]
- Roosevelt, A.C.; Da Costa, M.L.; Machado, C.L.; Michab, M.; Mercier, N.; Valladas, H.; Feathers, J.; Barnett, W.; Da Silveira, M.I.; Henderson, A.; et al. Paleoindian cave dwellers in the Amazon: The peopling of the Americas. Science 1996, 272, 373–384. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Rossen, J. Plant food and its implications for the peopling of the New World: A view from South America. In The First Americans: The Pleistocene Colonization of the New World; California Academy of Sciences: San Francisco, CA, USA, 2002; pp. 237–253. [Google Scholar]
- Prous, A.; Fogaça, E. Archaeology of the Pleistocene-Holocene boundary in Brazil. Quat. Int. 1999, 53, 21–41. [Google Scholar] [CrossRef]
- Akibode, C.S.; Maredia, M.K. Global and regional trends in production, trade and consumption of food legume crops. Economics 2012. [Google Scholar] [CrossRef]
- Stahl, A.B.; Dunbar, R.I.M.; Homewood, K.; Ikawa-Smith, F.; Kortlandt, A.; McGrew, W.C.; Milton, K.; Paterson, J.D.; Poirier, F.E.; Sugardjito, J.; et al. Hominid dietary selection before fire [and Comments and Reply]. Curr. Anthropol. 1984, 25, 151–168. [Google Scholar] [CrossRef]
- Bailey, R.C.; Headland, T.N. The tropical rain forest: Is it a productive environment for human foragers? Hum. Ecol. 1991, 19, 261–285. [Google Scholar] [CrossRef]
- Headland, T.N.; Bailey, R.C. Introduction: Have hunter-gatherers ever lived in tropical rain forest independently of agriculture? Hum. Ecol. 1991, 19, 115–122. [Google Scholar] [CrossRef]
- Milton, K. Protein and carbohydrate resources of the Maku Indians of northwestern Amazonia. Am. Anthropol. 1984, 86, 7–27. [Google Scholar] [CrossRef]
- Hart, T.B.; Hart, J.A. The ecological basis of hunter-gatherer subsistence in African rain forests: The Mbuti of Eastern Zaire. Hum. Ecol. 1986, 14, 29–55. [Google Scholar] [CrossRef]
- Roberts, P.; Boivin, N.; Lee-Thorp, J.; Petraglia, M.; Stock, J. Tropical forests and the genus Homo. Evol. Anthropol. Issues News Rev. 2016, 25, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Dillehay, T.D. Climate and human migrations. Science 2002, 298, 764–765. [Google Scholar] [CrossRef]
- Gamble, C.; Davies, W.; Pettitt, P.; Richards, M. Climate change and evolving human diversity in Europe during the last glacial. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Kuper, R.; Kröpelin, S. Climate-controlled Holocene occupation in the Sahara: Motor of Africa’s evolution. Science 2006, 313, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Núñez, L.; Grosjean, M.; Cartajena, I. Human occupations and climate change in the Puna de Atacama, Chile. Science 2002, 298, 821–824. [Google Scholar] [CrossRef]
- Van Der Hammen, T.; Hooghiemstra, H. The el Abra stadial, a Younger Dryas equivalent in Colombia. Quat. Sci. Rev. 1995, 14, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, X.; Chiessi, C.; Mulitza, S.; Zhang, X.; Lohmann, G.; Prange, M.; Behling, H.; Zabel, M.; Govin, A.; et al. Equatorial Pacific forcing of western Amazonian precipitation during Heinrich Stadial 1. Sci. Rep. 2016, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groot, M.; Bogotá, R.G.; Lourens, L.J.; Hooghiemstra, H.; Vriend, M.; Berrio, J.C.; Tuenter, E.; Van Der Plicht, J.; Van Geel, B.; Ziegler, M.; et al. Ultra-high resolution pollen record from the northern Andes reveals rapid shifts in montane climates within the last two glacial cycles. Clim. Past 2011, 7, 299–316. [Google Scholar] [CrossRef] [Green Version]
- Ludt, W.B.; Rocha, L.A. Shifting seas: The impacts of Pleistocene sea-level fluctuations on the evolution of tropical marine taxa. J. Biogeogr. 2015, 42, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Bogotá-A, R.; Hooghiemstra, H.; Berrio, J. North Andean environmental and climatic change at orbital to submillennial time-scales: Vegetation, water-levels and sedimentary regimes from Lake Fúquene between 284 and 130 ka. Rev. Palaeobot. Palynol. 2016, 226, 91–107. [Google Scholar] [CrossRef]
- Leite, Y.L.; Costa, L.P.; Loss, A.C.; Rocha, R.G.; Filho, H.B.; Bastos, A.C.; Quaresma, V.; Fagundes, V.; Paresque, R.; Passamani, M.; et al. Neotropical forest expansion during the last glacial period challenges refuge hypothesis. Proc. Natl. Acad. Sci. USA 2016, 113, 1008–1013. [Google Scholar] [CrossRef] [Green Version]
- Barnosky, A.D.; Lindsey, E.L. Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quat. Int. 2010, 217, 10–29. [Google Scholar] [CrossRef]
- Cooke, R.; Ranere, A.; Pearson, G.; Dickau, R. Radiocarbon chronology of early human settlement on the Isthmus of Panama (13,000–7000 BP) with comments on cultural affinities, environments, subsistence, and technological change. Quat. Int. 2013, 301, 3–22. [Google Scholar] [CrossRef]
- Piperno, D.R.; Bush, M.B.; Colinvaux, P.A. Paleoecological perspectives on human adaptation in central Panama. I. The Pleistocene. Geoarchaeology 1991, 6, 201–226. [Google Scholar] [CrossRef]
- Cooke, R. Prehistory of native Americans on the Central American land bridge: Colonization, dispersal, and divergence. J. Archaeol. Res. 2005, 13, 129–187. [Google Scholar] [CrossRef]
- Dickau, R. Integrating Zooarchaeology and Paleoethnobotany; Springer: New York, NY, USA, 2010; pp. 99–134. [Google Scholar]
- Kistler, L.; Maezumi, S.Y.; De Souza, J.G.; Przelomska, N.A.S.; Costa, F.M.; Smith, O.; Loiselle, H.; Ramos-Madrigal, J.; Wales, N.; Ribeiro, E.R.; et al. Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science 2018, 362, 1309–1313. [Google Scholar] [CrossRef] [Green Version]
- Posth, C.; Nakatsuka, N.; Lazaridis, I.; Skoglund, P.; Mallick, S.; Lamnidis, T.C.; Rohland, N.; Nägele, K.; Adamski, N.; Bertolini, E.; et al. Reconstructing the deep population history of Central and South America. Cell 2018, 175, 1185–1197.e22. [Google Scholar] [CrossRef] [Green Version]
- Prufer, K.M.; Alsgaard, A.V.; Robinson, M.; Meredith, C.R.; Culleton, B.J.; Dennehy, T.; Magee, S.; Huckell, B.B.; Stemp, W.J.; Awe, J.J.; et al. Linking late Paleoindian stone tool technologies and populations in North, Central and South America. PLoS ONE 2019, 14, e0219812. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Escobar, O.A.; Lucas, E.; Jaramillo, C.; Monro, A.; Morris, S.K.; Bogarín, D.; Greer, D.; Dodsworth, S.; Aguilar-Cano, J.; Meseguer, A.S.; et al. The origin and diversification of the hyperdiverse flora in the Chocó biogeographic region. Front. Plant Sci. 2019, 10, 1328. [Google Scholar] [CrossRef]
- Gentry, A.H. Species richness and floristic composition of Chocó region plant communities. Caldasia 1986, 15, 71–91. [Google Scholar]
- Behling, H.; Hooghiemstra, H.; Negret, A.J. Holocene history of the Chocó rain forest from Laguna Piusbi, Southern Pacific lowlands of Colombia. Quat. Res. 1998, 50, 300–308. [Google Scholar] [CrossRef]
- Poveda, I.C.; Rojas, C.A.; Rudas, A.; Rangel, O. El Chocó biogeográfico: Ambiente físico. In Colombia Diversidad Biótica IV, El Chocó Biogeográfico/Costa Pacífica; Instituto de Ciencias Naturales, Conservación Internacional: Bogotá, Colombia, 2004; pp. 1–12. [Google Scholar]
- Forero, E.; Gentry, A.H. Lista Anotada de las Plantas del Departamento del Chocó, Colombia; Instituto de Ciencias Naturales, Museo de Historia Natural: Bogotá, Colombia, 1989. [Google Scholar]
- Berrío, J.C.; Behling, H.; Hooghiemstra, H. Tropical rain-forest history from the Colombian Pacific area: A 4200-year pollen record from Laguna Jotaordó. Holocene 2000, 10, 749–756. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Ramirez, C.; Pino, M.; Collins, M.B.; Rossen, J.; Pino-Navarro, J.D. Monte Verde: Seaweed, food, medicine, and the peopling of South America. Science 2008, 320, 784–786. [Google Scholar] [CrossRef] [Green Version]
- Gassón, R.A. Orinoquia: The archaeology of the Orinoco River Basin. J. World Prehistory 2002, 16, 237–311. [Google Scholar] [CrossRef]
- Hooghiemstra, H.; Flantua, S. Colombia in the Quaternary: An overview of environmental and climatic change. In The Geology of Colombia; Servicio Geológico Colombiano: Bogotá, Colombia, 2019. [Google Scholar]
- Rangel-Ch, J.O.; Lowy, P.; Aguilar, M. Distribución de los tipos de vegetación en las regiones naturales de Colombia. Aproximación inicial. In Diversidad Biótica II. Tipos de Vegetación en Colombia; Universidad Nacional de Colombia-Instituto de Ciencias Naturales: Bogotá, Colombia, 1997; pp. 403–436. [Google Scholar]
- Behling, H.; Hooghiemstra, H. Late Quaternary palaeoecology and palaeoclimatology from pollen records of the savannas of the Llanos Orientales in Colombia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 139, 251–267. [Google Scholar] [CrossRef]
- Berrio, J.C.; Hooghiemstra, H.; Behling, H.; Botero, P.; Van der Borg, K. Late-Quaternary savanna history of the Colombian Llanos Orientales from Lagunas Chenevo and Mozambique: A transect synthesis. Holocene 2002, 12, 35–48. [Google Scholar]
- Behling, H.; Hooghiemstra, H. Environmental history of the Colombian savannas of the Llanos Orientales since the Last Glacial Maximum from lake records El Pinal and Carimagua. J. Paleolimnol. 1999, 21, 461–476. [Google Scholar] [CrossRef] [Green Version]
- Berrío, J.C.; Hooghiemstra, H.; Marchant, R.; Rangel, O. Late-glacial and Holocene history of the dry forest area in the south Colombian Cauca Valley. J. Quat. Sci. Publ. Quat. Res. Assoc. 2002, 17, 667–682. [Google Scholar] [CrossRef]
- Marchant, R.; Behling, H.; Berrio, J.C.; Cleef, A.; Duivenvoorden, J.; Hooghiemstra, H.; Kuhry, P.; Melief, B.; Schreve-Brinkman, E.; Van Geel, B.; et al. Pollen-based biome reconstructions for Colombia at 3000, 6000, 9000, 12,000, 15,000 and 18,000 14C yr ago: Late Quaternary tropical vegetation dynamics. J. Quat. Sci. Publ. Quat. Res. Assoc. 2002, 17, 113–129. [Google Scholar]
- Marchant, R.; Behling, H.; Berrio, J.C.; Cleef, A.; Duivenvoorden, J.; Hooghiemstra, H.; Kuhry, P.; Melief, B.; Van Geel, B.; Van der Hammen, T.; et al. Mid-to Late-Holocene pollen-based biome reconstructions for Colombia. Quat. Sci. Rev. 2001, 20, 1289–1308. [Google Scholar] [CrossRef]
- Van der Hammen, T.; González, E. Upper pleistocene and holocene climate and vegetation of the “Sabana de Bogota”(Colombia, South America). Leidse Geol. Meded. 1960, 25, 261–315. [Google Scholar]
- Vélez, M.; Hooghiemstra, H.; Metcalfe, S.; Wille, M.; Berrío, J. Late Glacial and Holocene environmental and climatic changes from a limnological transect through Colombia, northern South America. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 234, 81–96. [Google Scholar] [CrossRef]
- Correal Urrego, G. Nuevas Evidencias Culturales Pleistocénicas y Megafauna en Colombia. Bol. Arqueol. 1993, 8, 3–12. [Google Scholar]
- Van der Hammen, T.; Urrego, G.C. Mastodontes en el humedal pleistocénico en el valle del Magdalena (Colombia) con evidencias de la presencia del hombre en el pleniglacial. Boletín de Arqueología 2001, 16, 1. [Google Scholar]
- Aceituno, F.J.; Rojas-Mora, S. Lithic technology studies in Colombia during the late Pleistocene and early Holocene. Chungara Rev. Antropol. Chil. 2015, 47, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Correal Urrego, G. Evidencias Culturales y Megafauna Pleistocénica en Colombia; Fundación de Investigaciones Arqueológicas, Banco de la Republica: Bogotá, Colombia, 1981. [Google Scholar]
- Correal, G. Apuntes sobre el medio ambiente pleistocénico y el hombre prehistórico en Colombia. In New Evidence for the Pleistocene Peopling of the Americas; University of Maine: Orano, ME, USA, 1986; pp. 115–131. [Google Scholar]
- Muttillo, B.; Lembo, G.; Rufo, E.; Peretto, C.; Pérez, R.L. Revisiting the oldest known lithic assemblages of Colombia: A review of data from El Abra and Tibitó (Cundiboyacense Plateau, Eastern Cordillera, Colombia). J. Archaeol. Sci. Rep. 2017, 13, 455–465. [Google Scholar] [CrossRef]
- Correal-Urrego, G. Restos de megafauna asociados a artefactos en la Sabana de Bogotá. Caldasia 1982, 13, 487–547. [Google Scholar]
- Arroyave, V.; Herrera, L.; López, C. Tecnología, forma y función de instrumentos bifaciales multiuso enmangables (IBME) del Aeropuerto del Café (Palestina, Caldas, Colombia). Int. J. S. Am. Archaeol. 2018, 12, 26–43. [Google Scholar]
- Aceituno, F.J. Entre el Río y la Montaña: Nuevos Datos Para el Poblamiento Temprano del Cauca Medio Colombiano; Fondo Editorial FCSH: Medellin, Colombia, 2019. [Google Scholar]
- Gnecco, C. Ocupación Temprana de Bosques Tropicales de Montaña; Universidad del Cauca: Cauca, Colombia, 2000. [Google Scholar]
- Cavelier, I.; Rodríguez, C.; Herrera, L.F.; Morcote, G.; Mora, S. No sólo de caza vive el hombre: Ocupación del bosque amazónico, Holoceno temprano. In Ambito y Ocupaciones Tempranas de la América Tropical; Fundación Erigaie: Bogotá, Colombia, 1995; pp. 27–44. [Google Scholar]
- Morcote, G.; Aceituno, F.; León, T. Recolectores del Holoceno temprano en la floresta amazónica colombiana. In Antes de Orellana. Actas del 3er Encuentro Internacional de Arqueología Amazónica; Instituto Frances de Estudios Andinos: Lima, Peru, 2013; pp. 39–50. [Google Scholar]
- Ríos, G.M.; Becerra, G.C.; Rubio, D.M.; Calvo, C.E.F.; Cavalier, I. Las palmas entre los grupos cazadores-recolectores de la Amazonía colombiana. Caldasia 1998, 20, 57–74. [Google Scholar]
- Van der Hammen, T. Bases para una prehistoria ecológica amazónica y el caso de Chiribiquete. In Pueblos y Paisajes Antiguos de la Selva Amazónica; Morcote, G., Mora, S., Franky, C., Eds.; Universidad Nacional de Colombia, Facultad Ciencias, Taraxacum: Bogotá, Colombia, 2006; pp. 19–28. [Google Scholar]
- Vriesendorp, C.F.; Pitman, N.C.A.; Alvira, D.; Molano, A.S.; García, R.B.; Arciniegas, A.; de Souza, L.S.; del Campo, Á.; Stotz, D.F.; Wachter, T.; et al. (Eds.) Colombia: La Lindosa, Capricho, Cerritos; Field Museum: Chicago, IL, USA, 2018. [Google Scholar]
- Cárdenas, D.; Politis, G. Territorio, Movilidad, Etnobotánica y Manejo del Bosque de los Nukak Orientales: Amazonia Colombiana; Ediciones Uniandes, Departamento de Antropología: Bogotá, Colombia, 2000. [Google Scholar]
- Behling, H.; Hooghiemstra, H. Holocene Amazon rainforest–savanna dynamics and climatic implications: High-resolution pollen record from Laguna Loma Linda in eastern Colombia. J. Quat. Sci. Publ. Quat. Res. Assoc. 2000, 15, 687–695. [Google Scholar] [CrossRef]
- Bush, M.B. The resilience of Amazonian forests. Nature 2017, 541, 167–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [Green Version]
- Correal, G.; Piñeros, F.; Van Der Hammen, T. Guayabero I: Un sitio precerámico de la localidad Angostura II, San José del Guaviare. Caldasia 1990, 16, 245–254. [Google Scholar]
- Gheerbrant, A. La Expedición Orinoco-Amazonas (1948–1950); Banco de la República: Bogotá, Colombia, 1997. [Google Scholar]
- Andrefsky, W. Lithic studies. In Handbook of Archaeological Methods; Rowman Altamira: Walnut Creek, CA, USA, 2005; Volume 1, p. 715. [Google Scholar]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Rowman Altamira: Walnut Creek, CA, USA, 2006. [Google Scholar]
- Ríos, H.G.M.; Cañas, D.A.G.; Raz, L. Catálogo Ilustrado de Fitolitos Contemporáneos con Énfasis Arqueológico y Paleoecológico. I. Gramíneas Amazónicas de COLOMBIA; Universidad Nacional de Colombia, Facultad de Ciencias, Instituto de Ciencias Naturales: Bogotá, Colombia; Reference Publications, Inc: Algonac, MI, USA, 2015. [Google Scholar]
- Pesce, C. Oil Palms and other Oilseeds of the Amazon; Springer: Berlin/Heidelberg, Germany, 1985. [Google Scholar]
- Galeano, G.; Bernal, R. Palmas de Colombia: Guía de Campo; Panamericana Formas e Impreso SA: Bogotá, Colombia, 2010. [Google Scholar]
- Henderson, A.; Galeano, G.; Bernal, R. Field Guide to the Palms of the Americas; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar]
- Politis, G. Nukak: Ethnoarchaeology of an Amazonian People; Left Coast Press: Walnut Creek, CA, USA, 2009. [Google Scholar]
- Kress, W.J.; Betancur, J.; Echeverry, B. Heliconias: Llamaradas de la Selva Colombiana; Cristina Uribe Ediciones: Bogotá, Colombia, 1999. [Google Scholar]
- Galvis, G.; Mojica, J.I.; Duque, S.R.; Castellanos, C.; Sánchez-Duarte, P.; Arce, M.; Gutiérrez-Cortés, A.; Jimenez-Segura, L.F.; Santos-Acevedo, M.; Rivadeneira, S.V.; et al. Peces del Medio Amazonas región de Leticia; Panamericana, formas e impresos SA: Bogotá, Colombia, 2006. [Google Scholar]
- Toledo-Piza, M. Family Cynodontidae. In Checklist of the Freshwater Fishes of South and Central America; Edipucrs: Porto Alegre, Brazil, 2003; pp. 23–237. [Google Scholar]
- Arévalo, H.F.L.; Rodríguez, J.V.D.; Carrillo-Villamizar, J.Z.; Ordóñez, N.C.D. Guía de Mamíferos de San José del Guaviare: Guía de Campo del Instituto de Ciencias Naturales no 25; Universidad Nacional de Colombia: Bogotá, Colombia, 2019. [Google Scholar]
- Medina-Rangel, G.F.; Méndez-Galeano, M.Á.; Calderón-Espinosa, M.L. Herpetofauna of San José del Guaviare, Guaviare, Colombia. Biota Colomb. 2019, 20, 75–90. [Google Scholar] [CrossRef]
- Lasso, C.A.; Morales-Betancourt, M.A.; Vera-Ardila, M.L.; Linares, E.L. Moluscos Dulceacuícolas de Colombia; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2019. [Google Scholar]
- Morcote-Rios, G.; Bernal, R. Remains of palms (Palmae) at archaeological sites in the New World: A review. Bot. Rev. 2001, 67, 309–350. [Google Scholar] [CrossRef]
- Balick, M.J. Systematics and economic botany of the Oenocarpus-Jessenia (Palmae) complex. Adv. in Econ. Bot. 1986, 3, 1–140. [Google Scholar]
- Balick, M.J.; Gershoff, S.N. Nutritional evaluation of the Jessenia bataua palm: Source of high quality protein and oil from tropical America. Econ. Bot. 1981, 35, 261–271. [Google Scholar] [CrossRef]
- Magalhães, M.P. A Phýsis da Origem: O Sentido da História da Amazônia; Museu Paraense Emílio Goeldi: Belém, Brazil, 2005. [Google Scholar]
- LIMA, P. Paleoambiente e Paisagem Durante o Holoceno em Cannãa dos Carajás, Pará, Brasil. Ph.D. Thesis, Universidade Federal Rural do Pernambuco, Recife, Brazil, 2018; 189p. [Google Scholar]
- Prous, A. Fouilles de l’abri du Boquete, Minas Gerais, Brésil. J. Soc. Am. 1991, 77, 77–109. [Google Scholar] [CrossRef]
- Resende, E.T.; Prous, A. Os vestígios vegetais do Grande Abrigo de Santana do Riacho. Arq. Mus. História Nat. Univ. Fed. Minas Gerais 1991, 12, 87–111. [Google Scholar]
- Balslev, H.; Bernal, R.; Fay, M.F. Palms–emblems of tropical forests. Bot. J. Linn. Soc. 2016, 182, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Macía, M.J.; Armesilla, P.J.; Cámara-Leret, R.; Paniagua-Zambrana, N.; Villalba, S.; Balslev, H.; Pardo-de-Santayana, M. Palm uses in northwestern South America: A quantitative review. Bot. Rev. 2011, 77, 462–570. [Google Scholar] [CrossRef]
- Mesa, L.; Galeano, G. Usos de las Palmas en la Amazonia Colombiana: Palms uses in the Colombian Amazon. Caldasia 2013, 35, 351–369. [Google Scholar]
- Zambrana NY, P.; Byg, A.; Svenning, J.C.; Moraes, M.; Grandez, C.; Balslev, H. Diversity of palm uses in the western Amazon. Biodivers. Conserv. 2007, 16, 2771–2787. [Google Scholar] [CrossRef]
- Smith, N. Palms and People in the Amazon; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Henderson, A. The palms of the Amazon; Oxford University Press: New York, NY, USA, 1995. [Google Scholar]
- Wallace, A.R. Palm Trees of the Amazon and Their Uses; J. van Voorst: London, UK, 1853. [Google Scholar]
- Roosevelt, A.C. The Amazon and the Anthropocene: 13,000 years of human influence in a tropical rainforest. Anthropocene 2013, 4, 69–87. [Google Scholar] [CrossRef]
- Behling, H.; Berrio, J.C.; Hooghiemstra, H. Late Quaternary pollen records from the middle Caquetá river basin in central Colombian Amazon. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1999, 145, 193–213. [Google Scholar] [CrossRef]
- Hooghiemstra, H.; van der Hammen, T. Late Quaternary vegetation history and paleoecology of Laguna Pedro Palo (subandean forest belt, Eastern Cordillera, Colombia). Rev. Palaeobot. Palynol. 1993, 77, 235–262. [Google Scholar] [CrossRef]
- Rodríguez, C. Asentamientos de los bosques subandinos durante el Holoceno Medio. Ámbito Ocup. Tempranas Am. Trop. 1995, 115, 123. [Google Scholar]
- López Castaño, C.E. Ocupaciones Tempranas en las Tierras Bajas Tropicales del Valle Medio del río Magdalena Sitio 05-yon-002, Yondo-Antioquia; Fundación de Investigaciones Arqueológicas, Banco de la Republica: Bogotá, Colombia, 1999. [Google Scholar]
- López, C.E. Landscapes variability and the early peopling of the inter-Andean Magdalena Valley, Colombia (South America). Quat. Int. 2021, 578, 139–154. [Google Scholar] [CrossRef]
- Balick, M.J. Ethnobotany of palms in the Neotropics. Adv. Econ. Bot. 1984, 1, 9–23. [Google Scholar]
- Balick, M.J. The use of palms by the Apinayé and Guajajara Indians of Northeastern Brazil. Adv. Econ. Bot. 1988, 6, 65–90. [Google Scholar]
- Clement, C.R.; Denevan, W.M.; Heckenberger, M.J.; Junqueira, A.; Neves, E.G.; Teixeira, W.G.; Woods, W.I. The domestication of Amazonia before European conquest. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150813. [Google Scholar] [CrossRef] [Green Version]
- Levis, C.; Costa, F.R.C.; Bongers, F.; Peña-Claros, M.; Clement, C.R.; Junqueira, A.B.; Neves, E.G.; Tamanaha, E.K.; Figueiredo, F.O.G.; Salomão, R.P.; et al. Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science 2017, 355, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, G.; Dufour, D.; Thomas, E.; Van Zonneveld, M.; Salamanca, A.F.E.; Toro, A.G.; Rivera, A.; Duque, H.S.; Baron, H.S.; Gallego, G.; et al. An integrated hypothesis on the domestication of Bactris gasipaes. PLoS ONE 2015, 10, e0144644. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, M.; Morcote-Rios, G.; Aceituno, F.J.; Roberts, P.; Berrío, J.C.; Iriarte, J. ‘Moving South’: Late Pleistocene Plant Exploitation and the Importance of Palm in the Colombian Amazon. Quaternary 2021, 4, 26. https://doi.org/10.3390/quat4030026
Robinson M, Morcote-Rios G, Aceituno FJ, Roberts P, Berrío JC, Iriarte J. ‘Moving South’: Late Pleistocene Plant Exploitation and the Importance of Palm in the Colombian Amazon. Quaternary. 2021; 4(3):26. https://doi.org/10.3390/quat4030026
Chicago/Turabian StyleRobinson, Mark, Gaspar Morcote-Rios, Francisco Javier Aceituno, Patrick Roberts, Juan Carlos Berrío, and José Iriarte. 2021. "‘Moving South’: Late Pleistocene Plant Exploitation and the Importance of Palm in the Colombian Amazon" Quaternary 4, no. 3: 26. https://doi.org/10.3390/quat4030026
APA StyleRobinson, M., Morcote-Rios, G., Aceituno, F. J., Roberts, P., Berrío, J. C., & Iriarte, J. (2021). ‘Moving South’: Late Pleistocene Plant Exploitation and the Importance of Palm in the Colombian Amazon. Quaternary, 4(3), 26. https://doi.org/10.3390/quat4030026