
Neanderthal Fossils, Mobile Toolkit and a Hyena Den: The Archaeological Assemblage of Lateral Gallery 1 in Cova Del Gegant (NE Iberian Peninsula)

 ,
,  , and
, and
Abstract
:1. Introduction
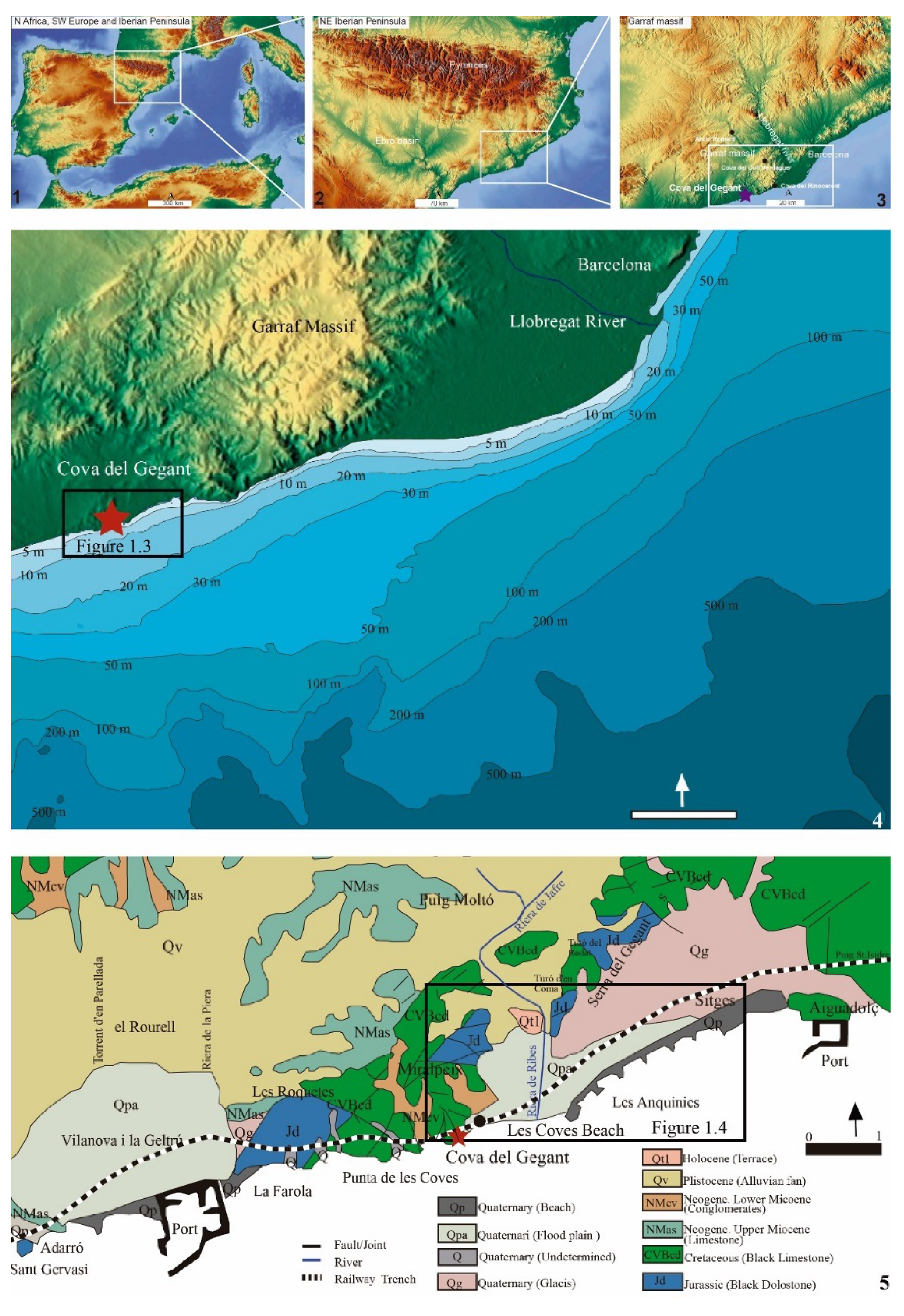
2. Study Area and Site Description
3. Materials and Methods
3.1. Dating Methods
3.2. Faunal Remains
3.3. Lithic Assemblage
3.4. Human Remains
4. Results
4.1. GL1 Stratigraphy and Chronology
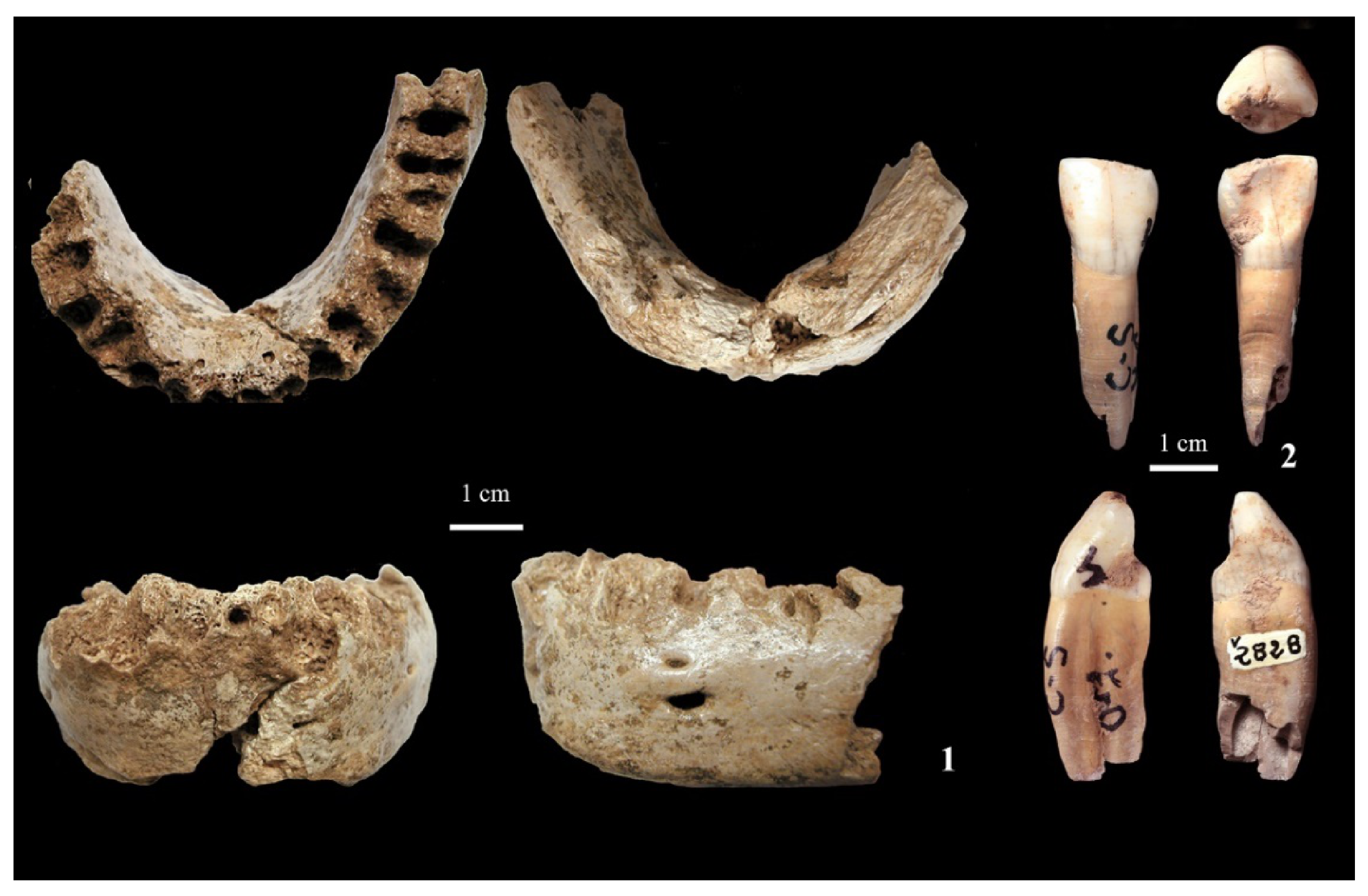
4.2. Neanderthal Remains
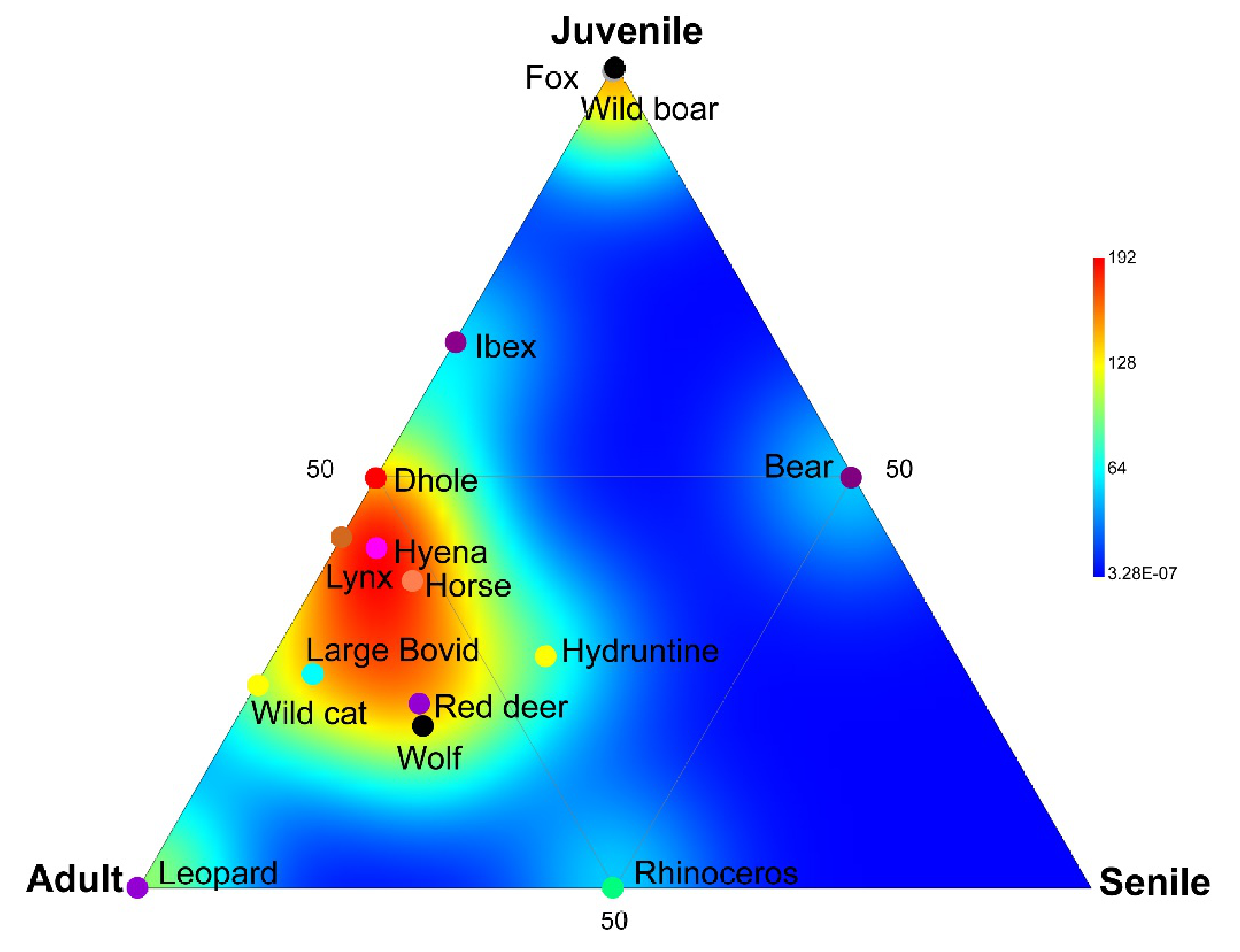
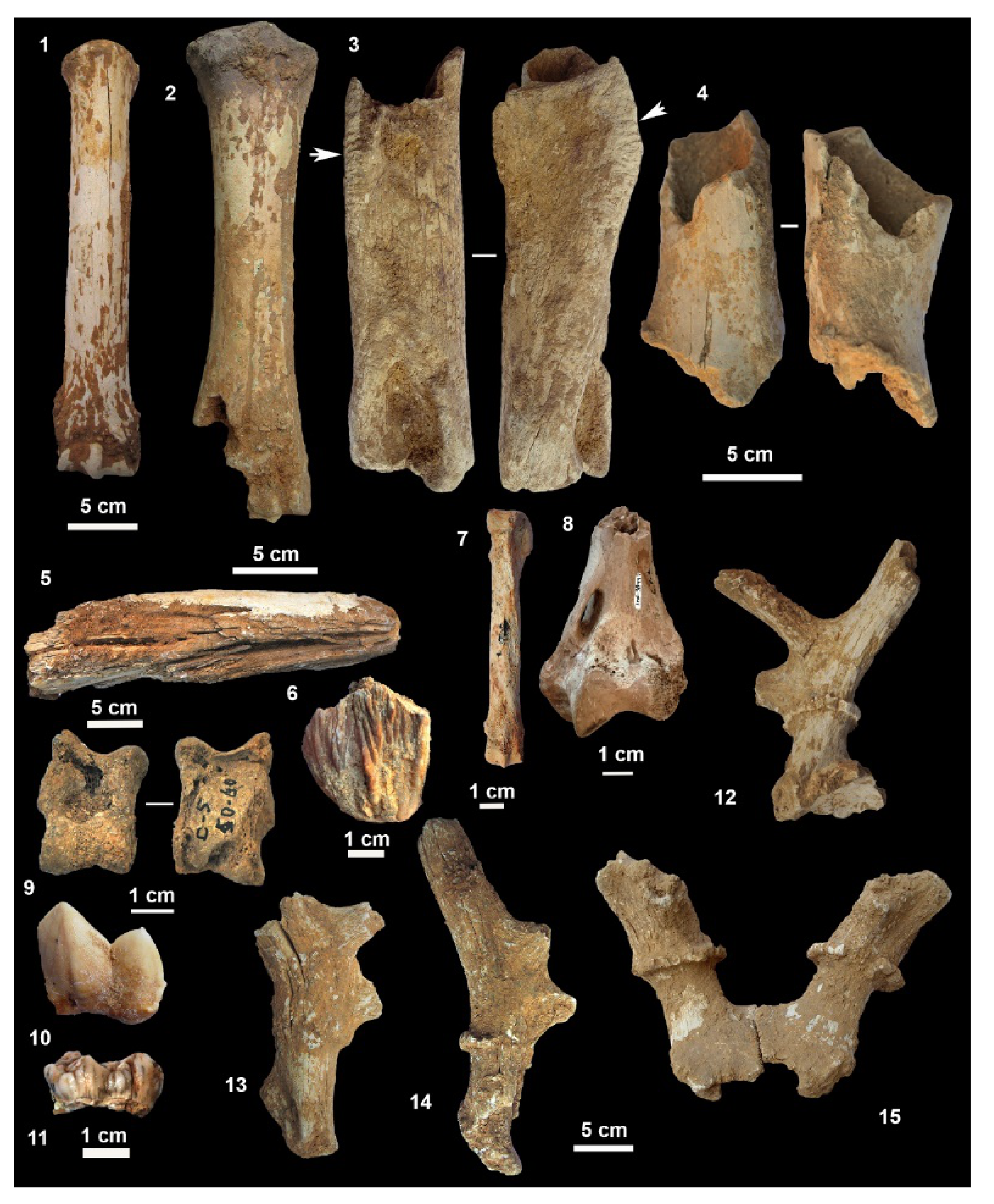
4.3. Large Mammals
4.4. Birds
4.5. Small Vertebrates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
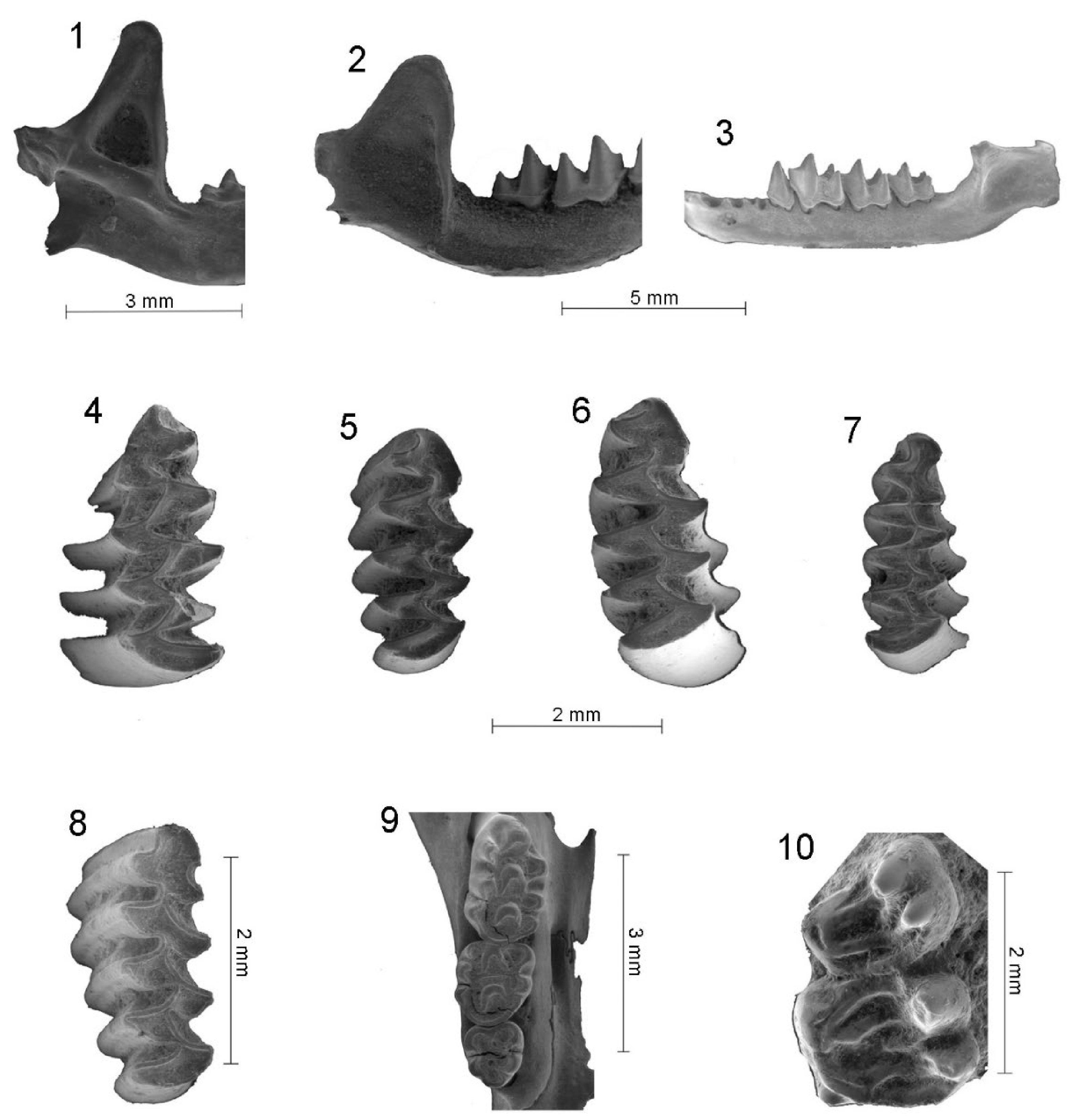{kind=link}
{kind=link}
{kind=link}
| Cova del Gegant | Currently at Garraf Massif |
|---|---|
| Order Eulipotyphla | |
| Erinaceus europaeus | |
| Talpa europaea | |
| Crocidura russula | Crocidura russula |
| Sorex gr. coronatus -araneus | |
| Order Rodentia | |
| Hystrix (A.) cf. brachyura-vinogradovi | |
| Sciurus vulgaris | |
| Arvicola sapidus | |
| Microtus arvalis | |
| Microtus agrestis | |
| Microtus (Iberomys) cabrerae | |
| Microtus (Terricola) duodecimcostatus | Microtus (Terricola) duodecimcostatus |
| Microtus (Terricola) pyrenaicus | |
| Apodemus sylvaticus | Apodemus sylvaticus |
| Rattus rattus | |
| Rattus norvegicus | |
| Mus spretus | |
| Mus musculus | |
| Eliomys quercinus | Eliomys quercinus |
4.6. Lithic Assemblage
5. Discussion
5.1. GL1 History and Correlation
5.2. GL1 Neanderthal Presence and Palaeoenvironmental Record
5.3. Cova Del Gegant in the Regional Context
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bailey, G. Time Perspectives, Palimpsests and the Archaeology of Time. J. Anthropol. Archaeol. 2007, 26, 198–223. [Google Scholar] [CrossRef] [Green Version]
- Bailey, G.; Galanidou, N. Caves, Palimpsests and Dwelling Spaces: Examples from the Upper Palaeolithic of South-East Europe. World Archaeol. 2009, 41, 215–241. [Google Scholar] [CrossRef]
- Stiner, M.C. The Faunas of Hayonim Cave, Israel: A 200,000-Year Record of Paleolithic Diet, Demography, and Society; Harvard University Press: Cambrige, MA, USA, 2005. [Google Scholar]
- Daura, J.; Sanz, M.; Oms, F.X.; Pedro, M.; Martínez, P.; Mendiela, S.; Oliva Poveda, M.; Gibaja, J.F.; Mozota, M.; Alonso-Eguíluz, M.; et al. Deciphering Neolithic Activities from a Cardial Burial Site (Cova Bonica) on the Western Mediterranean Coast. J. Archaeol. Sci. Rep. 2019, 23, 324–347. [Google Scholar] [CrossRef]
- Daura, J.; Sanz, M.; Pike, A.W.G.; Subirà, M.E.; Fornós, J.J.; Fullola, J.M.; Julià, R.; Zilhão, J. Stratigraphic Context and Direct Dating of the Neandertal Mandible from Cova Del Gegant (Sitges, Barcelona). J. Hum. Evol. 2010, 59, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Daura, J.; Sanz, M.; Demuro, M.; Arnold, L.J.; Costa, A.M.; Moreno, J.; Freitas, M.C.; Lopes, V.; Égüez, N.; Hoffman, D.L.; et al. A New Chronological Framework and Site Formation History for Cova Del Gegant (Barcelona): Implications for Neanderthal and Anatomically Modern Human Occupation of NE Iberian Peninsula. Quat. Sci. Rev. 2021, 270, 107–141. [Google Scholar] [CrossRef]
- Daura, J.; Sanz, M.; Subirá, M.E.; Quam, R.; Fullola, J.M.; Arsuaga, J.L. A Neandertal Mandible from the Cova Del Gegant (Sitges, Barcelona, Spain). J. Hum. Evol. 2005, 49, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; García-González, R.; Sanz, M.; Daura, J.; Quam, R.; Fullola, J.M.; Arsuaga, J.L. A Neanderthal Lower Incisor from Cova Del Gegant (Sitges, Barcelona, Spain). Bol. R. Soc. Esp. Hist. Nat. Sec. Geol. 2011, 105, 25–30. [Google Scholar]
- Arsuaga, J.L.; Quam, R.; Daura, J.; Sanz, M.; Subira, M.E.; Dalén, L.; Götherström, A. Neandertal MtDNA from a Late Pleistocene Human Mandible from the Cova Del Gegant (Spain). In Vertebrate Paleobiology and Paleoanthropology; Springer: Dordrecht, The Netherlands, 2011; pp. 213–217. [Google Scholar] [CrossRef]
- Sanz, M. Patrons d’acumulació de Restes de Fauna Del Plistocè Superior Al Nord-Est Peninsular (Àrea Del Massís Del Garraf-Ordal). Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 2013. [Google Scholar]
- Daura, J.; Sanz, M. Procedencia Estratigráfica de Los Restos Humanos Neandertales de La Cova Del Gegant (Sitges, Barcelona). Mainake 2011, 33, 215–232. [Google Scholar]
- Samper, S.C. Who Let the Hyenas Out? Taphonomic Analysis of the Faunal Assemblage from Upper Pleistocene Site of Cova Del Gegant (Sitges, Spain); The University of Edinburg: Edinburg, Scotland, 2011. [Google Scholar]
- Sanz, M.; Daura, J. La Fauna Del Pleistoceno Superior Asociada a Los Restos Humanos Neandertales de La Galería Lateral 1 de La Cova Del Gegant (Sitges, Barcelona). SAGVNTVM 2020, 20, 81–98. [Google Scholar]
- Arsuaga, J.L.; Martinez, I.; Arnold, L.J.; Aranburu, A.; Gracia-Tellez, A.; Sharp, W.D.; Quam, R.M.; Falgueres, C.; Pantoja-Perez, A.; Bischoff, J.; et al. Neandertal Roots: Cranial and Chronological Evidence from Sima de Los Huesos. Science 2014, 344, 1358–1363. [Google Scholar] [CrossRef]
- Meyer, M.; Arsuaga, J.-L.; de Filippo, C.; Nagel, S.; Aximu-Petri, A.; Nickel, B.; Martínez, I.; Gracia, A.; de Castro, J.M.B.; Carbonell, E.; et al. Nuclear DNA Sequences from the Middle Pleistocene Sima de Los Huesos Hominins. Nature 2016, 531, 504–507. [Google Scholar] [CrossRef]
- Zilhão, J. The Late Persistence of the Middle Palaeolithic and Neandertals in Iberia: A Review of the Evidence for and against the “Ebro Frontier” Model. Quat. Sci. Rev. 2021, 270, 107098. [Google Scholar] [CrossRef]
- Zilhão, J.; Angelucci, D.E.; Igreja, M.A.; Arnold, L.J.; Badal, E.; Callapez, P.; Cardoso, J.L.; D’Errico, F.; Daura, J.; Demuro, M.; et al. Last Interglacial Iberian Neandertals as Fisher-Hunter-Gatherers. Science 2020, 367, eaaz7943. [Google Scholar] [CrossRef]
- Quam, R.; Sanz, M.; Daura, J.; Robson Brown, K.; García-González, R.; Rodríguez, L.; Dawson, H.; Rodríguez, R.F.; Gómez, S.; Villaescusa, L.; et al. The Neandertals of Northeastern Iberia: New Remains from the Cova Del Gegant (Sitges, Barcelona). J. Hum. Evol. 2015, 81, 13–28. [Google Scholar] [CrossRef]
- Salas, R. El Malm i El Cretaci Inferior Entre El Massís Del Garraf i La Serra dEspadà. PhD Thesis, Universitat de Barcelona, Barcelona, Spain, 1987. [Google Scholar]
- Daura, J.; Sanz, M.; Fornós, J.; Asensio, A.; Julià, R. Karst Evolution of the Garraf Massif (Barcelona, Spain): Doline Formation, Chronology and Archaeo-Palaeontological Archives. J. Cave Karst Stud. 2014, 76, 69–87. [Google Scholar] [CrossRef]
- Marqués, M.A.; Julià, R. Geomorphological Mapping of Mediterranean Coastal Features, Northeast Spain. J. Coast. Res. 1987, 3, 29–36. [Google Scholar]
- Daura, J.; Sanz, M.; Ramos, J.; Riera, S.; Miras, Y.; Allué, E.; Picornell-Gelabert, L.; López-Reyes, D.; Albert, R.M.; Macià, L.; et al. Palaeoenvironmental Record of the Cal Maurici Wetland Sediment Archive in Barcelona (NE Iberian Peninsula) between c. 6000 and 4000 Cal. Yr BP. Holocene 2016, 26, 1020–1039. [Google Scholar] [CrossRef] [Green Version]
- Virella, A. Vilanova i La Geltrú. Imatges de La Ciutat i de La Comarca; Joan Rius Vila: Vilanova i la Geltrú, Spain, 1949. [Google Scholar]
- Daura, J.; Sanz, M. Nota Necrológica: Santiago Casanova i Giner (Forcall 1922–Barcelona 2011). Cypsela 2017, 20, 7–8. [Google Scholar]
- Viñas, R.; Villalta, J. El Depósito Cuaternario de La “Cova Del Gegant.” In Speleon V Symposium de Espeleología; Centre Excursionista Catalunya: Barcelona, Spain, 1975; pp. 19–33. [Google Scholar]
- Viñas, R. Observaciones Sobre Los Depósitos Cuaternarios de La Cova Del Gegant. Sitges (Barcelona). Speleon 1972, 19, 115–126. [Google Scholar]
- Bellmunt, J. Crónica de La Sección Arqueológica. Boletín Bibl. -Mus. Balaguer 1958, 5, 132–134. [Google Scholar]
- Martínez-Moreno, J.; Miret, J.; Mora, R.; Muro, I. Excavacions a La Cova Del Gegant. Butlletí d’estudis Sitgetans 1985, 32–33, 1. [Google Scholar]
- Martínez-Moreno, J. Informe Técnico de Los Restos Óseos de La Cova Del Gegant (Sitges, Garraf). Servei d’Arqueologia i Paleontologia; Centre d’Informació i Documentació del Patrimoni Cultural de la Generalitat de Catalunya: Barcelona, Spain, 1990. [Google Scholar]
- Martínez-Moreno, J.; Mora, R.; Roca, G.; Parcerisas, J. Memoria d’Excavació a La Cova Del Gegant 1989. Servei d’Arqueologia i Paleontologia; Centre d’Informació i Documentació del Patrimoni Cultural de la Generalitat de Catalunya: Barcelona, Spain, 1990. [Google Scholar]
- Martínez-Moreno, J.; Mora Torcal, R.; Muro Morales, I.; Miret i Mestre, J. Memòria de l’excavació d’urgència de La Cova Del Gegant (Sitges, Garraf) 1985; Centre d’Informació i Documentació del Patrimoni Cultural de la Generalitat de Catalunya: Barcelona, Spain, 1985. [Google Scholar]
- Sanz, M.; Daura, J.; Égüez, N.; Cabanes, D. On the Track of Anthropogenic Activity in Carnivore Dens: Altered Combustion Structures in Cova Del Gegant (NE Iberian Peninsula). Quat. Int. 2017, 437, 102–114. [Google Scholar] [CrossRef]
- Bischoff, J.L.; Julia, R.; Mora, R. Uranium-Series Dating of the Mousterian Occupation at Abric Romani, Spain. Nature 1988, 332, 68–70. [Google Scholar] [CrossRef]
- Rosenbauer, R.J. UDATE1: A Computer Program for the Calculation of Uranium-Series Isotopic Ages. Comput. Geosci. 1991, 17, 45–75. [Google Scholar] [CrossRef]
- Hoffmann, D.L.; Pike, A.W.G.; García-Diez, M.; Pettitt, P.B.; Zilhão, J. Methods for U-Series Dating of CaCO3 Crusts Associated with Palaeolithic Cave Art and Application to Iberian Sites. Quat. Geochronol. 2016, 36, 104–119. [Google Scholar] [CrossRef]
- Hoffmann, D.L. 230Th Isotope Measurements of Femtogram Quantities for U-Series Dating Using Multi Ion Counting (MIC) MC-ICPMS. Int. J. Mass Spectrom. 2008, 275, 75–79. [Google Scholar] [CrossRef]
- Hoffmann, D.L.; Prytulak, J.; Richards, D.A.; Elliott, T.; Coath, C.D.; Smart, P.L.; Scholz, D. Procedures for Accurate U and Th Isotope Measurements by High Precision MC-ICPMS. Int. J. Mass Spectrom. 2007, 264, 97–109. [Google Scholar] [CrossRef]
- Hoffmann, D.L.; Standish, C.D.; García-Diez, M.; Pettitt, P.B.; Milton, J.A.; Zilhão, J.; Alcolea-González, J.J.; Cantalejo-Duarte, P.; Collado, H.; de Balbín, R.; et al. U-Th Dating of Carbonate Crusts Reveals Neandertal Origin of Iberian Cave Art. Science 2018, 359, 912–915. [Google Scholar] [CrossRef] [Green Version]
- Millard, A.R.; Hedges, R.E.M. A Diffusion-Adsorption Model of Uranium Uptake by Archaeological Bone. Geochim. Et Cosmochim. Acta 1996, 60, 2139–2152. [Google Scholar] [CrossRef]
- Pike, A.W.G.; Eggins, S.; Grün, R.; Hedges, R.E.M.; Jacobi, R.M. U-Series Dating of the Late Pleistocene Mammalian Fauna from Wood Quarry (Steetley), Nottinghamshire, UK. J. Quat. Sci. 2005, 20, 59–65. [Google Scholar] [CrossRef]
- Valero-Garcés, B.L.; González-Sampériz, P.; Gil-Romera, G.; Benito, B.M.; Moreno, A.; Oliva-Urcia, B.; Aranbarri, J.; García-Prieto, E.; Frugone, M.; Morellón, M.; et al. A Multi-Dating Approach to Age-Modelling Long Continental Records: The 135 Ka El Cañizar de Villarquemado Sequence (NE Spain). Quat. Geochronol. 2019, 54, 101006. [Google Scholar] [CrossRef]
- Aitken, M.J. Thermoluminescence Dating; Academic Press: London, UK, 1985. [Google Scholar]
- Nambi, K.S.V.; Aitken, M.J. Annual Dose Conversion Factors for TL and ESR Dating. Archaeometry 1986, 28, 202–205. [Google Scholar] [CrossRef]
- Santafé, J.V.; Casanovas, M.L. Dicherorhinus hemitoechus (Falconer, 1868) (Mammalia, Perissodactyla) Del Yacimiento Pleistocénico de La Cueva Del Gegant (Garraf, Barcelona). Empúries Rev. Món Clàssic Antig. Tardana 1989, 48–50, 310–322. [Google Scholar]
- Samper Carro, S.C.; Martínez-Moreno, J. Who Let the Hyenas out? Taphonomic Analysis of the Faunal Assemblage from GL-1 of Cova Del Gegant (Sitges, Spain). Quat. Int. 2014, 330, 19–35. [Google Scholar] [CrossRef]
- Reitz, E.J.; Wing, E.S. Zooarchaeology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Poplin, F. Un Problème d’ostéologie Quantitative: Calcul d’effectif Initial d’après Appariements. Généralisation Aux Autres Types de Remontages et à d’autres Matériels Archéologiques. Rev. D’archéométrie 1981, 5, 159–165. [Google Scholar] [CrossRef]
- Lyman, R.L. Quantitative Paleozoology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar] [CrossRef]
- Brain, C.K. The Hunters or the Hunted?: An Introduction to African Cave Taphonomy; University of Chicago Press: Chicago, CA, USA, 1981. [Google Scholar]
- Lyman, R.L. Vertebrate Taphonomy; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar] [CrossRef]
- Fernández-Jalvo, Y.; Andrews, P. Atlas of Taphonomic Identifications; Vertebrate Paleobiology and Paleoanthropology; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Johnson, E. Current Developments in Bone Technology. In Advances in Archaeological Method and Theory; Elsevier: Amsterdam, The Netherlands, 1985; pp. 157–235. [Google Scholar] [CrossRef]
- Blumenschine, R.J.; Selvaggio, M.M. Percussion Marks on Bone Surfaces as a New Diagnostic of Hominid Behaviour. Nature 1988, 333, 763–765. [Google Scholar] [CrossRef]
- Haynes, G. A Guide for Differentiating Mammalian Carnivore Taxa Responsible for Gnaw Damage to Herbivore Limb Bones. Paleobiology 1983, 9, 164–172. [Google Scholar] [CrossRef]
- Fosse, P.; Avery, G.; Selva, N.; Smietana, W.; Okarma, H.; Wajrak, A.; Fourvel, J.B.; Madelaine, S. Taphonomie Comparée Des Os Longs d’ongulés Dévorés Par Les Grands Prédateurs Modernes d’Europe et d’Afrique (C. Lupus, P. Brunnea). In Prédateurs dans tous leurs états. Evolution, Biodiversité, Interactions, Mythes, Symboles. XXXIe Rencontres Internationales d’Archéologie et d’Histoire d’Antibes; Brugal, J.-P., Gardeisen, A., Zucker, A., Eds.; APDCA: Antibes, France, 2011; pp. 127–156. [Google Scholar]
- Behrensmeyer, A.K. Taphonomic and Ecologic Information from Bone Weathering. Paleobiology 1978, 4, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Blain, H.-A.; Bailon, S. Catalogue of Spanish Plio-Pleistocene Amphibians and Squamate Reptiles from the Museu de Geologia de Barcelona. Treb. Mus. Geol. Barc. 2006, 14, 61–80. [Google Scholar]
- López-García, J.M.; Agustí, J.; Cuenca-Bescós, G. Catalogue of the Late Quaternary Iberian Micromammals (Rodentia, Eulipothypla, Chiroptera) from the Museu de Geologia de Barcelona. Treb. Mus. Geol. Barc. 2007, 15, 5–23. [Google Scholar]
- López-García, J.M.; Blain, H.-A.; Cuenca-Bescós, G.; Arsuaga, J.L. Chronological, Environmental, and Climatic Precisions on the Neanderthal Site of the Cova Del Gegant (Sitges, Barcelona, Spain). J. Hum. Evol. 2008, 55, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Marco, A. Avifaunas Cuaternarias de La Península Ibérica: Sistemática, Paleocologia, Paleozoografia. Ph.D. Thesis, Universidad Autónoma de Madrid, Madrid, Spain, 2005. [Google Scholar]
- Mir, A. La Industria Lítica de La Cova Del Gegant. Sitges (Barcelona). Speleon. Monogr. Symp. Espeleol. 1975, 39–48. [Google Scholar]
- Mora, R. El Paleolítico Medio En Catalunya: Yacimientos En Cueva y Al Aire Libre. Ph.D. Thesis, Unversitat de Barcelona, Barcelona, Spain, 1988. [Google Scholar]
- Laplace, G. La Typologie Analytique et Structurale. Base Rationalle d’étude Des Industries Lithiques et Osseuses. In Branques des Données Archéologiques; Centre National de la Recherche Scientifique: Paris, France, 1974; pp. 91–143. [Google Scholar]
- Gómez-Alba, J. Catálogo Razonado de Los Vertebrados Fósiles de España Del Museo de Geología de Barcelona (1882-1982). Treb. Mus. Geol. Barc. 1997, 6, 1–296. [Google Scholar]
- Daura, J.; Sanz, M. Historiografia Dels Jaciments Plistocens Al Massís Del Garraf i Curs Baix Del Riu Llobregat. Treb. Mus. Geol. Barc. 2009, 16, 5–38. [Google Scholar]
- Masriera, A. Observaciones Sedimentológicas Sobre El Depósito Cuaternario de La Cova Del Gegant (Sitges, Barcelona). Speleon V Symposium de Espeleología; Centre Excursionista de Catalunya: Barcelona, Spain, 1975; pp. 35–38. [Google Scholar]
- Jaffey, A.H.; Flynn, K.F.; Glendenin, L.E.; Bentley, W.C.; Essling, A.M. Precision Measurement of Half-Lives and Specific Activities of 235U and 238U. Phys. Rev. C 1971, 4, 1889. [Google Scholar] [CrossRef]
- Cheng, H.; Edwards, R.L.; Hoff, J.; Gallup, C.D.; Richards, D.A.; Asmerom, Y. The Half-Lives of Uranium-234 and Thorium-230. Chem. Geol. 2000, 169, 17–33. [Google Scholar] [CrossRef]
- Holden, N.E. Total Half-Lives for Selected Nuclides. Pure Appl. Chem. 1990, 62, 941–958. [Google Scholar] [CrossRef] [Green Version]
- Discamps, E. Ungulate Biomass Fluctuations Endured by Middle and Early Upper Paleolithic Societies (SW France, MIS 5-3): The Contributions of Modern Analogs and Cave Hyena Paleodemography. Quat. Int. 2014, 337, 64–79. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Sánchez Marco, A. Avian Zoogeographical Patterns during the Quaternary in the Mediterranean Region and Paleoclimatic Interpretation. Ardeola 2004, 51, 91–132. [Google Scholar]
- Sánchez Marco, A. Avifauna de La Peña de Estebanvela (Segovia). In La Peña de Estebanvela: Grupos Magdalenienses en el Sur del Duero; Cacho, C., Ripoll, S., Muñoz, F.J., Eds.; Arqueología en Castilla y León 17; Arqueología en Castilla y León; Consejería de Cultura y Turismo de la Junta de Castilla y León: Valladolid, Spain, 2007; pp. 141–144. [Google Scholar]
- Tyrberg, T. Pleistocene Birds of the Palearctic: A Catalogue; Mass: Cambridge, MA, USA, 1998; Volume 27. [Google Scholar]
- Díaz, M.; Asensio, B.; Tellería, J.L. Aves Ibéricas: I. No Paseriformes.; Reyero, J.M., Ed.; Reyero, J.M.: Madrid, Spain, 1996. [Google Scholar]
- Tellería, J.L.; Asensio, B.; Díaz, M. Aves Ibéricas: II. Paseriformes; Reyero, J.M., Ed.; Reyero, J.M.: Madrid, Spain, 1999. [Google Scholar]
- Blanco, G.; Sánchez-Marco, A.; Negro, J.J. Night Capture of Roosting Cave Birds by Neanderthals: An Actualistic Approach. Front. Ecol. Evol. 2021, 9, 9. [Google Scholar] [CrossRef]
- Blain, H.-A.; Bailon, S.; Cuenca-Bescós, G.; Arsuaga, J.L.; Bermúdez de Castro, J.M.; Carbonell, E. Long-Term Climate Record Inferred from Early-Middle Pleistocene Amphibian and Squamate Reptile Assemblages at the Gran Dolina Cave, Atapuerca, Spain. J. Hum. Evol. 2009, 56, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blain, H.-A.; Lozano-Fernández, I.; Agustí, J.; Bailon, S.; Menéndez Granda, L.; Espígares Ortiz, M.P.; Ros-Montoya, S.; Jiménez Arenas, J.M.; Toro-Moyano, I.; Martínez-Navarro, B.; et al. Refining upon the Climatic Background of the Early Pleistocene Hominid Settlement in Western Europe: Barranco León and Fuente Nueva-3 (Guadix-Baza Basin, SE Spain). Quat. Sci. Rev. 2016, 144, 132–144. [Google Scholar] [CrossRef]
- Font-Tullot, I. Climatología de España y Portugal; Ediciones Universidad de Salamanca: Salamanca, Spain, 2000. [Google Scholar]
- Andrews, P. Owls, Caves and Fossils. Predation, Preservation and Accumulation of Small Mammañ Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip, Somerset, United Kingdom; Oxford University: Oxford, UK, 1990. [Google Scholar]
- Pokines, J.T.; Kerbis Peterhans, J.C. Barn Owl (Tyto Alba) Taphonomy in the Negev Desert, Israel. Isr. J. Ecol. Evol. 1998, 44, 19–27. [Google Scholar] [CrossRef]
- Palomo, L.J.; Gisbert, J.; Blanco, J.C. Atlas y Libro Rojo de Los Mamíferos Terrestres de España; Dirección General para la Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007. [Google Scholar]
- Cruz-Uribe, K. Distinguishing Hyena from Hominid Bone Accumulations. J. Field Archaeol. 1991, 18, 467–486. [Google Scholar]
- Fosse, P. Variabilité Des Assemblages Osseux Créés Par l’hyène Des Cavernes. Paléo 1997, 9, 15–54. [Google Scholar] [CrossRef]
- Alcaraz-Castaño, M.; Alcolea-González, J.; Kehl, M.; Albert, R.-M.; Baena-Preysler, J.; de Balbín-Behrmann, R.; Cuartero, F.; Cuenca-Bescós, G.; Jiménez-Barredo, F.; López-Sáez, J.-A.; et al. A Context for the Last Neandertals of Interior Iberia: Los Casares Cave Revisited. PLoS ONE 2017, 12, e0180823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsuaga, J.L.; Baquedano, E.; Pérez-González, A.; Sala, N.; Quam, R.M.; Rodríguez, L.; García, R.; García, N.; Álvarez-Lao, D.J.; Laplana, C.; et al. Understanding the Ancient Habitats of the Last-Interglacial (Late MIS 5) Neanderthals of Central Iberia: Paleoenvironmental and Taphonomic Evidence from the Cueva Del Camino (Spain) Site. Quat. Int. 2012, 275, 55–75. [Google Scholar] [CrossRef] [Green Version]
- Daura, J.; Sanz, M.; Julià, R.; García-Fernández, D.; Fornós, J.J.; Vaquero, M.; Allué, E.; López-García, J.M.; Blain, H.A.; Ortiz, J.E.; et al. Cova Del Rinoceront (Castelldefels, Barcelona): A Terrestrial Record for the Last Interglacial Period (MIS 5) in the Mediterranean Coast of the Iberian Peninsula. Quat. Sci. Rev. 2015, 114, 203–227. [Google Scholar] [CrossRef] [Green Version]
- Daura, J.; Sanz, M.; Allué, E.; Vaquero, M.; López-García, J.M.; Sánchez-Marco, A.; Domènech, R.; Martinell, J.; Carrión, J.S.; Ortiz, J.E.; et al. Palaeoenvironments of the Last Neanderthals in SW Europe (MIS 3): Cova Del Coll Verdaguer (Barcelona, NE of Iberian Peninsula). Quat. Sci. Rev. 2017, 177, 34–56. [Google Scholar] [CrossRef]
- Vaquero, M.; van der Made, J.; Blain, H.-A.; Ibáñez, N.; López-García, J.M.; Rivals, F.; Alonso, S.; Ameijenda, A.; Bennàsar, M.; Fernández-García, M.; et al. Fauna, Environment and Human Presence during MIS5 in the North of Spain: The New Site of Valdavara 3. Comptes Rendus Palevol 2018, 17, 557–593. [Google Scholar] [CrossRef]
- Villa, P.; Soressi, M. Stone Tools in Carnivore Sites: The Case of Bois Roche. J. Anthropol. Res. 2000, 56, 187–215. [Google Scholar] [CrossRef]
- Trinkaus, E.; Maki, J.; Zilhão, J. Middle Paleolithic Human Remains from the Gruta Da Oliveira (Torres Novas), Portugal. Am. J. Phys. Anthropol. 2007, 134, 263–273. [Google Scholar] [CrossRef]
- Camarós, E.; Cueto, M.; Lorenzo, C.; Villaverde, V.; Rivals, F. Large Carnivore Attacks on Hominins during the Pleistocene: A Forensic Approach with a Neanderthal Example. Archaeol. Anthropol. Sci. 2016, 8, 635–646. [Google Scholar] [CrossRef]
- Camarós, E.; Cueto, M.; Rosell, J.; Díez, J.C.; Blasco, R.; Duhig, C.; Darlas, A.; Harvati, K.; Jordá, J.; Montes, L.; et al. Hunted or Scavenged Neanderthals? Taphonomic Approach to Hominin Fossils with Carnivore Damage. Int. J. Osteoarchaeol. 2017, 27, 606–620. [Google Scholar] [CrossRef]
- Garralda, M.D.; Giacobini, G.; Vandermeersch, B. Neanderthal Cutmarks: Combe-Grenal and Marillac (France). A SEM Analysis. Anthropologie 2005, 43, 189–198. [Google Scholar] [CrossRef]
- Orschiedt, J. Der Fall Krapina—Neue Ergebnisse Zur Frage von Kannibalismus Beim Neandertaler. Quartär 2008, 55, 63–81. [Google Scholar]
- Pettitt, P. The Neanderthal Dead. Before Farming 2002, 2002, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Arsuaga, J.L.; Villaverde, V.; Quam, R.; Martínez, I.; Carretero, J.M.; Lorenzo, C.; Gracia, A. New Neandertal Remains from Cova Negra (Valencia, Spain). J. Hum. Evol. 2007, 52, 31–58. [Google Scholar] [CrossRef] [Green Version]
- Zilio, L.; Hammond, H.; Karampaglidis, T.; Sánchez-Romero, L.; Blasco, R.; Rivals, F.; Rufà, A.; Picin, A.; Chacón, M.G.; Demuro, M.; et al. Examining Neanderthal and Carnivore Occupations of Teixoneres Cave (Moià, Barcelona, Spain) Using Archaeostratigraphic and Intra-Site Spatial Analysis. Sci. Rep. 2021, 11, 4339. [Google Scholar] [CrossRef]
- Neruda, P.; Kaminská, L. Neanderthals at the Open-Air Site of Bojnice III: The Issue of “Missing” Artifacts. In Settlement Dynamics of the Middle Paleolithic and Middle Stone Age IV.; Conard, N.J., Delagnes, A., Eds.; Diane Marie Kerns Kerns Verlag: Tübingen, Germany, 2015; pp. 205–226. [Google Scholar]
- Picin, A.; Chacón, M.G.; Gómez de Soler, B.; Blasco, R.; Rivals, F.; Rosell, J. Neanderthal Mobile Toolkit in Short-Term Occupations at Teixoneres Cave (Moia, Spain). J. Archaeol. Sci. Rep. 2020, 29, 102165. [Google Scholar] [CrossRef]
- Vaquero, M.; Chacon, M.G.; Garcia-Anton, M.D.; de Soler, B.G.; Martinez, K.; Cuartero, F. Time and Space in the Formation of Lithic Assemblages: The Example of Abric Romani Level. J. Quat. Int. 2012, 247, 162–181. [Google Scholar] [CrossRef]
- Sanz-Royo, A.; Sanz, M.; Daura, J. Upper Pleistocene Equids from Terrasses de La Riera Dels Canyars (NE Iberian Peninsula): The Presence of Equus Ferus and Equus Hydruntinus Based on Dental Criteria and Their Implications for Palaeontological Identification and Palaeoenvironmental Reconstr. Quat. Int. 2020, 566–567, 78–90. [Google Scholar] [CrossRef]
- Burke, A.; Eisenmann, V.; Ambler, G.K. The Systematic Position of Equus Hydruntinus, an Extinct Species of Pleistocene Equid. Quat. Res. 2003, 59, 459–469. [Google Scholar] [CrossRef]
- Daura, J.; Sanz, M.; García, N.; Allué, E.; Vaquero, M.; Fierro, E.; Carrión, J.S.; López-García, J.M.; Blain, H.A.; Sánchez-Marco, A.; et al. Terrasses de La Riera Dels Canyars (Gavà, Barcelona): The Landscape of Heinrich Stadial 4 North of the “Ebro Frontier” and Implications for Modern Human Dispersal into Iberia. Quat. Sci. Rev. 2013, 60, 26–48. [Google Scholar] [CrossRef] [Green Version]
- López-García, J.M.; Blain, H.-A.; Sanz, M.; Daura, J. A Coastal Reservoir of Terrestrial Resources for Neanderthal Populations in North-Eastern Iberia: Palaeoenvironmental Data Inferred from the Small-Vertebrate Assemblage of Cova Del Gegant, Sitges, Barcelona. J. Quat. Sci. 2012, 27, 105–113. [Google Scholar] [CrossRef]
- Brugal, J.-P.; Jaubert, J. Les Gisements Paléontologiques Pléistocènes à Indices de Fréquentation Humaine: Un Nouveau Type de Comportement de Prédation? Paléo 1991, 3, 15–41. [Google Scholar] [CrossRef]
- Sanz, M.; Daura, J. Taphonomic Analysis of an Ungulate-Dominated Accumulation at the Pleistocene Cova Del Rinoceront Site near Barcelona, Spain (Northeastern Iberian Peninsula). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 498, 24–38. [Google Scholar] [CrossRef]
- Sanz, M.; Daura, J. Carnivore Involvement in Bone Assemblages Based on Taphonomic and Zooarchaeological Analyses of Cova Del Coll Verdaguer Site (Barcelona, Iberian Peninsula). Hist. Biol. 2018, 30, 807–820. [Google Scholar] [CrossRef]
- Beyries, S.; Boëda, E. Etude Technologique et Traces d’utilisation Des “Éclats Débordants” de Corbehem (Pas-de-Calais). Bull. Société Préhistorique Français 1983, 80, 275–279. [Google Scholar] [CrossRef]









| Year | Direction | Institution | Materials Stored | Acronyms |
|---|---|---|---|---|
| 1954 | S. Casanova | AMUNT | Arxiu Històric Municipal de Sites, Museu Mar i Cel, Museu Arqueològic de Catalunya | AHSI MMC MAC |
| 1972, 1974–75 | R. Viñas-J. Villalta | Secció Ecologia Quaternari- Institut Jaume Almera | Museu Geològic de Barcelona Museu Arqueològic de Catalunya | MGB MAC |
| ‘70 | Collected by speleologists (A. Asensio) | Centre Espeleològic Cervelló | Universitat de Barcelona | UB-1 |
| 1985 | J. Martínez- R. Mora- I. Muro- J. Miret | Centre de Recerques Palaeoecosocials de Girona i de la Universitat Autònoma de Barcelona | Universitat Autònoma de Barcelona | UAB |
| 1989 | J. Martínez- R. Mora- J. Parcerisas- G. Roca | Centre de Recerques Palaeoecosocials de Girona i de la Universitat Autònoma de Barcelona | Universitat Autònoma de Barcelona | UAB |
| ‘90 | Collected by speleologists (M. Nebot) | Grup Espeleològic Pedraforca | Institut Català de Paleontologia | ICP |
| 2007–2022 | J. Daura-M. Sanz | Grup Recerca del Quaternari-SERP (Universitat Barcelona) | Universitat de Barcelona | UB-2 |
| Luminescence (IRSL) | ||||||||
|---|---|---|---|---|---|---|---|---|
| # | Sample Name | Layer | Mineral | Grain Size (μm) | Total Dose Rate (Gy/ka) | De (Gy) | Age ka (1σ) | |
| 1 | MAD-5642 | XVb | Polymineral | 2–10 | 1.59 ± 0.10 | 95.5 ± 2.2 | 60.1 ± 3.9 | |
| U-series | ||||||||
| Lab # | Layer | 238U | 232Th | 234U/238U | 230Th/238U | 230Th/232Th | Age ka (2 σ) | |
| 2 | 6006 | XVa | 0.22 | 0.03 | 1.16 ± 0.03 | - | 10.48 | 49.4 ± 1.8 |
| 3 | JD52a | XVa/b | n/a | n/a | 1.262 ± 0.017 | 0.495 ± 0.023 | >100 | 53.2 ± 3.3 |
| 4 | JD52b | XVa/b | n/a | n/a | 1.262 ± 0.017 | 0.495 ± 0.023 | >100 | 53.2 ± 3.3 |
| 5 | UEVA 1028 | XVa | 367.64 ± 3.58 | 37.63 ± 0.39 | 1.1615 ± 0.0024 | 0.4281 ± 0.0023 | 12.78 ± 0.06 | 47.11 ± 1.21 |
| 6 | UEVA 1029 | XVa | 37.63 ± 0.70 | 0.191 ± 0.004 | 1.1621 ± 0.0030 | 0.3933 ± 0.0028 | 237.01 ± 2.40 | 44.51 ± 0.42 |
| NISP | MNI | |
|---|---|---|
| Carnivora | 329 | 55 |
| Canis lupus | 28 | 5 |
| Vulpes vulpes | 11 | 1 |
| Ursus sp./U. arctos | 19 | 2 |
| Meles meles | 3 | 1 |
| Crocuta crocuta | 179 | 24 |
| Felis silvestris | 40 | 8 |
| Lynx pardinus | 30 | 7 |
| Panthera pardus | 13 | 5 |
| Cuon alpinus | 6 | 2 |
| Artiodactyla | 662 | 47 |
| Sus scrofa | 10 | 3 |
| Cervus elaphus | 448 | 22 |
| Bos/Bison | 103 | 19 |
| Bison priscus | 18 | |
| Bos primigenius | 65 | |
| Capra pyrenaica | 18 | 3 |
| Perissodactyla | 930 | 49 |
| Equus ferus | 823 | 40 |
| Equus hydruntinus | 71 | 7 |
| Stephanorhinus hemitoechus | 36 | 2 |
| Proboscidea | 7 | 1 |
| Elephas sp. | 3 | |
| Proboscidea indet. | 4 | 1 |
| Undetermined | ||
| Macrofauna indet. | 674 | |
| Total | 2602 | 152 |
| Layers | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | I–II | II | III | IV | Unknown | Total NISP | Total MNI | |||||||||
| Taxa | NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | NISP | MNI | NISP | %NISP | MNI | %MNI |
| Puffinus puffinus | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Falco tinnunculus | 2 | 2 | 3 | 1 | 5 | 1.4 | 3 | 2.1 | ||||||||
| Accipiter nisus | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Accipitriformes indet. | 1 | 1 | 1 | 1 | 2 | 0.5 | 2 | 1.4 | ||||||||
| Alectoris rufa | 5 | 2 | 6 | 4 | 1 | 1 | 1 | 1 | 2 | 1 | 15 | 4.1 | 9 | 6.2 | ||
| Coturnix coturnix | 5 | 2 | 1 | 1 | 1 | 1 | 4 | 2 | 11 | 3.0 | 6 | 4.1 | ||||
| Columba livia s. oenas | 9 | 3 | 5 | 3 | 2 | 2 | 5 | 2 | 21 | 5.7 | 10 | 6.8 | ||||
| Tyto alba | 1 | 1 | 1 | 1 | 2 | 0.5 | 2 | 1.4 | ||||||||
| Bubo bubo | 2 | 1 | 2 | 0.5 | 1 | 0.7 | ||||||||||
| Athene noctua | 2 | 1 | 3 | 2 | 1 | 1 | 6 | 1.6 | 4 | 2.7 | ||||||
| Strix aluco | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Apus apus | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Delichon urbica | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Anthus spinoletta | 1 | 1 | 2 | 1 | 3 | 0.8 | 2 | 1.4 | ||||||||
| Turdus viscivorus | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Emberiza calandra | 1 | 1 | 2 | 2 | 3 | 0.8 | 3 | 2.1 | ||||||||
| Fringilla coelebs | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Carduelis chloris | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 0.8 | 3 | 2.1 | ||||||
| Loxia pytyopsittacus | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Pinicola enucleator | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Pyrrhula pyrrhula | 1 | 1 | 1 | 0.3 | 1 | 0.7 | ||||||||||
| Coccothraustes coccothraustes | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 0.8 | 3 | 2.1 | ||||||
| Sturnus sp. | 2 | 1 | 2 | 0.5 | 1 | 0.7 | ||||||||||
| Pica pica | 1 | 1 | 1 | 1 | 3 | 1 | 5 | 1.4 | 3 | 2.1 | ||||||
| Pyrrhocorax pyrrhocorax | 76 | 16 | 7 | 3 | 48 | 15 | 13 | 5 | 62 | 9 | 206 | 55.8 | 48 | 32.9 | ||
| Pyrrhocorax graculus | 10 | 6 | 14 | 6 | 10 | 8 | 14 | 2 | 48 | 13.0 | 22 | 15.1 | ||||
| Corvus monedula | 8 | 4 | 2 | 2 | 2 | 2 | 8 | 4 | 20 | 5.4 | 12 | 8.2 | ||||
| Corvus corone | 1 | 1 | 1 | 1 | 2 | 0.5 | 2 | 1.4 | ||||||||
| Total | 369 | 100 | 146 | 100 | ||||||||||||
| Taxon | NISP | MNI | |
|---|---|---|---|
| Amphibians | Pelobates cultripes | 4 | 1 |
| Bufo gr. B. bufo | 4 | 2 | |
| Epidalea calamita | 11 | 3 | |
| Bufonidae indet. | 15 | 0 | |
| Ranidae indet. | 2 | 1 | |
| Anura indet. | 1 | 0 | |
| Testudines | Testudo hermanni | 20 | 2 |
| Squamates | Timon cf. lepidus | 2 | 1 |
| Malpolon monspessulanus | 10 | 1 | |
| Zamenis scalaris | 14 | 1 | |
| Colubrinae indet. | 2 | 0 | |
| Vipera sp. | 2 | 1 | |
| Ophidia indet. | 7 | 0 | |
| Insectivores | Erinaceus europaeus | 1 | 1 |
| Sorex gr. coronatus-araneus | 6 | 4 | |
| Crocidura russula | 10 | 9 | |
| Talpa europaea | 2 | 1 | |
| Bats | Rhinolophus ferrumequinum | 2 | 1 |
| Myotis myotis | 3 | 3 | |
| Myotis sp. | 1 | 1 | |
| Miniopterus schreibersii | 3 | 2 | |
| Rodents | Microtus arvalis | 5 | 4 |
| Microtus agrestis | 5 | 4 | |
| Microtus (Iberomys) cabrerae | 11 | 7 | |
| Microtus (Terricola) duodecimcostatus | 12 | 9 | |
| Microtus (Terricola) pyrenaicus | 2 | 2 | |
| Apodemus sylvaticus | 69 | 29 | |
| Eliomys quercinus | 15 | 6 | |
| Hystrix (Acanthion) cf. brachyura-vinogradovi | 4 | 2 | |
| Total | 245 | 98 |
| Very Small | Small | Medium | Large | Very Large | Total | |
|---|---|---|---|---|---|---|
| Cores | 2 | 2 | ||||
| Flakes | 6 | 8 | 3 | 1 | 18 | |
| Flake fragments | 2 | 1 | 3 | |||
| Retouched artifacts | 3 | 1 | 2 | 6 | ||
| Total | 8 | 9 | 3 | 4 | 5 | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daura, J.; Sanz, M.; Vaquero, M.; López-García, J.M.; Blain, H.-A.; Marco, A.S. Neanderthal Fossils, Mobile Toolkit and a Hyena Den: The Archaeological Assemblage of Lateral Gallery 1 in Cova Del Gegant (NE Iberian Peninsula). Quaternary 2022, 5, 12. https://doi.org/10.3390/quat5010012
Daura J, Sanz M, Vaquero M, López-García JM, Blain H-A, Marco AS. Neanderthal Fossils, Mobile Toolkit and a Hyena Den: The Archaeological Assemblage of Lateral Gallery 1 in Cova Del Gegant (NE Iberian Peninsula). Quaternary. 2022; 5(1):12. https://doi.org/10.3390/quat5010012
Chicago/Turabian StyleDaura, Joan, Montserrat Sanz, Manuel Vaquero, Juan Manuel López-García, Hugues-Alexandre Blain, and Antonio Sánchez Marco. 2022. "Neanderthal Fossils, Mobile Toolkit and a Hyena Den: The Archaeological Assemblage of Lateral Gallery 1 in Cova Del Gegant (NE Iberian Peninsula)" Quaternary 5, no. 1: 12. https://doi.org/10.3390/quat5010012
APA StyleDaura, J., Sanz, M., Vaquero, M., López-García, J. M., Blain, H. -A., & Marco, A. S. (2022). Neanderthal Fossils, Mobile Toolkit and a Hyena Den: The Archaeological Assemblage of Lateral Gallery 1 in Cova Del Gegant (NE Iberian Peninsula). Quaternary, 5(1), 12. https://doi.org/10.3390/quat5010012