6.2. Palynology
Laboratory techniques allow for the achievement of pollen counts ranging from 150 to 600 grains per sample in large samples. It is noted that a maximal differentiation of pollen taxa was achieved in conjunction with University of Texas at Austin Herbarium (Main Building) materials. A differentiation of
Carya was not absolutely achieved, but a tendency of the examined pollen grains from BK and NB to exhibit relatively heavy exine thickening around pores was noted which corresponded to
Carya illinoinensis (this was less evident in hickory species of
Carya). An absolute separation of all individual specimens was not possible. It was further noted that the
Betula pollen grains encountered exhibited poor vestibulum development (thin endexines) and very likely belonged to the species
B. nigra or river birch, according to comparative morphology [
9]. Preservation was pristine for the examined samples. All dating estimates below are presented as calibrated ages in years BP.
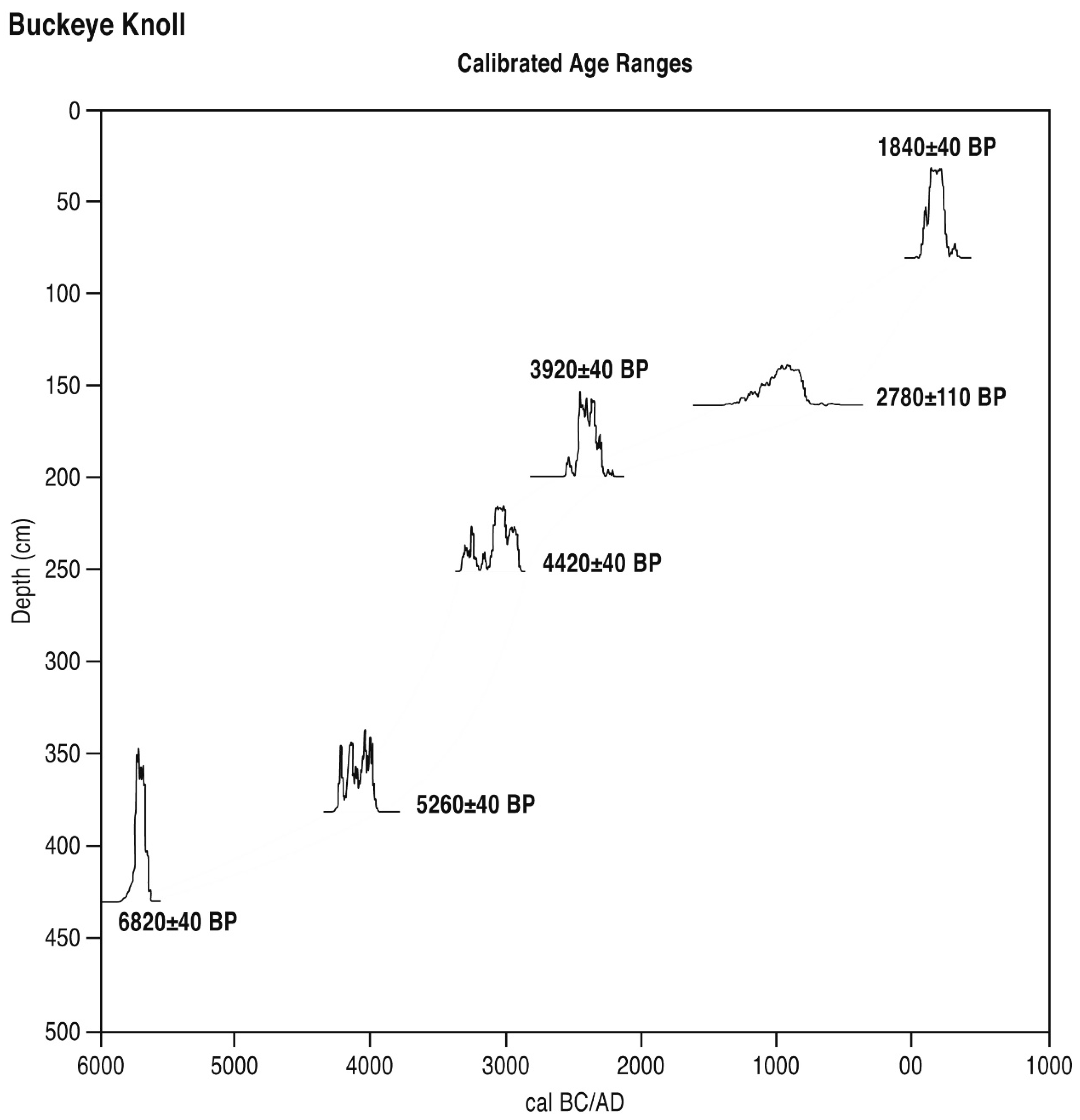
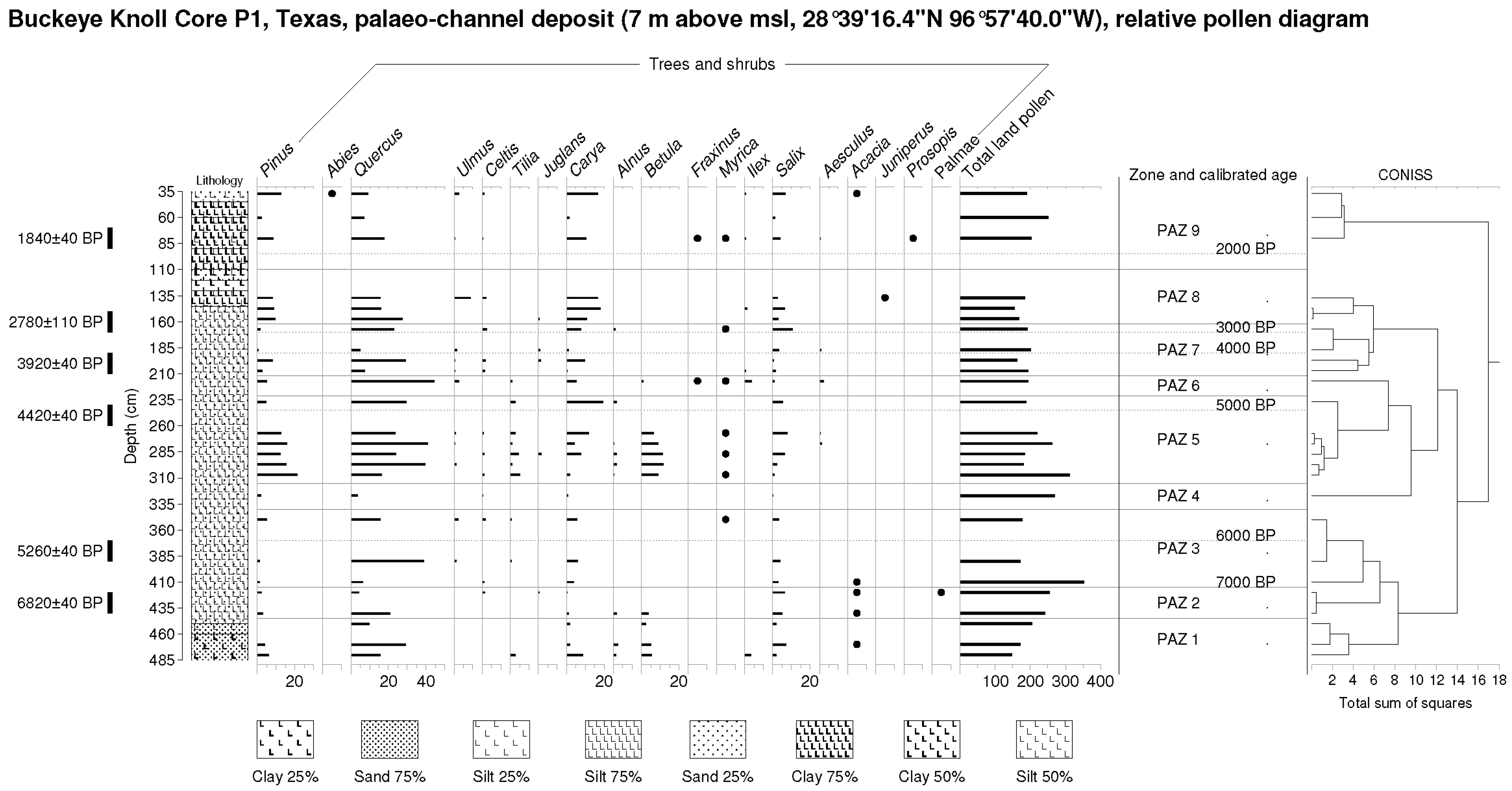
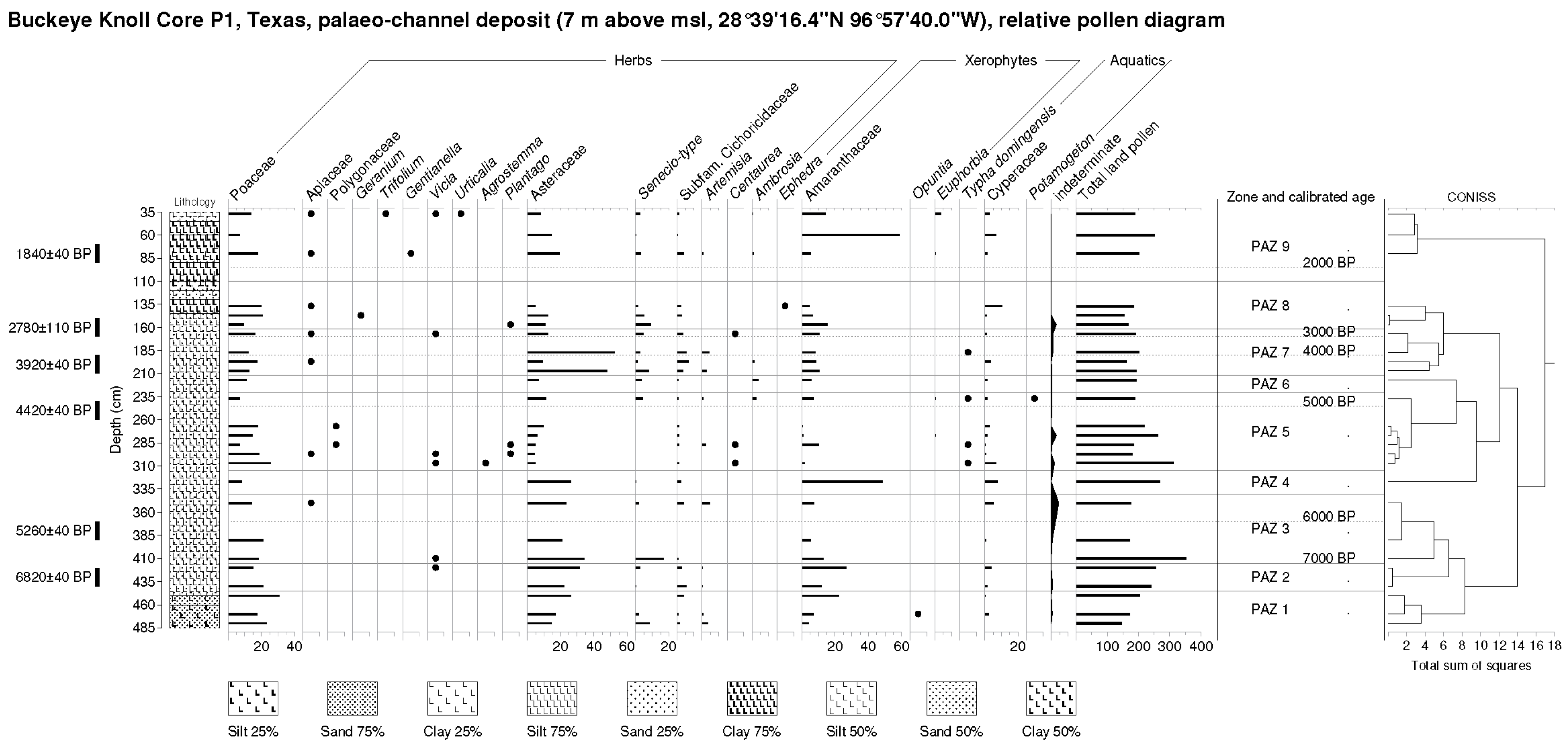
BK Zone 1 (450–480 cm, est. 9300–8300 BP) Quercus (29.2% max.), Carya (8.8% max.), Betula (5.4% max.) and riparian Salix are important trees and shrubs in this pollen zone. Isolates of Alnus and Acacia are noted here. Poaceae (30.7% max.), Asteraceae and other composites are important herbs. Artemisia is present, while values of the xerophyte Amaranthaceae rise (4.2–22.4%). An Opuntia isolate is noted. Freshwater isolates include Cyperaceae.
BK Zone 2 (420–440 cm, 8000–7300 BP). Amaranthaceae values (26.7% max.) remain elevated in this zone, while values of tree pollen (esp. Carya and Betula) decline; however, those of shrubs of the riparian verge as represented by Salix remain unchanged (ca. 5–6%). Isolates of Acacia and Palmae are noted. Herbs include most importantly Poaceae and Asteraceae. Isolates of Cyperaceae are noted.
BK Zone 3 (350–410 cm, 7000–5750 BP). Amaranthaceae pollen levels decline (20.9% min.) in this zone, while tree pollen, particularly Quercus (39.0% max.), rises. Carya establishes a significant presence (ca. 4–6%). Herbs include most importantly Poaceae and Asteraceae. Isolates of Cyperaceae are also noted.
BK Zone 4 (327 cm, 5600 BP). A secondary maximum of Amaranthaceae pollen (48.7%) is noted here as Quercus pollen declines to an absolute minimum (3.7%) as other trees decline. Forbs, as represented by general Asteraceae, exceed grasses, as represented by Poaceae pollen for the first time. Cyperaceae rises here.
BK Zone 5 (237–307 cm, 5400–4800 BP). Tree and shrub pollen led by Pinus (15.6% max.), Quercus (40.8% max.), Carya (19.6% max.), Betula (11.7% max.), and Salix (7.7% max.) increases very significantly in this zone as Amaranthaceae pollen declines markedly (to 0.5% min.). Isolates of Juglans, Alnus, and Myrica are noted. Poaceae rises (to 18.9% max.), also, as Asteraceae declines (to 4.4% min.). Typha grains are further noted.
BK Zone 6 (217 cm, 4600 BP). Quercus (44.3%) dominates tree pollen here, while lower levels of Carya, an absence of Betula, and isolates of Fraxinus and Myrica are noted. Herbaceous pollen is represented by lower values of Poaceae (10.8%) and Asteraceae (6.7%).
BK Zone 7 (167–207 cm, 4400–3200 BP). Lower Quercus, oscillating values of Carya, and rising Salix characterize the tree and shrub pollen of this zone. Herbaceous pollen, particularly Asteraceae (absolute max. 52.5%), rises as Amaranthaceae also increases somewhat (10.5% max.). An isolate of Typha is noted.
BK Zone 8 (137–157 cm, 2900–2600 BP). A further rise of Carya (18.1% max.) is noted along with Quercus pollen as Salix declines in this zone. Herbaceous pollen is constituted principally of Poaceae as Asteraceae declines (to 4.9% min). A maximum of Cyperaceae (10.3%) is noted.
BK Zone 9 (37–80 cm, 1800–400 BP). Tree and shrub pollen consists largely of (declining) Quercus, (oscillating) Carya and Salix in this zone. Isolates of Fraxinus, Myrica and Acacia are noted. Herbaceous pollen consists largely of Poaceae and Asteraceae, while an absolute maximum of Amaranthaceae pollen (58.6%) is noted.
Figure 4.
Buckeye Knoll (BK) relative pollen diagram (arboreal pollen as percentage of TLP) and Troels-Smith lithology. Note redox (red coloration) features.
Figure 4.
Buckeye Knoll (BK) relative pollen diagram (arboreal pollen as percentage of TLP) and Troels-Smith lithology. Note redox (red coloration) features.
Figure 5.
Buckeye Knoll (BK) relative pollen diagram (non-arboreal pollen as percentage of TLP) and Troels-Smith lithology. Note redox (red coloration) features.
Figure 5.
Buckeye Knoll (BK) relative pollen diagram (non-arboreal pollen as percentage of TLP) and Troels-Smith lithology. Note redox (red coloration) features.
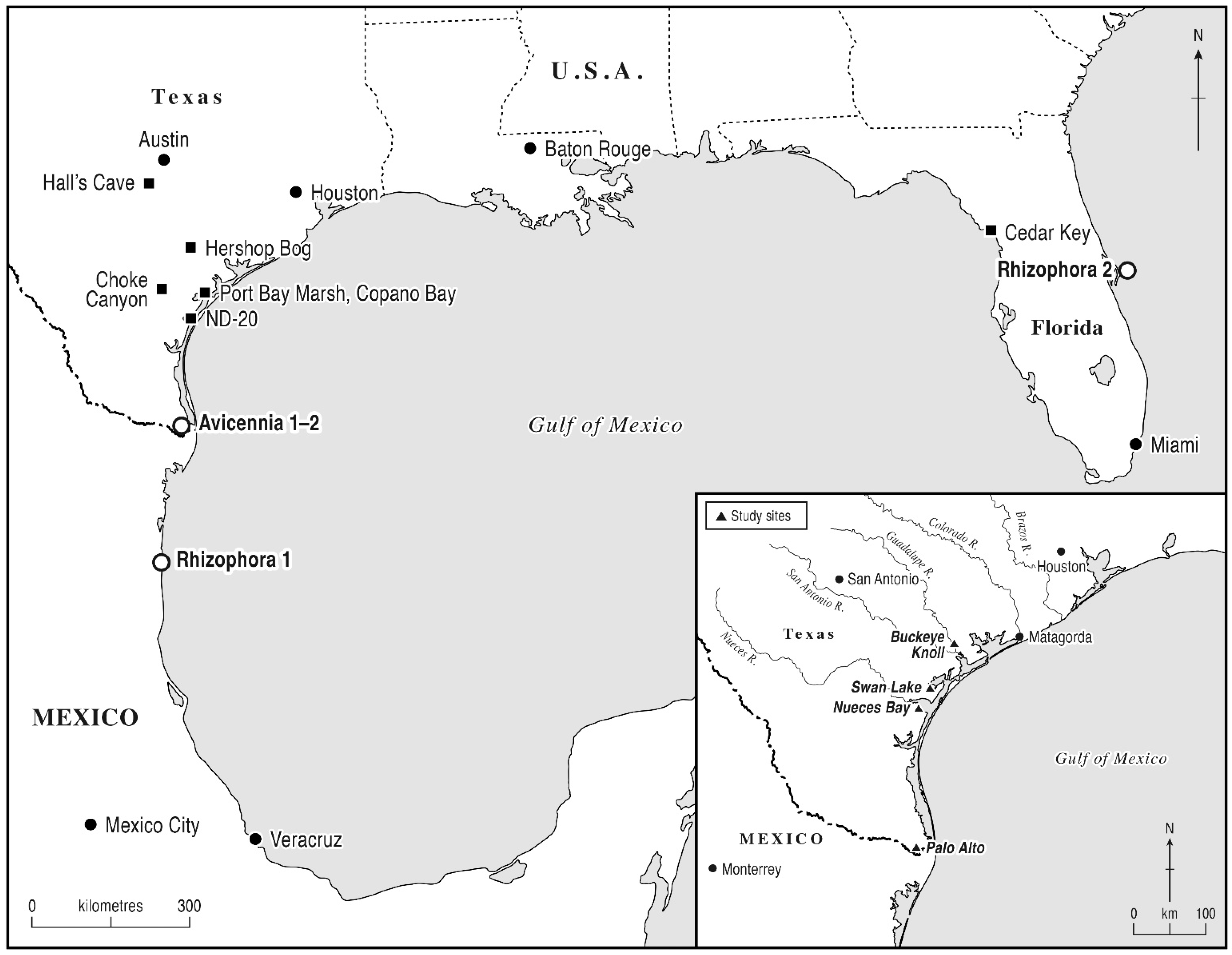
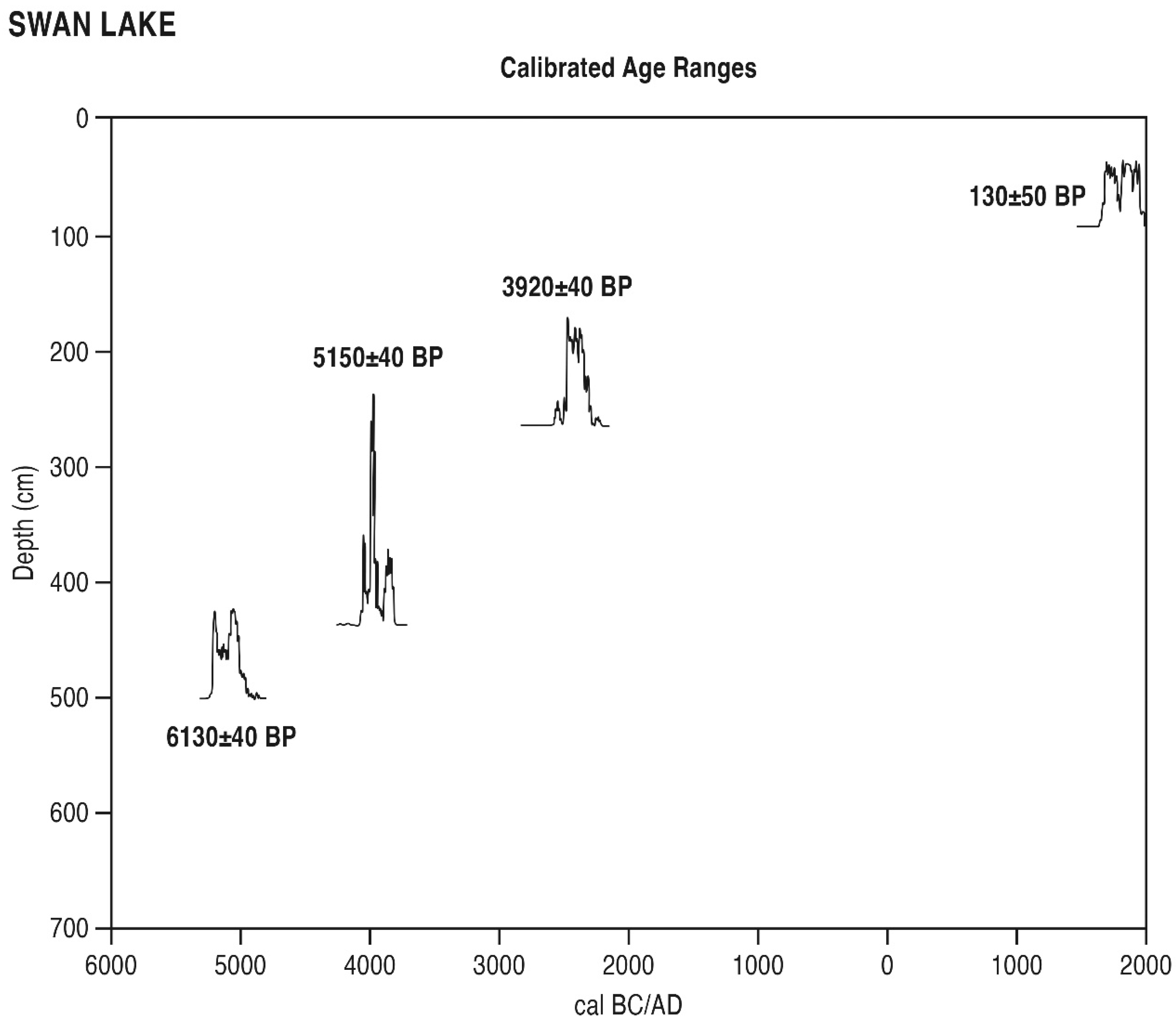
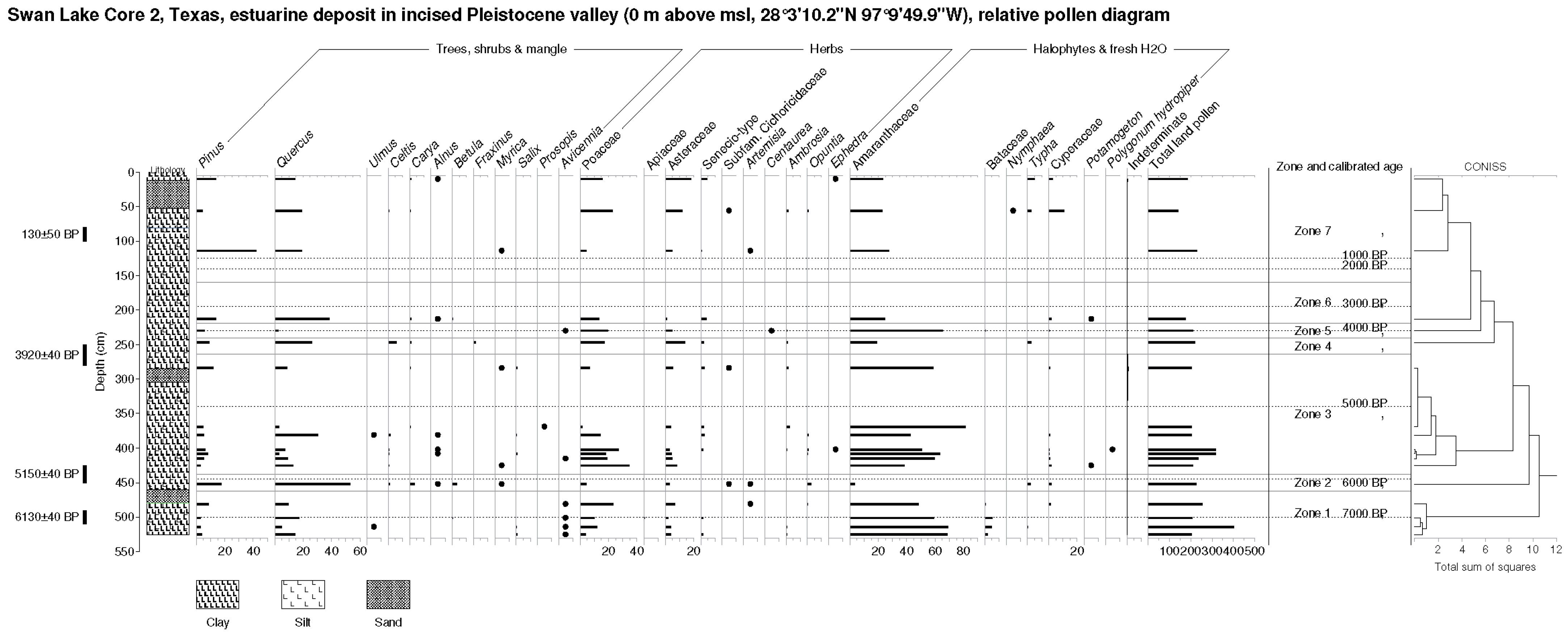
6.2.2. Swan Lake (SL, Figure 6)
SL Zone 1 (481–525 cm, 7100–6650 BP). Relatively low levels of Quercus pollen are observed here, along with isolated Ulmus, Betula, and Salix. A rational representation of Avicennia is observed. Herbaceous pollen consists largely of Amaranthaceae with a significant admixture of Poaceae, Asteraceae, and Bataceae pollen. Freshwater types are represented by isolates only of Typha and Cyperaceae.
SL Zone 2 (452 cm, 5800 BP). Pollen types of trees and shrubs, including, most importantly, Quercus, Pinus, Carya, and Betula, rise in this zone. Isolated Alnus and Myrica also appear here. Herbaceous pollen types include rising Poaceae and Asteraceae. Notable are finds of Opuntia and Artemisia pollen. A rise in Typha and Cyperaceae pollen is observed.
SL Zone 3 (284–425 cm, 5700–4250 BP). Pollen types of trees and shrubs, including, most importantly, Quercus and Carya, decline in this zone. Isolated Ulmus, Celtis, Alnus, Myrica, Salix, Prosopis and Avicennia also appear here. Herbaceous pollen types include rising Poaceae and Asteraceae, along with declining Amaranthaceae. Initial Potamogeton, Polygonum hydropiper and declining Cyperaceae pollen are further observed here.
SL Zone 4 (247 cm, 3650 BP). Pollen types of trees and shrubs, including Quercus and Celtis, rise in this zone. Isolated Fraxinus also appears here. Herbaceous pollen types include rising Poaceae and Asteraceae, along with declining Amaranthaceae. Typha pollen is also observed.
SL Zone 5 (230 cm, 3450 BP). Pollen of trees and shrubs, including, most importantly, Quercus, decline in this zone. Isolated Avicennia also appears. Herbaceous pollen types include Poaceae and Asteraceae, along with rising Amaranthaceae and a Bataceae grain.
SL Zone 6 (213 cm, 3050 BP). Pollen of trees and shrubs, including, most importantly, Pinus and Quercus, rise in this zone. Isolated Celtis and Alnus also appear. Herbaceous pollen types include declining Poaceae and Asteraceae, along with rising Amaranthaceae. Low levels of Typha and an isolated Potamogeton grain are noted.
SL Zone 7 (10–114 cm, 700 BP to recent). Pollen of trees and shrubs declines progressively in this zone. Isolated Alnus and Myrica also appear. Herbaceous pollen types include rising Poaceae and Asteraceae, along with moderate levels of Amaranthaceae and rising terminal values of Typha and Cyperaceae. An isolate of (freshwater) Nymphaea is further noted. Finally, low levels of both Opuntia and Ambrosia are observed here.
Figure 6.
Swan Lake (SL) relative pollen diagram (percentage of TLP) and Troels-Smith lithology. Note redox features and pollen hiatus 130–280 cm and absence of pollen samples in sand layers.
Figure 6.
Swan Lake (SL) relative pollen diagram (percentage of TLP) and Troels-Smith lithology. Note redox features and pollen hiatus 130–280 cm and absence of pollen samples in sand layers.
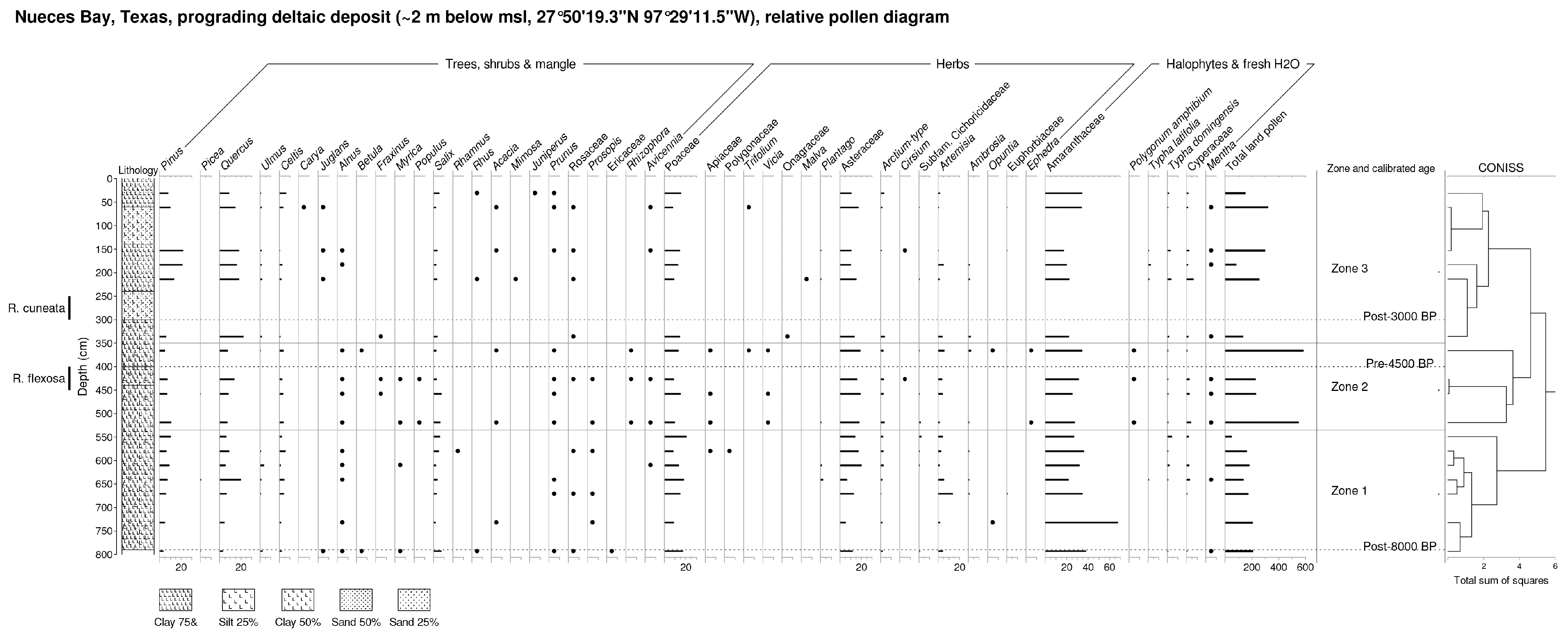
6.2.3. Nueces Bay (NB, Figure 7)
NB Zone 1 (549–793 cm, ~8000–6000 BP). Arboreal pollen (esp. Quercus, Celtis, Ulmus and Salix) rises in this zone. Isolates include Juglans, Alnus, Betula, Myrica, Rhamnus, Rhus, Acacia, Prunus, Prosopis, Ericaceae (cf. Arbutus) and Avicennia (terminally). Herbaceous pollen types include Poaceae and Asteraceae, along with declining Amaranthaceae (site max., at base of sequence) and rising terminal values of Typha and Cyperaceae.
NB Zone 2 (366–519 cm, ~6000–4500 BP). Levels of AP, including Quercus, Celtis and Salix, are stable. In addition to moderate levels of Pinus, isolates of Alnus, Betula, Acacia, Prunus, and Prosopis occur. A near-rational representation of isolates of Avicennia as well as Rhizophora is achieved. Among herbaceous pollen types, Poaceae values decline somewhat, while those of Asteraceae and Amaranthaceae remain stable. Pollen grains of Vicia and Apiaceae are further noted. Freshwater plant pollen now includes Typha, Cyperaceae, Polygonum amphibium and Mentha.
NB Zone 3 (31–336 cm, post-4000 BP). With the exception of Pinus, AP declines, led by Quercus, Celtis and Salix. Isolates of Ulmus, Carya, Juglans, Alnus, Fraxinus, Acacia, Mimosa, Prunus and Avicennia occur. Non-arboreal pollen, led by Poaceae, Asteraceae and Amaranthaceae, are oscillatory. Maxima of Typha and Cyperaceae are noted in the middle part of this zone (~3000 BP).
Figure 7.
Nueces Bay (NB) relative pollen diagram (percentage of TLP) and Troels-Smith lithology. Note that the shellfish stratum with Rangia cuneata is unsampled due to the density of this shellfish bed.
Figure 7.
Nueces Bay (NB) relative pollen diagram (percentage of TLP) and Troels-Smith lithology. Note that the shellfish stratum with Rangia cuneata is unsampled due to the density of this shellfish bed.
6.2.4. Definition of the Forest–Prairie Ecotone
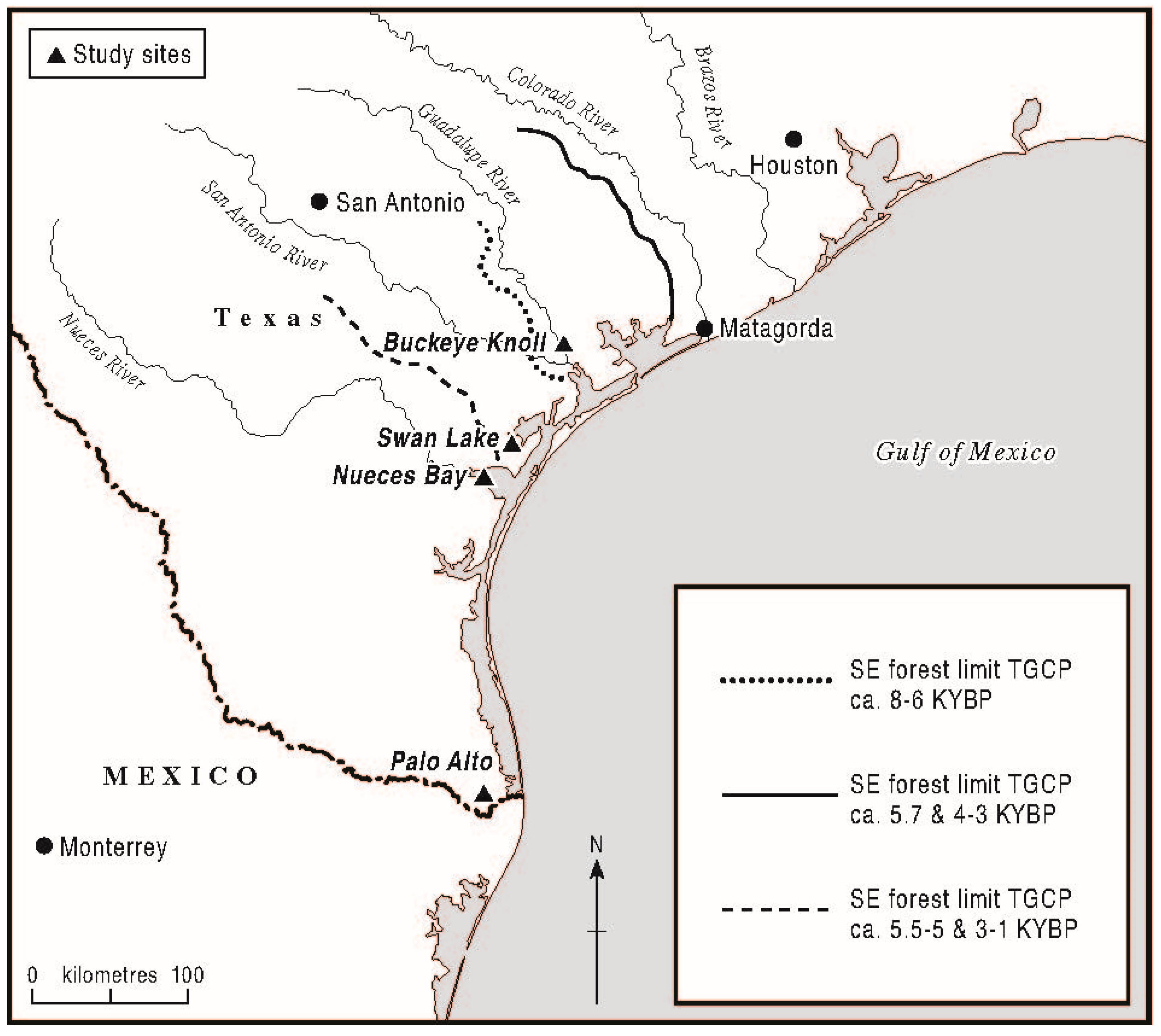
In the reconstruction of the past, the forest–prairie vegetation changes in the TGCP; a definition of the regional forest–prairie ecotone according to pollen statistics is necessary. In this section, the most recent, top-set samples of the examined pollen sites and the supplemental site of Palo Alto at the southern limit of the TGCP are employed (as analogs) to define the forest–prairie ecotone in terms of AP percentage levels. The sum of AP is calculated as all tree and shrub pollen minus (long-distance)
Pinus pollen. These samples were used to statistically define ideal vegetation types, with error bands of the AP level defined by upper confidence limits of AP (%) at prairie sites and the lower confidence limit of AP (%) at the forest site (BK). According to pollen analyses (
Section 6.2.1,
Section 6.2.2 and
Section 6.2.3, please see the Palo Alto pollen diagram
supplemental file), the top-set samples at BK, SL, NB and Palo Alto produced AP values ranging from 36.5% at BK in the north to 3.5% at Palo Alto in the south. Notably, top-set AP levels at SL and NB, within 13 min latitude of each other, produced near-equivalent AP levels, with a uniform trend towards higher AP at higher latitudes being evident (small sample number, low statistical noise;
Table 8). The sample at SL was chosen to establish expected AP levels in the prairie environment near the ecotone because of its higher pollen count (smaller confidence interval) and biogeography (prairie sample in closest proximity to forested zone). The confidence interval upper boundary here was 22.4% AP, while the lower boundary of BK (sole forest zone site) was 30.0% (
bold). According to these data, it was inferred that an AP above 30.0% indicates a forest vegetation type in paleo-pollen spectra. Below 22.4% AP, a prairie vegetation type is indicted, while intermediate AP levels (22.4–30.0%) are deemed transitional or indeterminate as to vegetation type and contain the theoretical ecotone. The application of these results is discussed below (
Section 7.1) in an assessment of Holocene climate changes according to earlier sediment core pollen data and comparative (
Section 3) data.
6.2.5. Results of Mangrove Pollen Analog Studies
Analogs for mangrove pollen of
Avicennia and
Rhizophora species from the Gulf of Mexico and the Central Florida Atlantic coast demonstrate that low levels of mangrove pollen can indicate a local presence (
Table 9), conformably with the poor dispersion of most insect pollinated species. In general, pollen from the analog sites was dominated by the local herbaceous halophytic taxa (esp. Bataceae and Amaranthaceae) or (in the case of Orchid Island, Florida) other arboreal taxa of the floodplain. In the case of the Orchid Island sample (
Rhizophora 2), a major pollen taphonomic influence of the Indian River (regional hydrological transport) was inferred according to the high pollen percentages of the mixed oak–pine forest (absent from the local sampling site). From La Pesca data at the northern climatic limit of
Rhizophora in Mexico (
Rhizophora 1),
Rhizophora mangle pollen was found to be absent (comparatively, the pollen response of
Rhizophora increased markedly in lower-latitude coastal regions of Mexico [
53]). In the case of sampled
Avicennia sites from the Laguna Atascosa Wildlife Refuge in the Rio Grande valley,
Avicennia pollen percentages approached 2% TLP in both samples, although the sample from the central part of the mangrove stand (
Avicennia 2) contained greater quantities of Bataceae, inferably due to greater local, non-anemophilous pollen influence with lower wind velocities. According to these analogs, isolated mangrove pollen grains at the SL and NB sites may thus be interpreted as small mangles representing vegetation patches in the order of one to ten hectares. An absence of mangrove pollen, however, was expected where local mangles were small, in the order of one hectare or less.
In the case of NB (Zone 2), a presence of isolated grains of
Rhizophora pollen indicated the local presence of this fully tropical mangrove and thus a boreal extension of its climatic limit. This inference further indicates a tropical Middle Holocene climate in the central TGCP circa 6000 to 4500 BP, according to biostratigraphic dating of proximal
Rangia flexuosa fortuitously encountered in the NB pollen core. Importantly,
R. flexuosa itself enjoys a tropical distribution today in the Gulf Coast of Mexico, supporting the interpretation of the analog data by virtue of an analogous climatic range change (see
Section 7.1).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}