Do Mixed Fire Regimes Shape Plant Flammability and Post-Fire Recovery Strategies?



Abstract
:1. Introduction
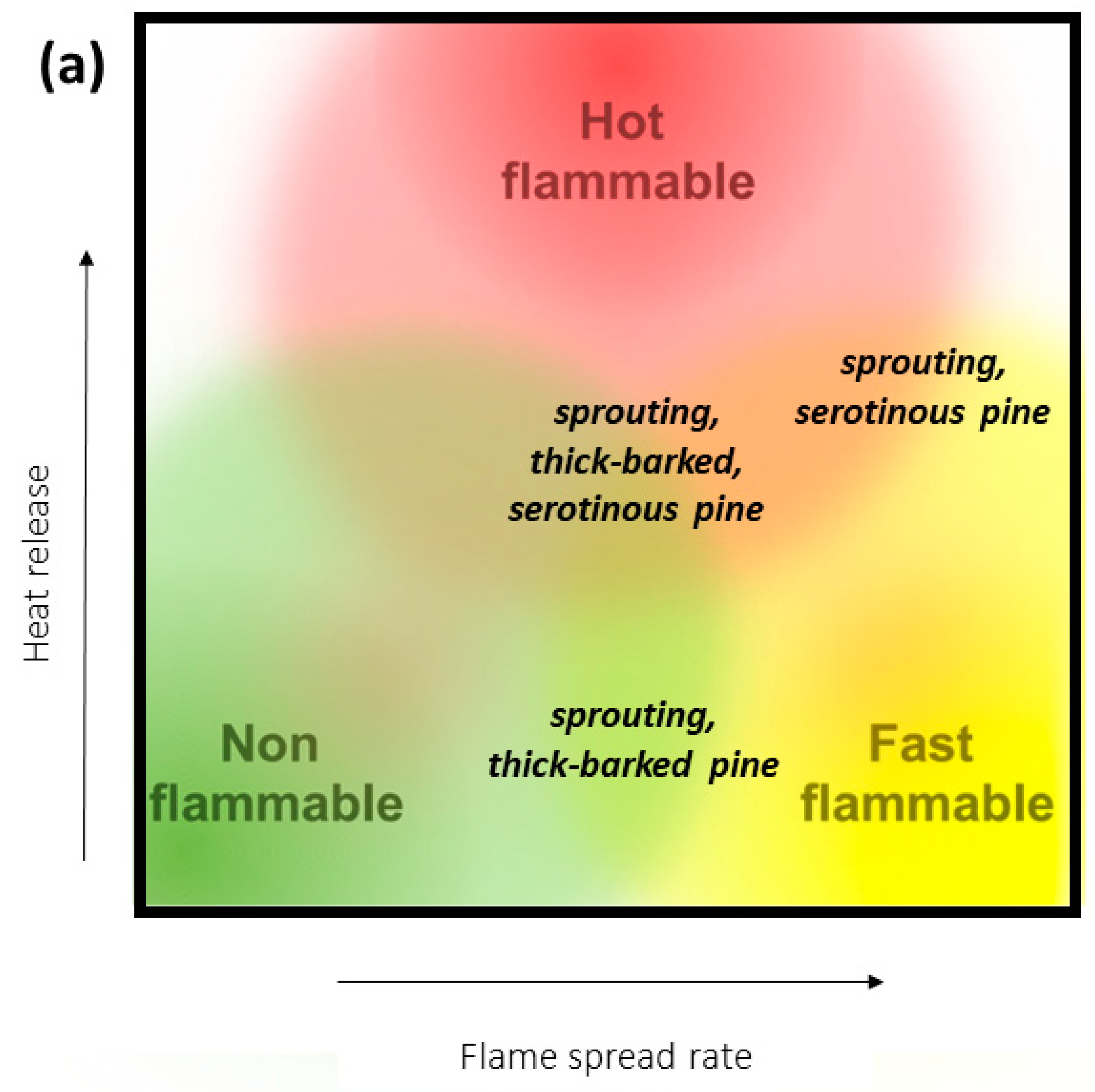
2. Species Exhibiting Multiple Flammability Strategies
3. Case Studies
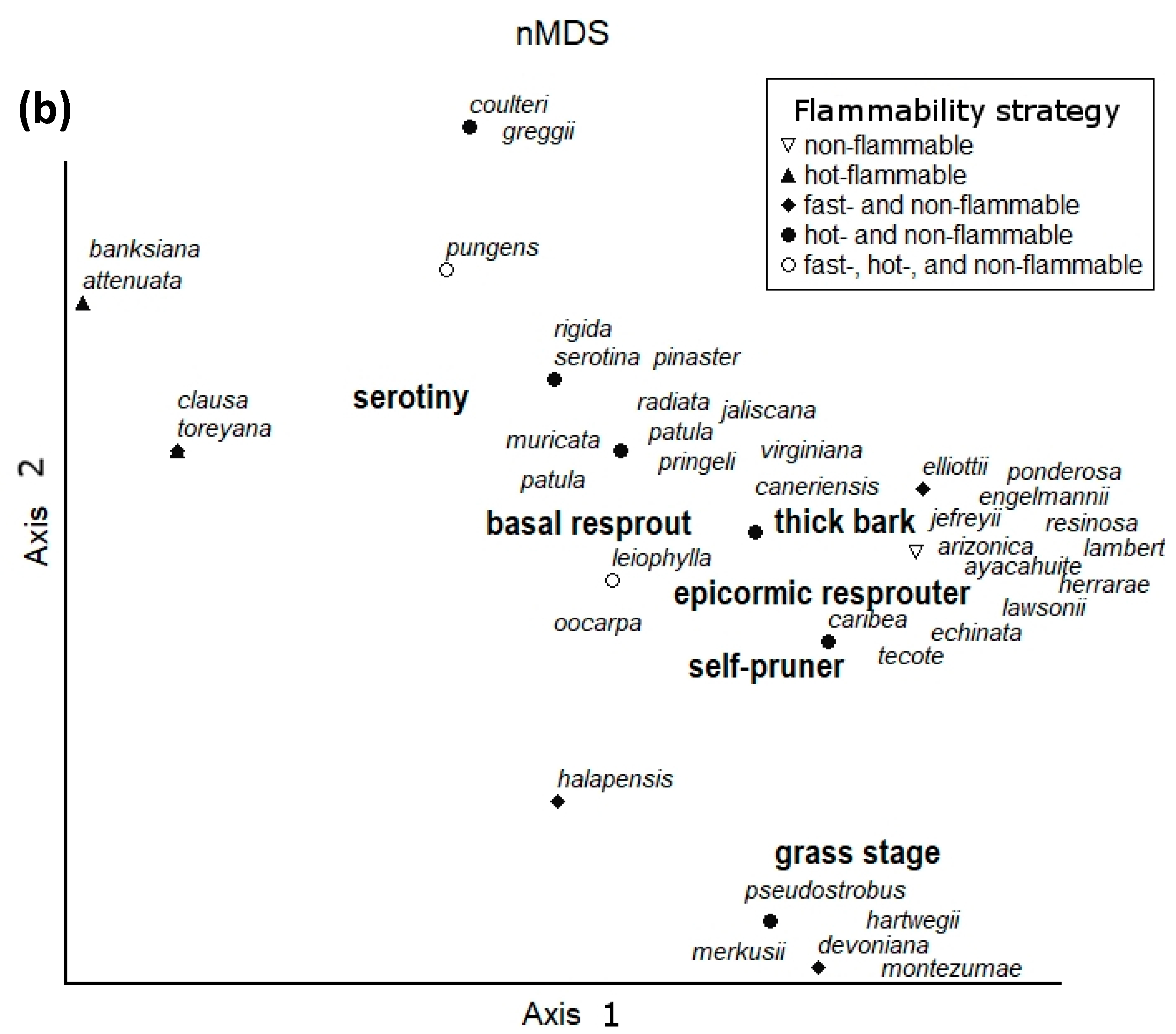
3.1. Pinus
3.2. Proteaceae of the Cape Floristic Region, South Africa
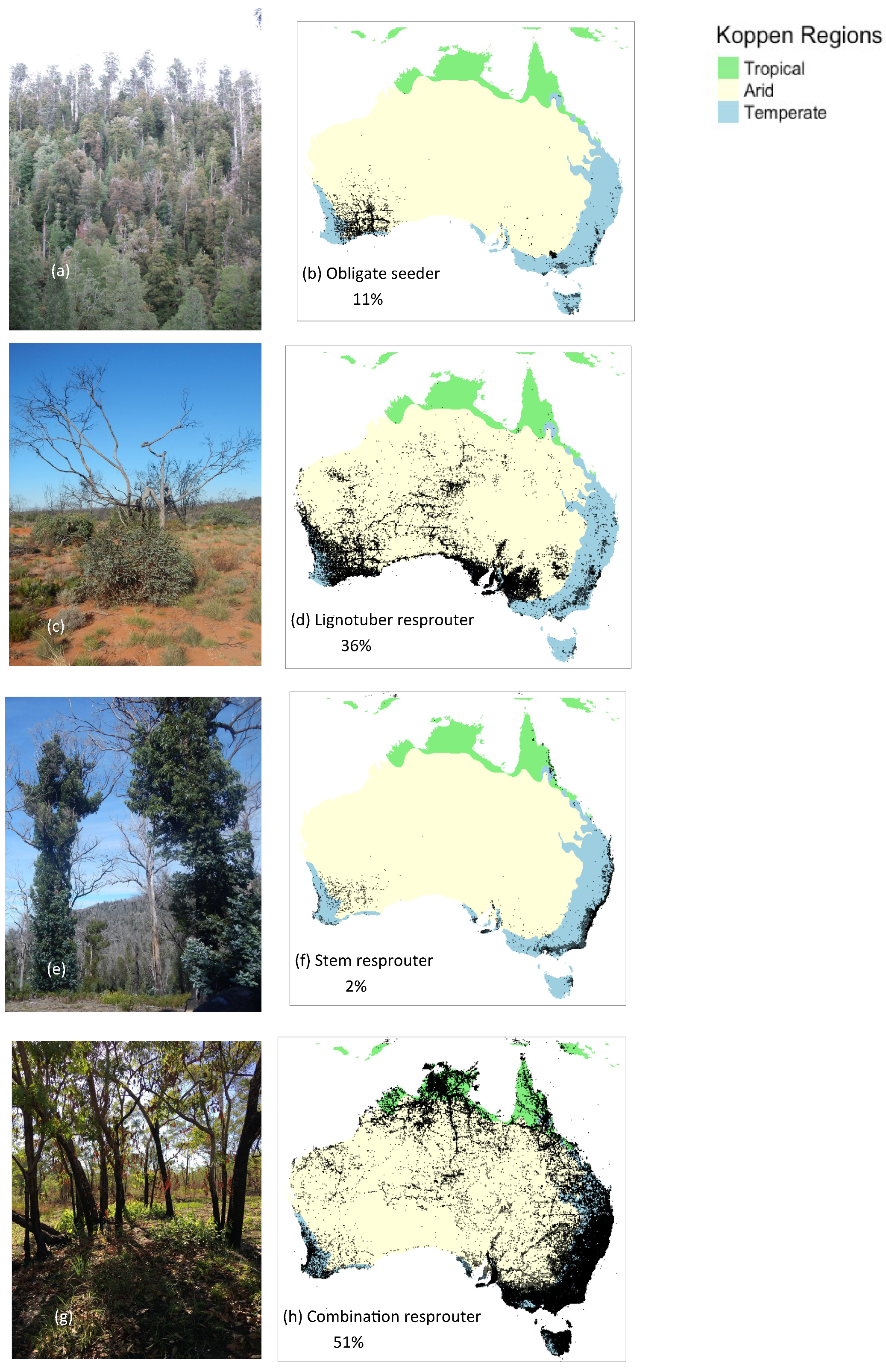
3.3. Eucalyptus
4. Mixed-Fire Regimes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerling, A.L. Increasing western us forest wildfire activity: Sensitivity to changes in the timing of spring. Philos. Trans. R. Soc. B 2016, 371, 20150178. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Williamson, G.J.; Abatzoglou, J.T.; Kolden, C.A.; Cochrane, M.A.; Smith, A.M.S. Human exposure and sensitivity to globally extreme wildfire events. Nat. Ecol. Evol. 2017, 1, 0058. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.J.; Midgley, J.J. Fire and the angiosperm revolutions. Int. J. Plant Sci. 2012, 173, 569–583. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. The evolutionary ecology of sprouting in woody plants. Int. J. Plant Sci. 2003, 164, S103–S114. [Google Scholar] [CrossRef]
- He, T.; Pausas, J.G.; Belcher, C.M.; Schwilk, D.W.; Lamont, B.B. Fire-adapted traits of pinus arose in the fiery cretaceous. New Phytol. 2012, 194, 751–759. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Belcher, C.M.; Lamont, B.B.; Lim, S.L. A 350-million-year legacy of fire adaptation among conifers. J. Ecol. 2016, 104, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Bell, D.T. Ecological response syndromes in the flora of southwestern Western Australia: Fire resprouters versus reseeders. Bot. Rev. 2001, 67, 417–440. [Google Scholar] [CrossRef]
- Verdú, M. Ecological and evolutionary differences between Mediterranean seeders and resprouters. J. Veg. Sci. 2000, 11, 265–268. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C. Role of fire in regeneration from seed. In Seeds: The Ecology of Regeneration in Plant Communities; CABI Publishing: Wallingford, UK, 2000; pp. 311–330. [Google Scholar]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Badik, K.J.; Jahner, J.P.; Wilson, J.S. A biogeographic perspective on the evolution of fire syndromes in pine trees (pinus: Pinaceae). R. Soc. Open Sci. 2018, 5, 172412. [Google Scholar] [CrossRef] [PubMed]
- Schwilk, D.W.; Ackerly, D.D. Flammability and serotiny as strategies: Correlated evolution in pines. Oikos 2001, 94, 326–336. [Google Scholar] [CrossRef]
- Bradshaw, S.D.; Dixon, K.W.; Hopper, S.D.; Lambers, H.; Turner, S.R. Response to Keeley et al.: Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 405. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutch, R.W. Wildland fires and ecosystems—A hypothesis. Ecology 1970, 51, 1046–1051. [Google Scholar] [CrossRef]
- Rowe, J.S. Concepts of fire effects on plant individuals and species. In The Role of Fire in Northern Circumpolar Ecosystems: Scientific Committee on Problems of the Environment; Wein, R.W., MacLean, D.A., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 1983; pp. 135–180. [Google Scholar]
- Schwilk, D.W. Dimensions of plant flammability. New Phytol. 2015, 206, 486–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pausas, J.G.; Keeley, J.E.; Schwilk, D.W. Flammability as an ecological and evolutionary driver. J. Ecol. 2017, 105, 289–297. [Google Scholar] [CrossRef]
- Krawchuk, M.A.; Haire, S.L.; Coop, J.; Parisien, M.A.; Whitman, E.; Chong, G.; Miller, C. Topographic and fire weather controls of fire refugia in forested ecosystems of northwestern north America. Ecosphere 2016, 7, e01632. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.; Gómez-Dans, J.L.; Bradstock, R.A. Defining pyromes and global syndromes of fire regimes. Proc. Natl. Acad. Sci. USA 2013, 201211466. [Google Scholar] [CrossRef] [PubMed]
- Archibald, S.; Lehmann, C.E.; Belcher, C.; Bond, W.J.; Bradstock, R.A.; Daniau, A.; Dexter, K.; Forrestel, E.; Greve, M.; He, T.; et al. Biological and geophysical feedbacks with fire in the earth system. Environ. Res. Lett. 2018, 13, 033003. [Google Scholar] [CrossRef]
- Kasischke, E.S.; Turetsky, M.R. Recent changes in the fire regime across the North American boreal region—Spatial and temporal patterns of burning across Canada and Alaska. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef] [Green Version]
- Halofsky, J.; Donato, D.; Hibbs, D.; Campbell, J.; Cannon, M.D.; Fontaine, J.; Thompson, J.R.; Anthony, R.; Bormann, B.; Kayes, L.; et al. Mixed-severity fire regimes: Lessons and hypotheses from the Klamath-Siskiyou ecoregion. Ecosphere 2011, 2, 1–19. [Google Scholar] [CrossRef]
- Kane, V.R.; Cansler, C.A.; Povak, N.A.; Kane, J.T.; McGaughey, R.J.; Lutz, J.A.; Churchill, D.J.; North, M.P. Mixed severity fire effects within the rim fire: Relative importance of local climate, fire weather, topography, and forest structure. For. Ecol. Manag. 2015, 358, 62–79. [Google Scholar] [CrossRef]
- Keeley, J.E.; Zedler, P.H. Evolution of life histories in pinus. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: New York, NY, USA, 2000; pp. 219–251. [Google Scholar]
- Barton, A.M. Intense wildfire in southeastern Arizona: Transformation of a Madrean oak–pine forest to oak woodland. For. Ecol. Manag. 2002, 165, 205–212. [Google Scholar] [CrossRef]
- Barton, A.M. Pines versus oaks: Effects of fire on the composition of Madrean forests in Arizona. For. Ecol. Manag. 1999, 120, 143–156. [Google Scholar] [CrossRef]
- Bennett, P.S.; Kunzmann, M.R. The applicability of generalized fire prescriptions to burning of madrean evergreen forest and woodland. J. Arizona-Nevada Acad. Sci. 1992, 24–25, 79–84. [Google Scholar]
- Ffolliott, P.F.; Gottfried, G.J.; Bennett, D.A.; Ortega-Rubio, A.; Hamre, R. Tech. Coords. 1992. Ecology and Management of Oak and Associated Woodlands: Perspectives in the Southwestern United States and Northern Mexico; 1992 April 27–30; Sierra Vista, AZ; Gen. Tech. Rep. RM-GTR-218; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1992; p. 224.
- Preston, R.J. North American Trees; Iowa State College Press: Ames, IA, USA, 1948. [Google Scholar]
- Barden, L.S. Serotiny and seed viability of pinus pungens in the southern Appalachians. Castanea 1979, 1, 44–47. [Google Scholar]
- Mohr, H.H.; Waldrop, T.A.; Shelburne, V.B. Optimal Seedbed Requirements for Regenerating Table Mountain Pine; Gen. Tech. Rep. SRS-48; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 306–309.
- Williams, C.E. History and status of table mountain pine–pitch pine forests of the southern Appalachian Mountains (USA). Nat. Areas J. 1998, 18, 81–90. [Google Scholar]
- Stone, E.; Stone, M. Root collar sprouts in pine. J. For. 1954, 52, 487–491. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, version 6; MjM Software: Gleneden Beach, OR, USA, 2011. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorites. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A. Saso Proteas: A Field Guide to the Proteas of Southern Africa; Fernwood Press in Association with the National Botanical Institute: Vlaeberg, South Africa, 2001; p. 240. [Google Scholar]
- Thuiller, W.; Slingsby, J.A.; Privett, S.D.; Cowling, R.M. Stochastic species turnover and stable coexistence in a species-rich, fire-prone plant community. PLoS ONE 2007, 2, e938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, B.B.; Le Maitre, D.; Cowling, R.; Enright, N. Canopy seed storage in woody plants. Bot. Rev. 1991, 57, 277–317. [Google Scholar] [CrossRef]
- Tonnabel, J.; Schurr, F.M.; Boucher, F.; Thuiller, W.; Renaud, J.; Douzery, E.J.; Ronce, O. Life-history traits evolved jointly with climatic niche and disturbance regime in the genus leucadendron (proteaceae). Am. Nat. 2017, 191, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E. Epicormic resprouting in fire-prone ecosystems. Trends Plant Sci. 2017, 22, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Climent, J.; Chambel, M.R.; López, R.; Mutke, S.; Alía, R.; Gil, L. Population divergence for heteroblasty in the canary island pine (Pinus canariensis, pinaceae). Am. J. Bot. 2006, 93, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Trejo, D.A.; Fulé, P.Z. Fire ecology of Mexican pines and a fire management proposal. Int. J. Wildland Fire 2003, 12, 23–37. [Google Scholar] [CrossRef]
- Viveros-Viveros, H.; Sáenz-Romero, C.; Vargas-Hernández, J.J.; López-Upton, J.; Ramírez-Valverde, G.; Santacruz-Varela, A. Altitudinal genetic variation in Pinus hartwegii Lindl. I: Height growth, shoot phenology, and frost damage in seedlings. For. Ecol. Manag. 2009, 257, 836–842. [Google Scholar] [CrossRef]
- Lamant, T. Vegetative reproduction in gymnosperms. J. l’Association des Parcs Botaniques de France 2012, 53, 5. [Google Scholar]
- Burns, R.M.; Barbara, H. Silvics of North America: 1. Conifers; 2. Hardwoods; Agriculture Handbook 654; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 2, p. 877.
- Ketcham, D.; Bethune, J. Fire resistance of south Florida slash pine. J. For. 1963, 61, 529–530. [Google Scholar]
- Tapias, R.; Climent, J.; Pardos, J.A.; Gil, L. Life histories of Mediterranean pines. Plant Ecol. 2004, 171, 53–68. [Google Scholar] [CrossRef]
- Ne’eman, G.; Goubitz, S.; Nathan, R. Reproductive traits of Pinus halepensis in the light of fire—A critical review. Plant Ecol. 2004, 171, 69–79. [Google Scholar] [CrossRef]
- Daskalakou, E.N.; Thanos, C.A. Aleppo pine (Pinus halepensis) postfire regeneration: The role of canopy and soil seed banks. Int. J. Wildland Fire 1996, 6, 59–66. [Google Scholar] [CrossRef]
- Koskela, J. Early shoot and diameter growth in four Pinus merkusii populations from Thailand. J. Trop. For. Sci. 2000, 12, 707–716. [Google Scholar]
- Andersson, F. Coniferous Forests; Elsevier: San Diego, CA, USA, 2005; Volume 6. [Google Scholar]
- Farjon, A. A Handbook of the World’s Conifers (2 Vols.); Brill Academic Publisher: Boston, MA, USA, 2010. [Google Scholar]
- Fernandes, P.M.; Rigolot, E. The fire ecology and management of maritime pine (Pinus pinaster ait.). For. Ecol. Manag. 2007, 241, 1–13. [Google Scholar] [CrossRef]
- Gil, L.; López, R.; García-Mateos, Á.; González-Doncel, I. Seed provenance and fire-related reproductive traits of Pinus pinaster in central Spain. Int. J. Wildland Fire 2010, 18, 1003–1009. [Google Scholar] [CrossRef]
- Little, S. Prescribed burning as a tool of forest management in the northeastern states. J. For. 1953, 51, 496–500. [Google Scholar]
- Niering, W.A.; Dreyer, G.D. Effects of prescribed burning on andropogon scoparius in postagricultural grasslands in Connecticut. Am. Midl. Nat. 1989, 122, 88–102. [Google Scholar] [CrossRef]
- Crutchfield, D.; Trew, I. Investigation of natural regeneration of pond pine. J. For. 1961, 59, 264–266. [Google Scholar]
- Christensen, N.L. Shrubland fire regimes and their evolutionary consequences. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: London, UK, 1985; pp. 85–100. [Google Scholar]
- Wyse, S.V.; Perry, G.L.; O’Connell, D.M.; Holland, P.S.; Wright, M.J.; Hosted, C.L.; Whitelock, S.L.; Geary, I.J.; Maurin, K.J.; Curran, T.J. A quantitative assessment of shoot flammability for 60 tree and shrub species supports rankings based on expert opinion. Int. J. Wildland Fire 2016, 25, 466–477. [Google Scholar] [Green Version]
- Bamber, R.; Mullette, K. Studies of the lignotubers of eucalyptus gummifera (gaertn. & hochr.). Ii. Anatomy. Aust. J. Bot. 1978, 26, 15–22. [Google Scholar]
- Carrodus, B.; Blake, T. Studies on the lignotubers of eucalyptus obliqua l’heri. I. The nature of the lignotuber. New Phytol. 1970, 69, 1069–1072. [Google Scholar] [CrossRef]
- Crisp, M.D.; Burrows, G.E.; Cook, L.G.; Thornhill, A.H.; Bowman, D.M. Flammable biomes dominated by eucalypts originated at the cretaceous-palaeogene boundary. Nat. Commun. 2011, 2, 193. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.D.; Bristow, M.; Vanclay, J.K. Mixed-species plantations: Prospects and challenges. For. Ecol. Manag. 2006, 233, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Lacey, C.J. Rhizomes in tropical eucalypts and their role in recovery from fire damage. Aust J. Bot. 1974, 22, 29–38. [Google Scholar] [CrossRef]
- Nicolle, D. A classification and census of regenerative strategies in the eucalypts (angophora, corymbia and eucalyptus—Myrtaceae), with special reference to the obligate seeders. Aust. J. Bot. 2006, 54, 391–407. [Google Scholar] [CrossRef]
- Lacey, C.; Whelan, P. Observations on the ecological significance of vegetative reproduction in the katherine-darwin region of the northern territory. Aust. For. 1976, 39, 131–139. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bradstock, R.A.; Boer, M.M.; Carter, J.; Cary, G.J.; Cochrane, M.A.; Fensham, R.J.; Russell-Smith, J.; Williamson, G.J.; Bowman, D.M. Fire regimes of Australia: A pyrogeographic model system. J. Biogeogr. 2013, 40, 1048–1058. [Google Scholar] [CrossRef]
- Tng, D.; Williamson, G.; Jordan, G.; Bowman, D. Giant eucalypts—Globally unique fire-adapted rain-forest trees? New Phytol. 2012, 196, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.A.; Balmer, J.; Kirkpatrick, J. Stand-replacing wildfires? The incidence of multi-cohort and single-cohort eucalyptus regnans and e. Obliqua forests in southern Tasmania. For. Ecol. Manag. 2009, 258, 366–375. [Google Scholar] [CrossRef]
- Bowman, D.M.; Williamson, G.J.; Prior, L.D.; Murphy, B.P. The relative importance of intrinsic and extrinsic factors in the decline of obligate seeder forests. Glob. Ecol. Biogeogr. 2016, 25, 1166–1172. [Google Scholar] [CrossRef] [Green Version]
- Lindenmayer, D.; Cunningham, R.; Donnelly, C.; Franklin, J. Structural features of old-growth Australian montane ash forests. For. Ecol. Manag. 2000, 134, 189–204. [Google Scholar] [CrossRef]
- Gosper, C.R.; Prober, S.M.; Yates, C.J. Continental-scale syntheses of Australian pyromes–misclassification of south-western eucalypt woodlands misinforms management. J. Biogeogr. 2016, 43, 858–861. [Google Scholar] [CrossRef]
- Moore, G.M. The role of lignotubers (basal burls) in the stress recovery of messmate stringybark, eucalyptus obliqua l’herit. Seedlings and its arboricultural implications. Arboricult. J. 2015, 37, 113–125. [Google Scholar] [CrossRef]
- Dutkowski, G.W.; Potts, B.M. Geographic patterns of genetic variation in eucalyptus globulus ssp. Globulus and a revised racial classification. Aust. J. Bot. 1999, 47, 237–263. [Google Scholar]
- Ladiges, P.Y.; Ashton, D. Variation in some Central Victoria populations of eucalyptus viminalis labill. Aust. J. Bot. 1974, 22, 81–102. [Google Scholar] [CrossRef]
- Shugart Jr, H.; Noble, I. A computer model of succession and fire response of the high-altitude eucalyptus forest of the brindabella range, Australian Capital Territory. Aust. J. Bot. 1981, 6, 149–164. [Google Scholar] [CrossRef]
- Bowman, D.M.; Murphy, B.P.; Neyland, D.L.; Williamson, G.J.; Prior, L.D. Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Glob. Chang. Biol. 2014, 20, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Bassett, O.D.; Prior, L.D.; Slijkerman, C.M.; Jamieson, D.; Bowman, D.M. Aerial sowing stopped the loss of alpine ash (eucalyptus delegatensis) forests burnt by three short-interval fires in the alpine national park, Victoria, Australia. For. Ecol. Manag. 2015, 342, 39–48. [Google Scholar] [CrossRef]
- Doherty, M.D.; Gill, A.M.; Cary, G.J.; Austin, M.P. Seed viability of early maturing alpine ash (eucalyptus delegatensis subsp. Delegatensis) in the Australian Alps, south-eastern Australia, and its implications for management under changing fire regimes. Aust. J. Bot. 2017, 65, 517–523. [Google Scholar] [CrossRef]
- Bowman, D.; Kirkpatrick, J. Establishment, suppression and growth of eucalyptus delegatensis r.T. Baker in multiaged forests. I. The effects of fire on mortality and seedling establishment. Aust. J. Bot. 1986, 34, 63–72. [Google Scholar] [CrossRef]
- Ellis, R. The relationships among eucalypt forest, grassland and rainforest in a highland area in north-eastern Tasmania. Aust. J. Ecol. 1985, 10, 297–314. [Google Scholar] [CrossRef]
- Kolden, C.A.; Bleeker, T.; Smith, A.; Poulos, H.M.; Camp, A.E. Fire effects on historical wildfire refugia in contemporary wildfires. Forests 2017, 8, 400. [Google Scholar] [CrossRef]
- Enright, N.; Keith, D.A.; Clarke, M.F.; Miller, B.P. Fire regimes in Australian sclerophyllous shrubby ecosystems: Heathlands, heathy woodlands. In Flammable Australia: Fire Regimes and Biodiversity in a Changing World; CSIRO Publishing: Collingwood, Australia, 2012; pp. 215–235. [Google Scholar]
- Gonzalez, M.E.; Veblen, T.T.; Sibold, J.S. Influence of fire severity on stand development of araucaria araucana–Nothofagus pumilio stands in the Andean cordillera of south-central Chile. Aust. Ecol. 2010, 35, 597–615. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Heinlein, T.A.; Moore, M.M.; Covington, W.W.; Verkamp, G. Mixed-severity fire regime in a high-elevation forest of Grand Canyon, Arizona, USA. Landsc. Ecol. 2003, 18, 465–486. [Google Scholar] [CrossRef]
- Cohn, J.S.; Di Stefano, J.; Christie, F.; Cheers, G.; York, A. How do heterogeneity in vegetation types and post-fire age-classes contribute to plant diversity at the landscape scale? For. Ecol. Manag. 2015, 346, 22–30. [Google Scholar] [CrossRef]
- Parr, C.L.; Andersen, A.N. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv. Biol. 2006, 20, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Lawes, M.J.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Lamont, B.B. Baptism by fire: The pivotal role of ancient conflagrations in evolution of the earth’s flora. Nat. Sci. Rev. 2017, 5, 237–254. [Google Scholar] [CrossRef]
- Lamont, B.B.; Enright, N.J.; He, T. Fitness and evolution of resprouters in relation to fire. Plant Ecol. 2011, 212, 1945–1957. [Google Scholar] [CrossRef]
- Perry, D.A.; Hessburg, P.F.; Skinner, C.N.; Spies, T.A.; Stephens, S.L.; Taylor, A.H.; Franklin, J.F.; McComb, B.; Riegel, G. The ecology of mixed severity fire regimes in Washington, Oregon, and northern California. For. Ecol. Manag. 2011, 262, 703–717. [Google Scholar] [CrossRef]
- Baker, W.L. Transitioning western us dry forests to limited committed warming with bet-hedging and natural disturbances. Ecosphere 2018, 9, e02288. [Google Scholar] [CrossRef]
- Barga, S.; Dilts, T.E.; Leger, E.A. Climate variability affects the germination strategies exhibited by arid land plants. Oecologia 2017, 185, 437–452. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Lamont, B.B.; Enright, N.J.; D’Agui, H.M.; Stock, W. Environmental drivers and genomic architecture of trait differentiation in fire-adapted banksia attenuata ecotypes. J. Integr. Plant Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Radeloff, V.C.; Mladenoff, D.J.; Guries, R.P.; Boyce, M.S. Spatial patterns of cone serotiny in pinus banksiana in relation to fire disturbance. For. Ecol. Manag. 2004, 189, 133–141. [Google Scholar] [CrossRef]
- Muir, P.S.; Lotan, J.E. Disturbance history and serotiny of pinus contorta in western Montana. Ecology 1985, 66, 1658–1668. [Google Scholar] [CrossRef]
- Givnish, T.J. Serotiny, geography, and fire in the pine barrens of New Jersey. Evolution 1981, 35, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Ledig, F.T.; Fryer, J.H. A pocket of variability in pinus rigida. Evolution 1972, 26, 259–266. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Taxon | Grass Stage | Basal Resprout | Epicormic Resprout | Seed Banker | Serotiny | Thick Bark | Self-Pruning | Flammability Strategy | Source |
|---|---|---|---|---|---|---|---|---|---|
| (A) Pinaceae | |||||||||
| Pinus canariensis | + | + | + | - | - | + | - | fast and non-flammable | [26,42,43] |
| P. caribaea var. hondurensis | - | + | + | - | - | + | + | fast and non-flammable | [6,44] |
| P. devoniana | ± | - | - | - | - | + | + | fast and non-flammable | [26,45] |
| P. echinata | - | + | + | - | - | + | + | fast and non-flammable | [46] |
| P. elliottii var. densa | - | + | - | - | - | + | + | fast and non-flammable | [13,47,48] |
| P. hartwegii | ± | + | + | - | - | + | + | fast, hot, and non-flammable | [44,46] |
| P. halapensis | ± | - | - | - | ± | + | + | fast and hot-flammable | [49,50,51] |
| P. leiophylla | - | + | + | - | ± | + | + | fast, hot, and non-flammable | [27,28,29,30] |
| P. merkusii | ± | - | - | - | - | + | + | fast and non-flammable | [26,52] |
| P. montezumae | ± | - | - | - | - | + | + | fast and non-flammable | [26,53] |
| P. oocarpa | - | + | + | - | ± | + | + | fast, hot and non-flammable | [26,54] |
| P. patula | - | - | - | - | ± | + | + | fast and hot-flammable | [26,44] |
| P. pinaster | - | - | - | - | ± | + | + | fast and non-flammable | [49,55,56] |
| P. pseudostrobus | ± | + | + | - | - | + | + | fast, hot, and non-flammable | [6,44,46] |
| P. pungens | - | + | - | - | ± | + | - | fast, hot, and non-flammable | [26,32,33,34,35] |
| P. rigida | - | + | + | - | ± | + | - | fast and non-flammable | [26,57] |
| P. serotina | - | + | + | - | ± | + | - | fast, hot, and non-flammable | [26,58,59,60] |
| P. tecote | - | + | + | - | - | + | + | fast, hot, and non-flammable | [6,44,46] |
| P. torreyana | - | - | - | - | + | + | - | fast and non-flammable | [26] |
| P. virginiana | - | + | + | - | - | + | - | fast and non-flammable | [6,26] |
| (B) Cape Floristic Region Proteaceae | |||||||||
| Aulax pallasia | - | + | - | - | + | - | - | fast and hot flammable | [38] |
| Leucospermum cuneiforme | - | + | - | + | - | - | - | fast and hot flammable | [38] |
| Leucospermum conocarpodendron | - | - | - | + | - | + | - | hot and non-flammable | [38] |
| Mimetes fimbriifolious | - | - | - | + | - | + | - | hot and non-flammable | [38] |
| Mimetes cucculatus | - | + | - | + | - | - | - | fast and hot flammable | [38] |
| Leucadendron salignum | - | + | - | - | + | - | - | fast and hot flammable | [38] |
| Leucadendron argenteum | - | - | - | - | + | + | - | hot and non-flammable | [38] |
| Protea cynaroides | - | + | - | - | + | - | - | fast and hot flammable | [38] |
| Protea nitida | - | - | + | - | - | + | - | fast and non-flammable | [38] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poulos, H.M.; Barton, A.M.; Slingsby, J.A.; Bowman, D.M.J.S. Do Mixed Fire Regimes Shape Plant Flammability and Post-Fire Recovery Strategies? Fire 2018, 1, 39. https://doi.org/10.3390/fire1030039
Poulos HM, Barton AM, Slingsby JA, Bowman DMJS. Do Mixed Fire Regimes Shape Plant Flammability and Post-Fire Recovery Strategies? Fire. 2018; 1(3):39. https://doi.org/10.3390/fire1030039
Chicago/Turabian StylePoulos, Helen M., Andrew M. Barton, Jasper A. Slingsby, and David M.J.S. Bowman. 2018. "Do Mixed Fire Regimes Shape Plant Flammability and Post-Fire Recovery Strategies?" Fire 1, no. 3: 39. https://doi.org/10.3390/fire1030039
APA StylePoulos, H. M., Barton, A. M., Slingsby, J. A., & Bowman, D. M. J. S. (2018). Do Mixed Fire Regimes Shape Plant Flammability and Post-Fire Recovery Strategies? Fire, 1(3), 39. https://doi.org/10.3390/fire1030039