Post-Fire Carbon Dynamics in Subalpine Forests of the Rocky Mountains

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Model Description
2.2. Study Sites and Data Collection
2.3. Model Inputs and Parameterization
2.4. Model Simulation Scenarios
2.5. Model Evaluation and Statistical Analyses
3. Results
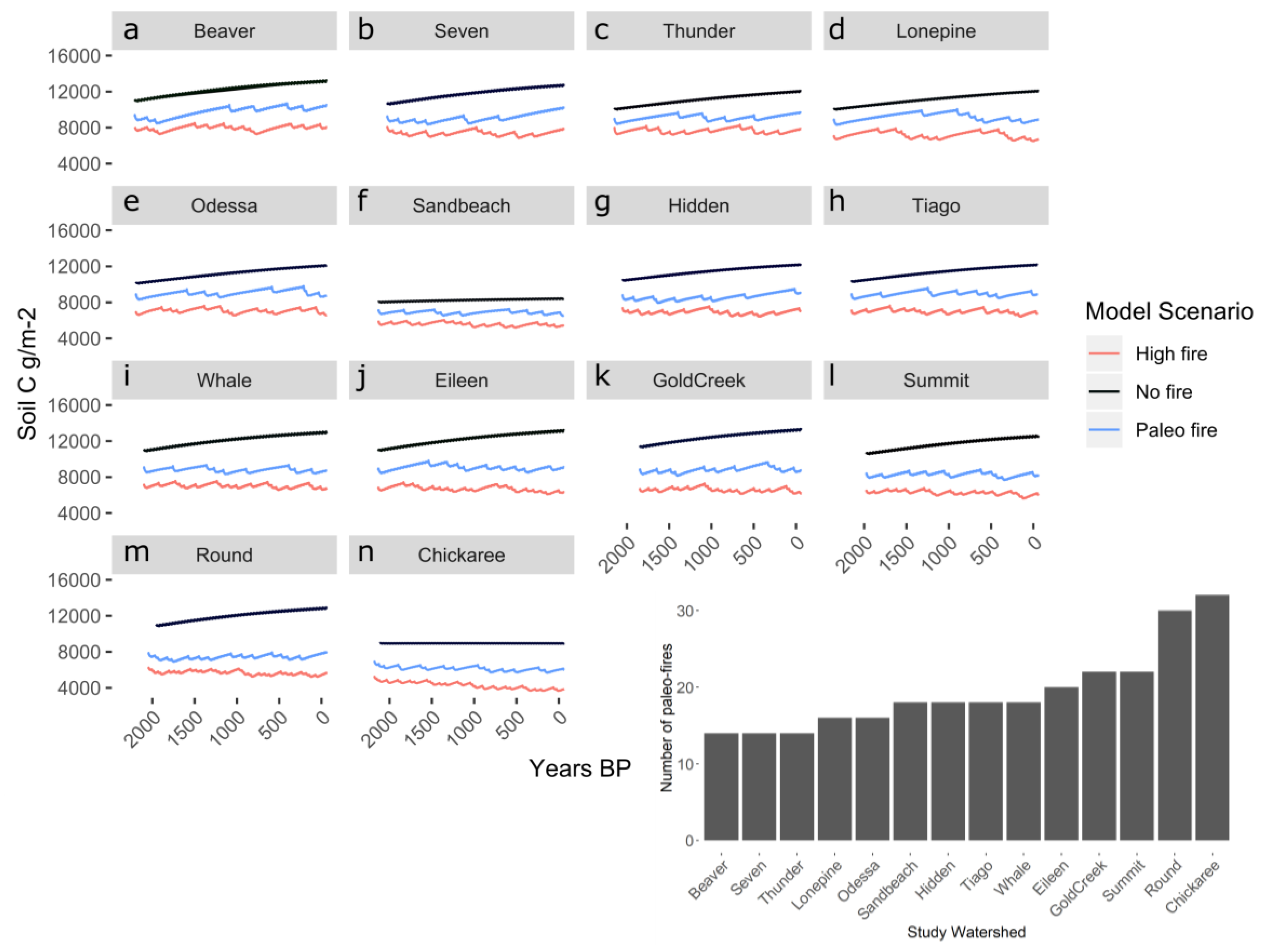
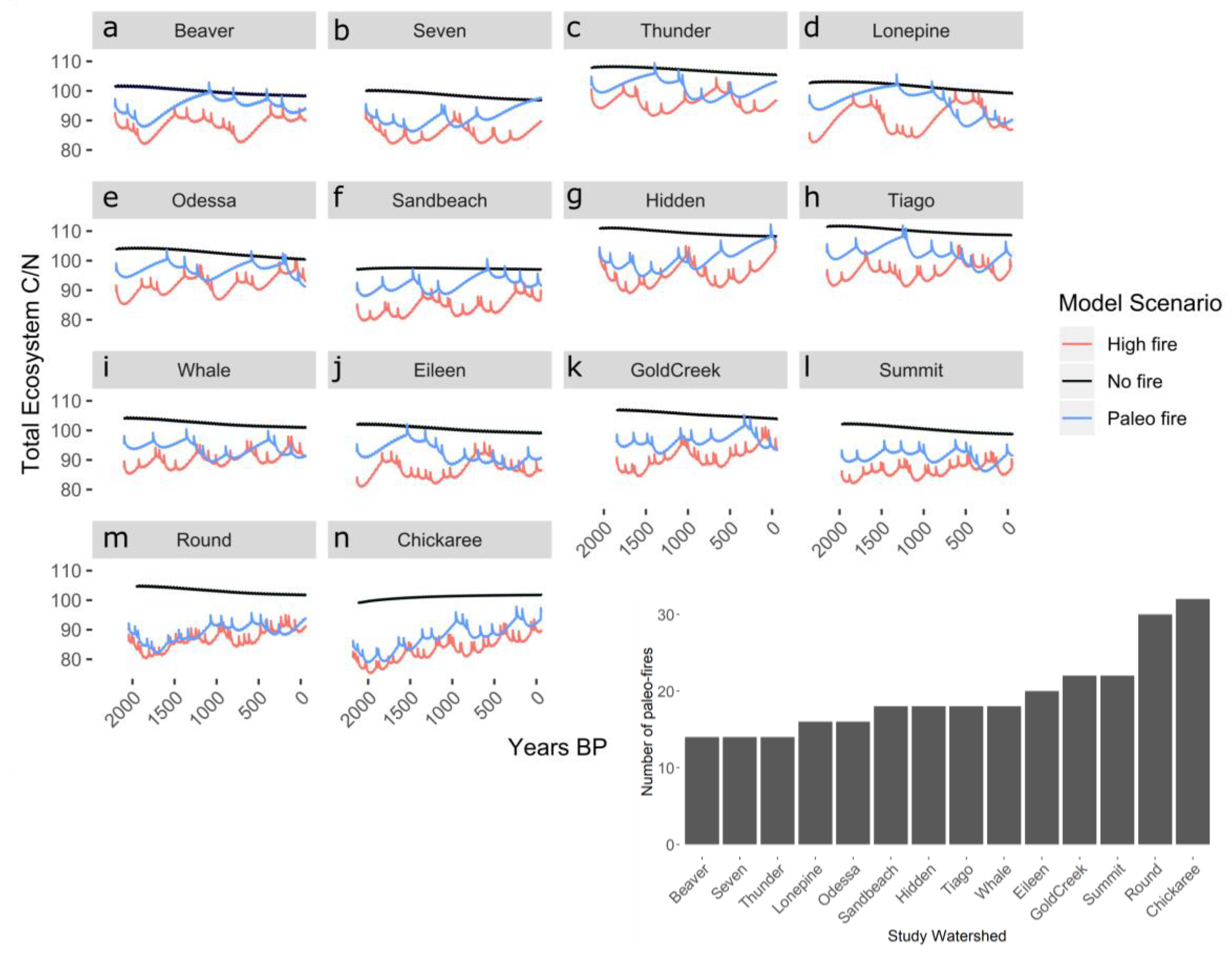
3.1. Impacts of Varying Fire Frequency on Long-Term Carbon and Nitrogen Dynamics of Subalpine Forests
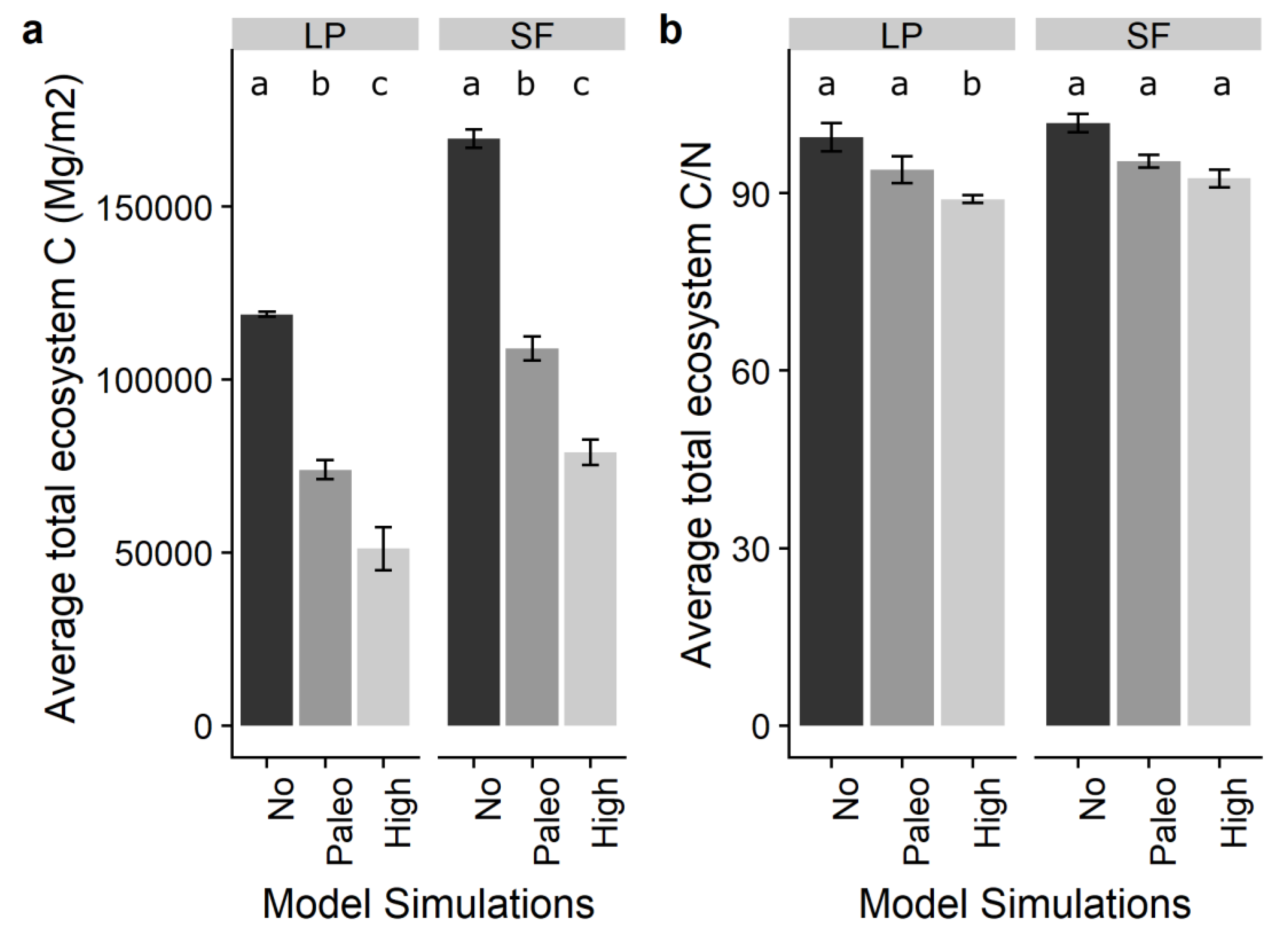
3.2. Impacts of Forest Type on Carbon and Nitrogen Dynamics
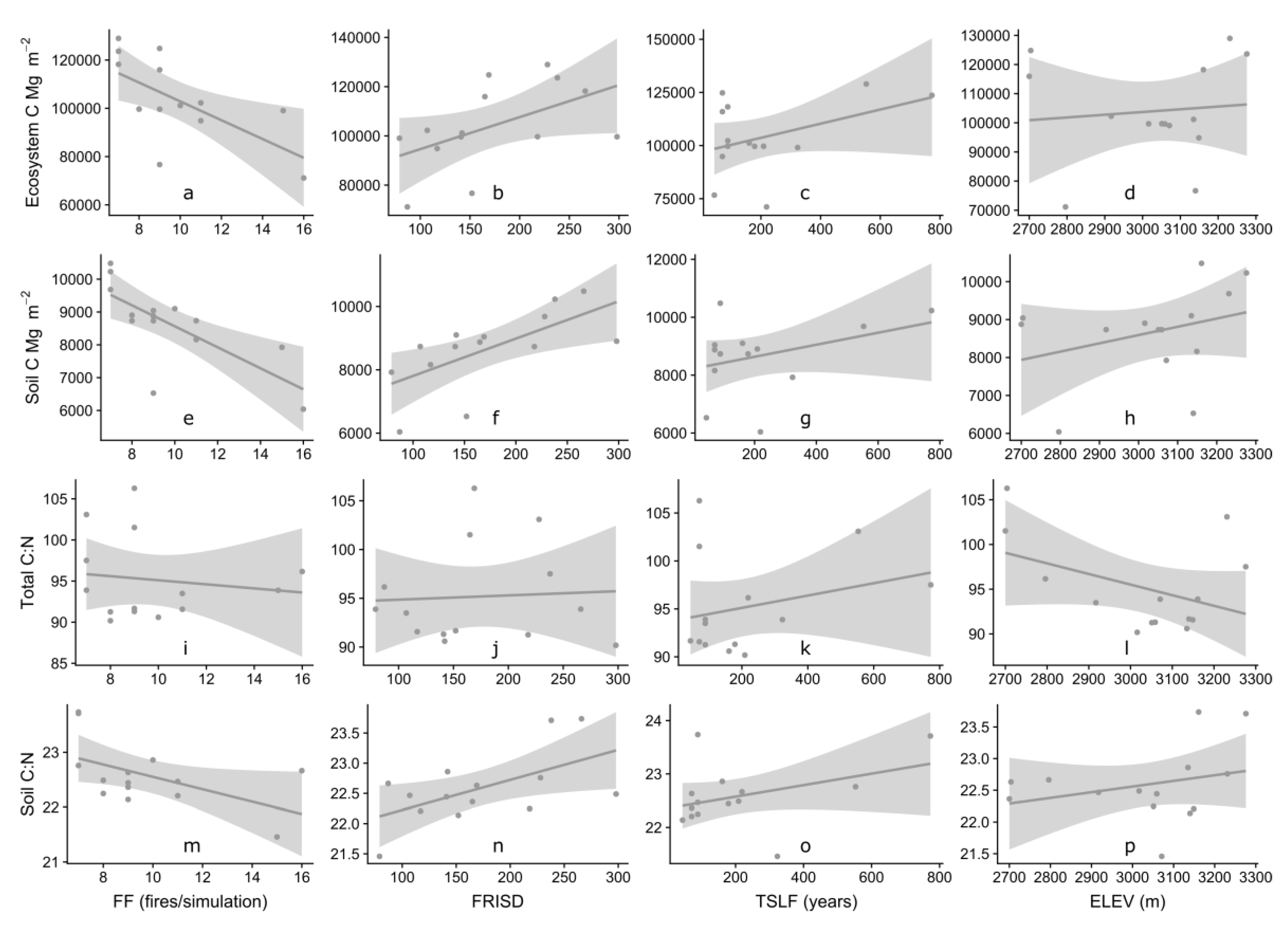
3.3. Influence of Fire Frequency and Site Characteristics on C and N Dynamics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Le Quéré, C.; Andrew, R.M.; Friedlingstein, P.; Sitch, S.; Hauck, J.; Pongratz, J.; Pickers, P.A.; Korsbakken, J.I.; Peters, G.P.; Canadell, J.G.; et al. Global Carbon Budget 2018. Earth Syst. Sci. Data 2018, 10, 2141–2194. [Google Scholar] [CrossRef] [Green Version]
- Intergovernmental Panel on Climate Change IPCC. Part A: Global and Sectoral Aspects. In Climate Change 2014: Impacts, Adaptation, and Vulnerability; Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; pp. 1–32. [Google Scholar]
- Hudiburg, T.W.; Law, B.E.; Turner, D.P.; Campbell, J.L.; Donato, D.C.; Duane, M. Carbon dynamics of Oregon and Northern California forests and potential land-based carbon storage. Ecol. Appl. 2009, 19, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Buotte, P.C.; Levis, S.; Law, B.E.; Hudiburg, T.W.; Rupp, D.E.; Kent, J.J. Near-future forest vulnerability to drought and fire varies across the western United States. Glob. Chang. Biol. 2019, 25, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwalm, C.R.; Williams, C.A.; Schaefer, K.; Baldocchi, D.; Black, T.A.; Goldstein, A.H.; Law, B.E.; Oechel, W.C.; Scott, R.L. Reduction in carbon uptake during turn of the century drought in western North America. Nat. Geosci. 2012, 5, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Hudiburg, T.W.; Higuera, P.E.; Hicke, J.A. Fire-regime variability impacts forest carbon dynamics for centuries to millennia. Biogeosciences 2017, 14, 3873–3882. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.; Genet, H.; McGuire, A.D.; Hu, F.S. Palaeodata-informed modelling of large carbon losses from recent burning of boreal forests. Nat. Clim. Chang. 2015, 6, 79–82. [Google Scholar] [CrossRef]
- Amiro, B.D.; Barr, A.G.; Barr, J.G.; Black, T.A.; Bracho, R.; Brown, M.; Chen, J.; Clark, K.L.; Davis, K.J.; Desai, A.R.; et al. Ecosystem carbon dioxide fluxes after disturbance in forests of North America. J. Geophys. Res. Biogeosci. 2010, 115. [Google Scholar] [CrossRef]
- Berner, L.T.; Law, B.E.; Meddens, A.J.; Hicke, J.A. Tree mortality from fires, bark beetles, and timber harvest during a hot and dry decade in the western United States (2003–2012). Environ. Res. Lett. 2017, 12, 065005. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Hurteau, M.D.; Westerling, A.L. Potential decline in carbon carrying capacity under projected climate-wildfire interactions in the Sierra Nevada. Sci. Rep. 2017, 7, 2420. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Spies, T.A. Disturbance legacies increase the resilience of forest ecosystem structure, composition, and functioning. Ecol. Appl. 2014, 24, 2063–2077. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.G.; Braziunas, K.H.; Hansen, W.D.; Harvey, B.J. Short-interval severe fire erodes the resilience of subalpine lodgepole pine forests. Proc. Natl. Acad. Sci. USA 2019, 116, 11319–11328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoennagel, T.; Veblen, T.T.; Romme, W.H. The Interaction of Fire, Fuels, and Climate across Rocky Mountain Forests. Bioscience 2004, 54, 661–676. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E.; Hoffman, W.; Kenny, B.; Lloret, F.; Trabaud, L. Plant functional traits in relation to fire in crown-fire ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef] [Green Version]
- Clarke, P.J.; Lawes, M.J.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulos, H.; Barton, A.; Slingsby, J.; Bowman, D. Do Mixed Fire Regimes Shape Plant Flammability and Post-Fire Recovery Strategies? Fire 2018, 1, 39. [Google Scholar] [CrossRef] [Green Version]
- Archibald, S.; Hempson, G.P.; Lehmann, C. A unified framework for plant life-history strategies shaped by fire and herbivory. New Phytol. 2019. [Google Scholar] [CrossRef]
- Leys, B.; Higuera, P.E.; McLauchlan, K.K.; Dunnette, P.V. Wildfires and geochemical change in a subalpine forest over the past six millennia. Environ. Res. Lett. 2016, 11, 125003. [Google Scholar] [CrossRef]
- Pompeani, D.P.; McLauchlan, K.K.; Chileen, B.V.; Wolf, K.D.; Higuera, P.E. Variation of key elements in soils and plant tissues in subalpine forests of the northern Rocky Mountains, USA. Biogeosci. Discuss. 2018, 1–19. [Google Scholar] [CrossRef]
- Davis, K.T.; Dobrowski, S.Z.; Higuera, P.E.; Holden, Z.A.; Veblen, T.T.; Rother, M.T.; Parks, S.A.; Sala, A.; Maneta, M.P. Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 6193–6198. [Google Scholar] [CrossRef] [Green Version]
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2018, 21, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balch, J.K.; Bradley, B.A.; Abatzoglou, J.T.; Nagy, R.C.; Fusco, E.J.; Mahood, A.L. Human-started wildfires expand the fire niche across the United States. Proc. Natl. Acad. Sci. USA 2017, 114, 2946–2951. [Google Scholar] [CrossRef] [Green Version]
- Littell, J.S.; Mckenzie, D.; Peterson, D.L.; Anthony, L. Climate and Wildfire Area Burned in Western U.S. Ecoprovinces, 1916–2003. Ecol. Appl. 2009, 19, 1003–1021. [Google Scholar] [CrossRef] [PubMed]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littell, J.S.; Peterson, D.L.; Riley, K.L.; Liu, Y.; Luce, C.H. A review of the relationships between drought and forest fire in the United States. Glob. Chang. Biol. 2016, 22, 2353–2369. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Safford, H.D.; Crimmins, M.; Thode, A.E. Quantitative evidence for increasing forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA. Ecosystems 2009, 12, 16–32. [Google Scholar] [CrossRef]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2016, 53, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S., III; Woodwell, G.M.; Randerson, J.T.; Rastetter, E.B.; Lovett, G.M.; Baldocchi, D.D.; Clark, D.A.; Harmon, M.E.; Schimel, D.S.; Valentini, R.; et al. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems 2006, 9, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G.B.; Doney, S.C. Climate, ecosystems, and planetary futures: The challenge to predict life in Earth system models. Science 2018, 359, eaam8328. [Google Scholar] [CrossRef] [Green Version]
- Law, B.E.; Hudiburg, T.W.; Berner, L.T.; Kent, J.J.; Buotte, P.C.; Harmon, M.E. Land use strategies to mitigate climate change in carbon dense temperate forests. Proc. Natl. Acad. Sci. USA 2018, 115, 3663–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, R.A.; Koven, C.D.; Anderegg, W.R.L.; Christoffersen, B.O.; Dietze, M.C.; Farrior, C.E.; Holm, J.A.; Hurtt, G.C.; Knox, R.G.; Lawrence, P.J.; et al. Vegetation demographics in Earth System Models: A review of progress and priorities. Glob. Chang. Biol. 2018, 24, 35–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hantson, S.; Arneth, A.; Harrison, S.P.; Kelley, D.I.; Prentice, I.C.; Rabin, S.S.; Archibald, S.; Mouillot, F.; Arnold, S.R.; Artaxo, P.; et al. The status and challenge of global fire modelling. Biogeosciences 2016, 13, 3359–3375. [Google Scholar] [CrossRef] [Green Version]
- Calder, W.J.; Parker, D.; Stopka, C.J.; Jiménez-Moreno, G.; Shuman, B.N. Medieval warming initiated exceptionally large wildfire outbreaks in the Rocky Mountains. Proc. Natl. Acad. Sci. USA 2015, 112, 13261–13266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunnette, P.V.; Higuera, P.E.; Mclauchlan, K.K.; Derr, K.M.; Briles, C.E.; Keefe, M.H. Biogeochemical impacts of wildfires over four millennia in a Rocky Mountain subalpine watershed. New Phytol. 2014, 203, 900–912. [Google Scholar] [CrossRef]
- Higuera, P.E.; Briles, C.E.; Whitlock, C. Fire-regime complacency and sensitivity to centennial through millennial-scale climate change in Rocky Mountain subalpine forests, Colorado, USA. J. Ecol. 2014, 102, 1429–1441. [Google Scholar] [CrossRef]
- Bai, E.; Houlton, B.Z. Coupled isotopic and process-based modeling of gaseous nitrogen losses from tropical rain forests. Glob. Biogeochem. Cycles 2009, 23, 1–10. [Google Scholar] [CrossRef]
- Hartman, M.D.; Baron, J.S.; Ojima, D.S. Application of a coupled ecosystem-chemical equilibrium model, DayCent-Chem, to stream and soil chemistry in a Rocky Mountain watershed. Ecol. Modell. 2007, 200, 493–510. [Google Scholar] [CrossRef]
- Savage, K.E.; Parton, W.J.; Davidson, E.A.; Trumbore, S.E.; Frey, S.D. Long-term changes in forest carbon under temperature and nitrogen amendments in a temperate northern hardwood forest. Glob. Chang. Biol. 2013, 19, 2389–2400. [Google Scholar] [CrossRef] [Green Version]
- Hengl, T.; De Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [Green Version]
- Stenzel, J.E.; Bartowitz, K.J.; Hartman, M.D.; Lutz, J.A.; Crystal, A.; Parton, W.J.; Hudiburg, T.W. Fixing a snag in carbon emissions estimates from wildfires. Glob. Chang. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Law, B.E.; Arkebauer, T.; Chen, J.; Campbell, J.L.; Sun, O.; Schwartz, M.; van Ingen, C.; Verma, S. Terrestrial Carbon Observations: Protocols for Vegetation Sampling and Data Submission; Global Terrestrial Observing System, FAO: Rome, Italy, 2008. [Google Scholar]
- Sampson, D.A.; Allen, H.L. Direct and indirect estimates of Leaf Area Index (LAI) for lodgepole and loblolly pine stands. Trees 1995, 9, 119–122. [Google Scholar] [CrossRef]
- Aplet, G.H.; Smith, F.W.; Laven, R.D. Stemwood Biomass and Production during Spruce-Fir Stand Development. J. Ecol. 1989, 77, 70–77. [Google Scholar] [CrossRef]
- Thornton, P.E.; Thornton, M.M.; Mayer, B.W.; Wilhelmi, N.; Wei, Y.; Devarakonda, R.; Cook, R. Daymet: Daily surface weather on a 1 km grid for North America, 1980–2008. Oak Ridge Natl. Lab. Distrib. Act. Arch. Cent. Biogeochem. Dyn. DAAC 2012. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Brown, C.D.; Johnstone, J.F. How does increased fire frequency affect carbon loss from fire? A case study in the northern boreal forest. Int. J. Wildl. Fire 2011, 20, 829–837. [Google Scholar] [CrossRef]
- Law, B.E.; Waring, R.H. Carbon implications of current and future effects of drought, fire and management on Pacific Northwest forests. For. Ecol. Manag. 2015, 355, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Knicker, H. How does fire affect the nature and stability of soil organic nitrogen and carbon? A review. Biogeochemistry 2007, 85, 91–118. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, Y.; Finzi, A.C. Carbon and nitrogen dynamics during forest stand development: A global synthesis. New Phytol. 2011, 190, 977–989. [Google Scholar] [CrossRef]
- Caldwell, T.G.; Johnson, D.W.; Miller, W.W.; Qualls, R.G. Forest floor carbon and nitrogen losses due to prescription fire. Soil Sci. Soc. Am. J. 2002, 66, 262–267. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juárez-Orozco, S.M.; Siebe, C.; Fernández y Fernández, D. Causes and Effects of Forest Fires in Tropical Rainforests: A Bibliometric Approach. Trop. Conserv. Sci. 2017, 10, 194008291773720. [Google Scholar] [CrossRef]
- Lasslop, G.; Thonicke, K.; Kloster, S. SPITFIRE within the MPI Earth system model: Model development and evaluation. J. Adv. Model. Earth Syst. 2014, 6, 740–755. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Lat., Long. | Sub-Region | Forest Type | Elevation (m) | Mean FRI (yr) [SD] | Simulation Length |
|---|---|---|---|---|---|---|
| Eileen | 40.902, −106.674 | MBR-NF | Spruce-fir | 3135 | 220 [142] | 2197 |
| Seven | 40.896, −106.682 | MBR-NF | Upper-treeline spruce-fir | 3276 | 298 [238] | 2089 |
| Gold Creek | 40.782, −106.678 | MBR-NF | Spruce-fir | 2917 | 174 [107] | 1909 |
| Hidden | 40.771, −106.827 | MBR-NF | Spruce-fir | 2704 | 234 [169] | 2107 |
| Beaver | 40.753, −106.687 | MBR-NF | Spruce-fir | 3161 | 283 [266] | 1981 |
| Tiago | 40.579, −106.613 | MBR-NF | Spruce-fir | 2700 | 244 [165] | 2197 |
| Whale | 40.556, −106.675 | MBR-NF | Spruce-fir | 3059 | 240 [141] | 2161 |
| Summit | 40.545, −106.682 | MBR-NF | Upper-treeline spruce-fir | 3149 | 185 [117] | 2035 |
| Round | 40.473, −106.663 | MBR-NF | Spruce-fir | 3071 | 134 [79] | 2107 |
| Chickaree | 40.334, −105.840 | RMNP | Lodgepole | 2796 | 136 [87] | 2180 |
| Odessa | 40.330, −105.685 | RMNP | Spruce-fir | 3051 | 281 [218] | 2251 |
| Lonepine | 40.232, −105.730 | RMNP | Spruce-fir | 3016 | 302 [298] | 2416 |
| Thunder | 40.221, −105.647 | RMNP | Spruce-fir | 3231 | 315 [228] | 2206 |
| Sandbeach | 40.218, −105.601 | RMNP | Lodgepole | 3140 | 243 [152] | 2191 |
| Scenario | Description | Climate |
|---|---|---|
| Equilibrium | Fire prescribed using the mean fire return interval (FRI) of the paleo-fire record | Modern-recycled |
| Paleo-fire | Fire prescribed using site-specific paleo-fire record | Modern-recycled |
| High-fire | Fire prescribed by doubling the site-specific paleo-fire record; e.g., fire-history is repeated twice in the 2000-year record | Modern-recycled |
| No-fire | No disturbance/fire | Modern-recycled |
| FF | FRISD | TSLF | ELEV | ||
|---|---|---|---|---|---|
| TEC | r2 | 0.4043 | 0.2671 | 0.1696 | 0.0101 |
| Soil C | r2 | 0.5293 | 0.4259 | 0.0128 | 0.176 |
| Total C:N | r2 | 0.0182 | 0.0033 | 0.0704 | 0.1822 |
| Soil C:N | r2 | 0.2835 | 0.3278 | 0.1489 | 0.0779 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartowitz, K.J.; Higuera, P.E.; Shuman, B.N.; McLauchlan, K.K.; Hudiburg, T.W. Post-Fire Carbon Dynamics in Subalpine Forests of the Rocky Mountains. Fire 2019, 2, 58. https://doi.org/10.3390/fire2040058
Bartowitz KJ, Higuera PE, Shuman BN, McLauchlan KK, Hudiburg TW. Post-Fire Carbon Dynamics in Subalpine Forests of the Rocky Mountains. Fire. 2019; 2(4):58. https://doi.org/10.3390/fire2040058
Chicago/Turabian StyleBartowitz, Kristina J., Philip E. Higuera, Bryan N. Shuman, Kendra K. McLauchlan, and Tara W. Hudiburg. 2019. "Post-Fire Carbon Dynamics in Subalpine Forests of the Rocky Mountains" Fire 2, no. 4: 58. https://doi.org/10.3390/fire2040058
APA StyleBartowitz, K. J., Higuera, P. E., Shuman, B. N., McLauchlan, K. K., & Hudiburg, T. W. (2019). Post-Fire Carbon Dynamics in Subalpine Forests of the Rocky Mountains. Fire, 2(4), 58. https://doi.org/10.3390/fire2040058