
Low-Severity Wildfire Shifts Mixed Conifer Forests toward Historical Stand Structure in Guadalupe Mountains National Park, Texas, USA

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
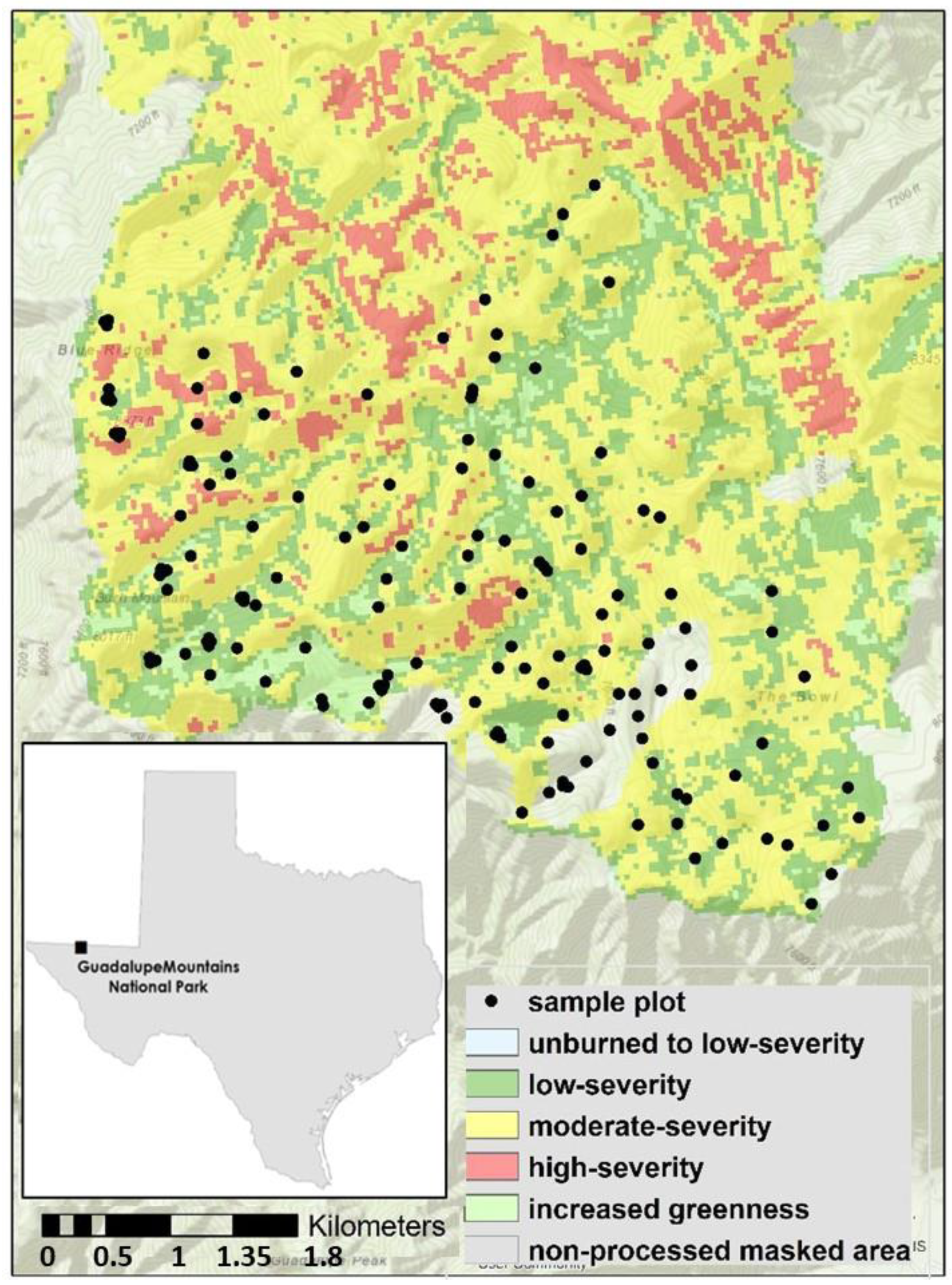
2.1. Study Area
2.2. The Coyote Fire
2.3. Stand Structure and Fuels Plots
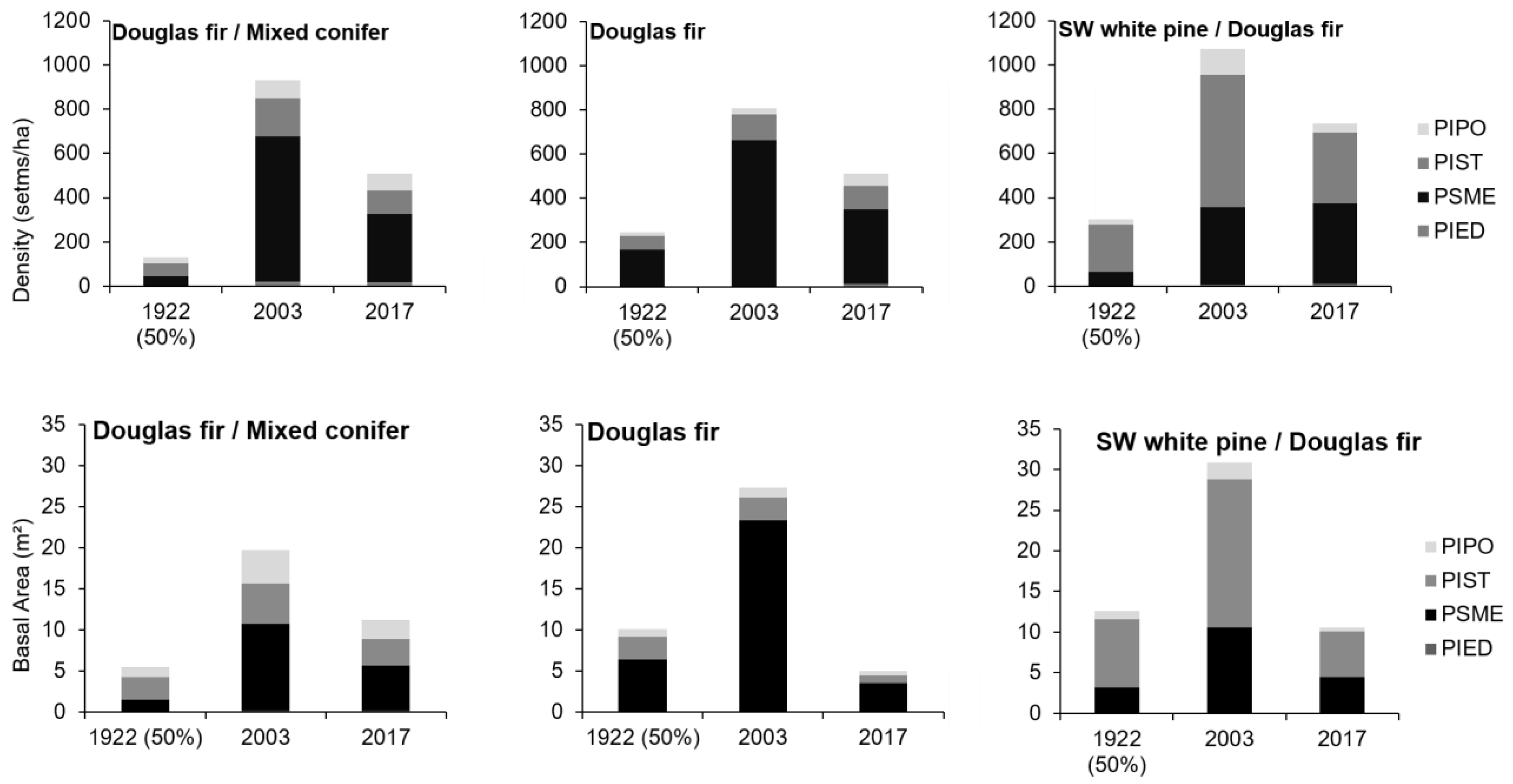
2.4. Stand Structure Reconstruction
2.5. Changes in Stand Composition
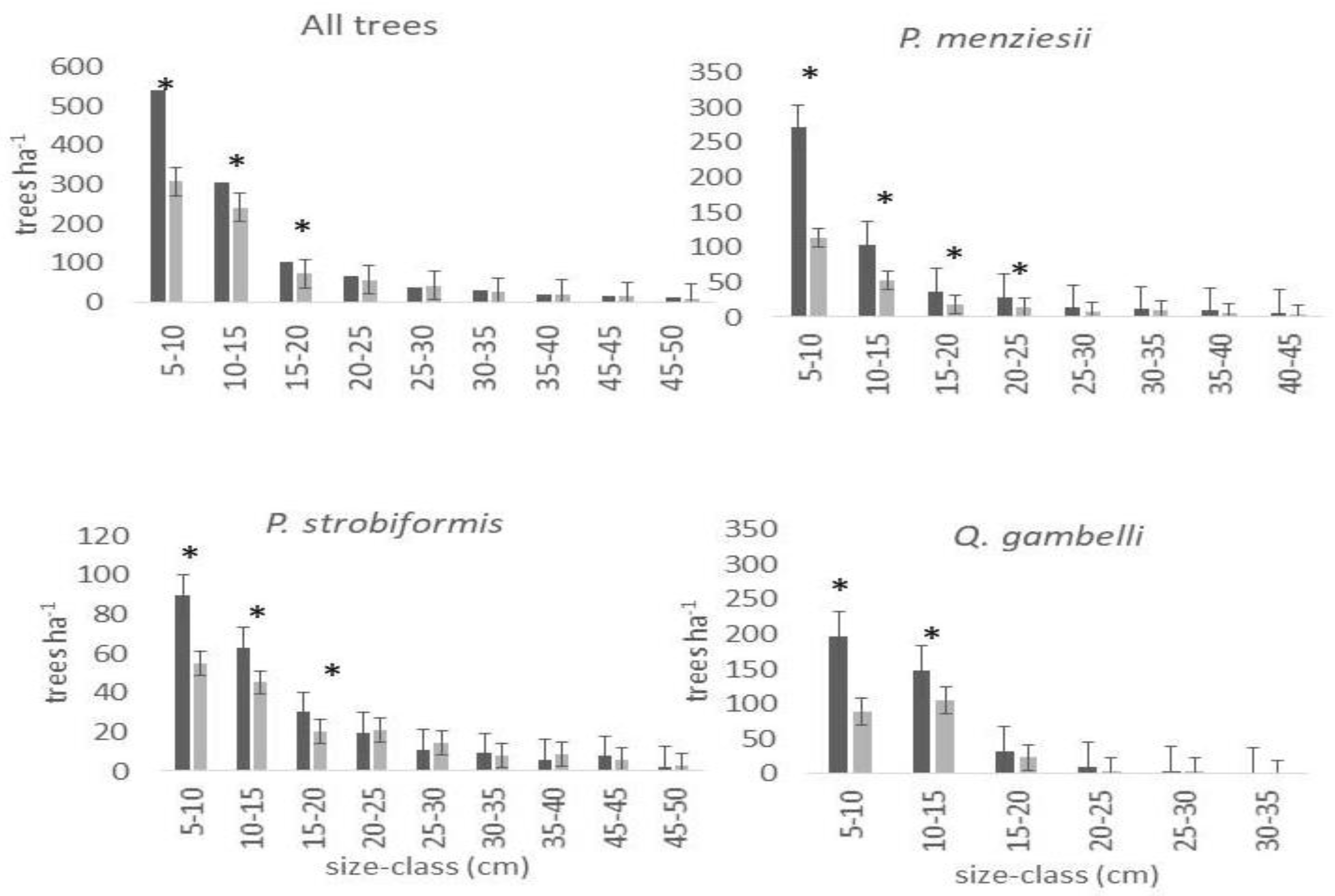
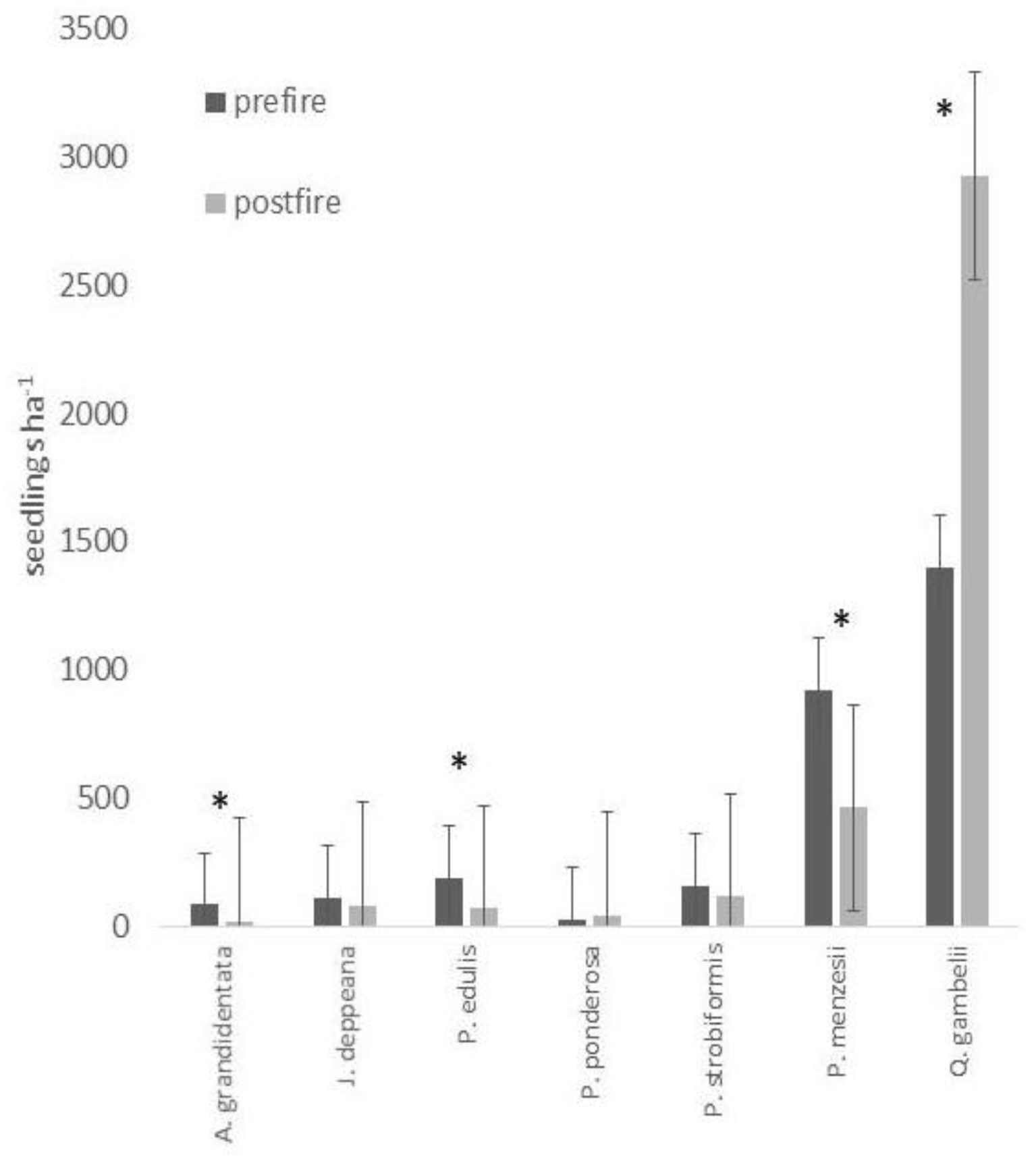
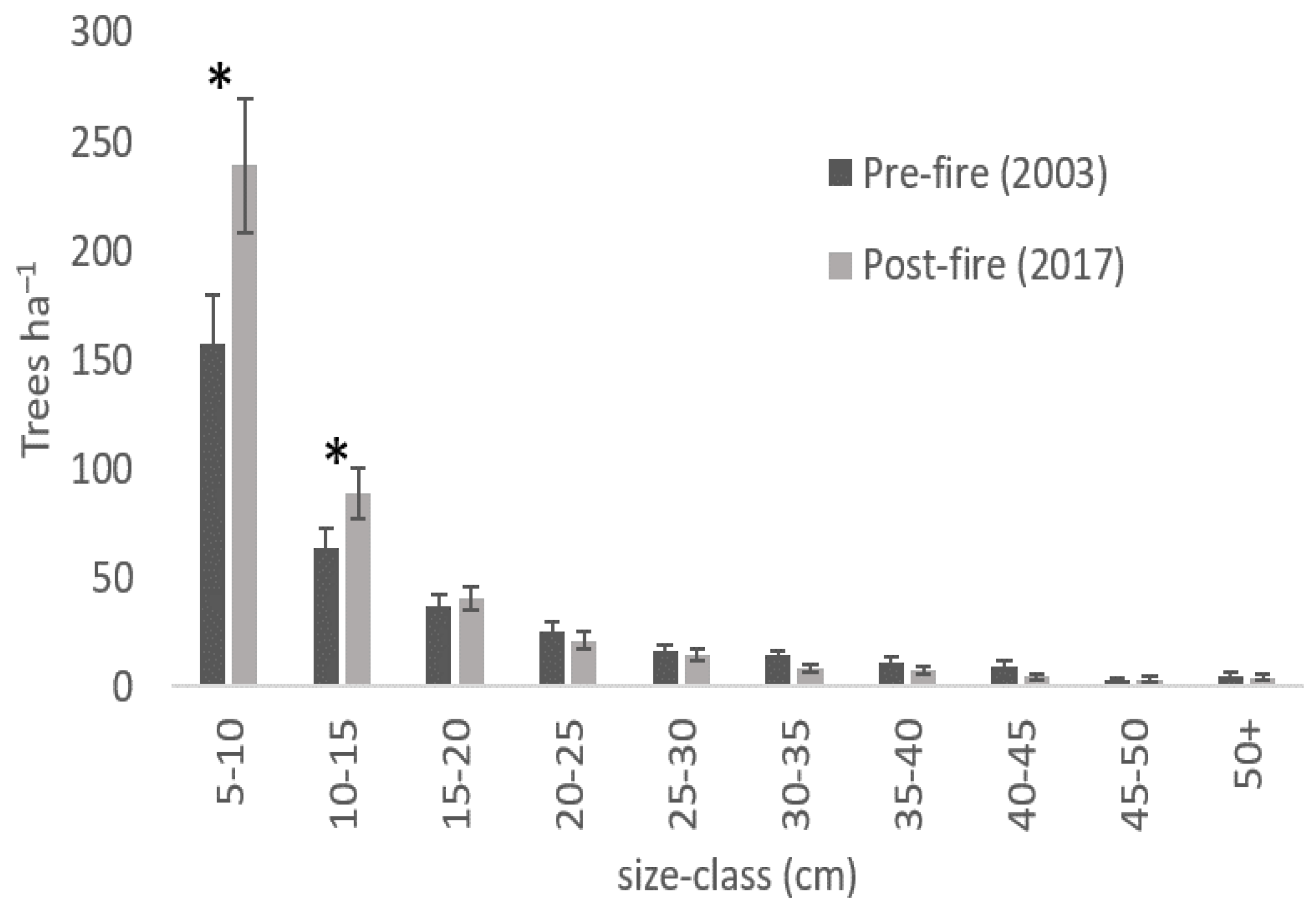
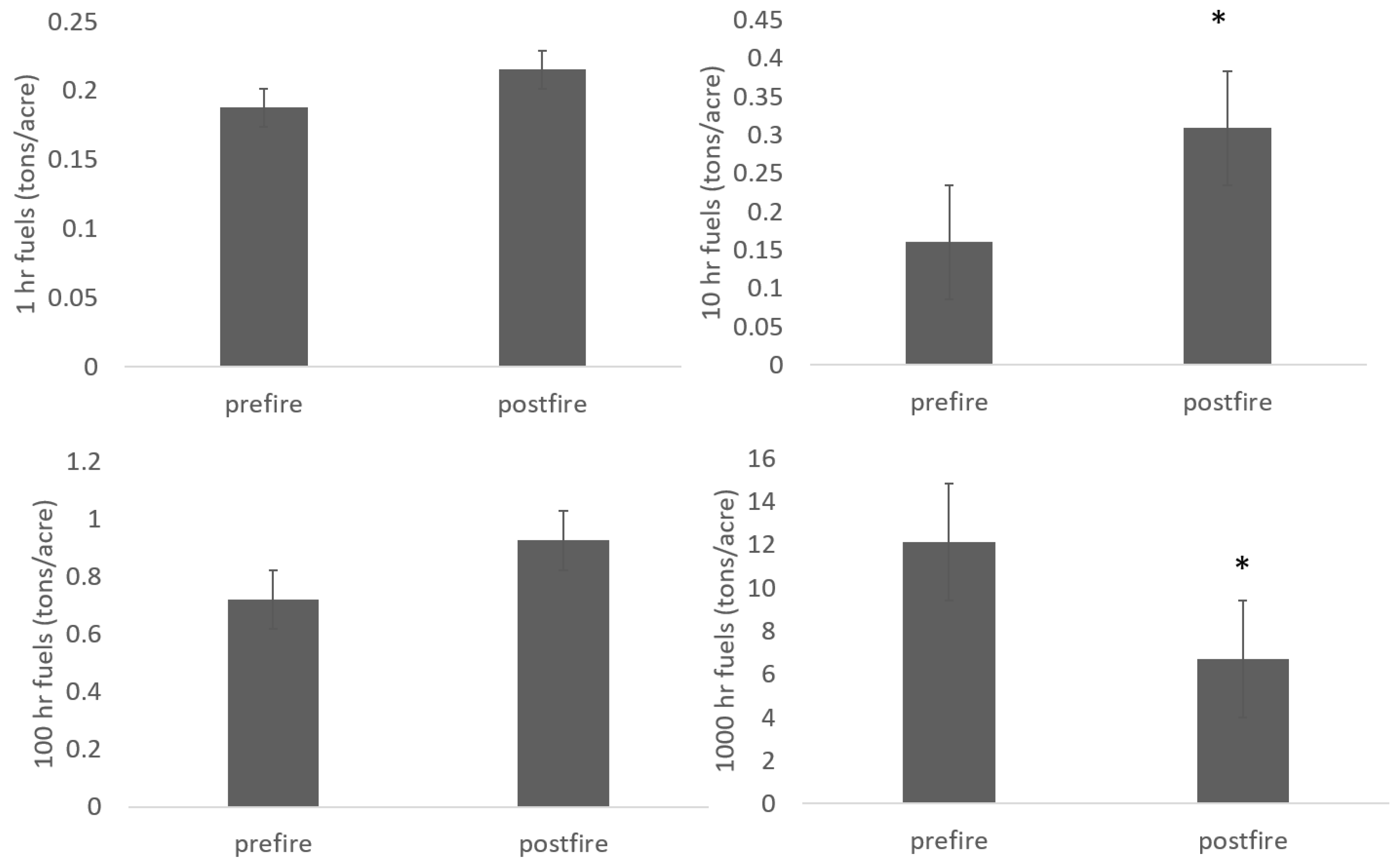
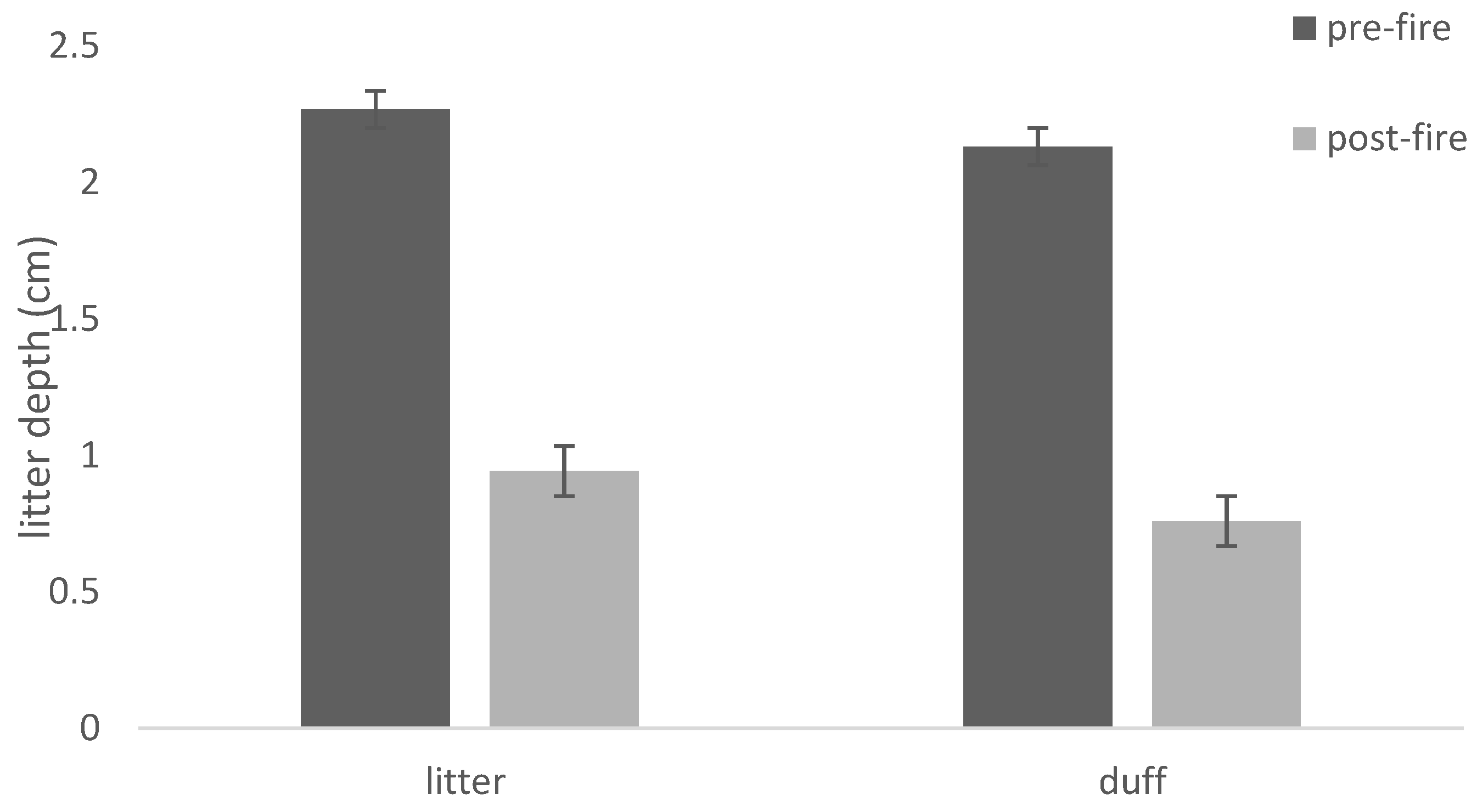
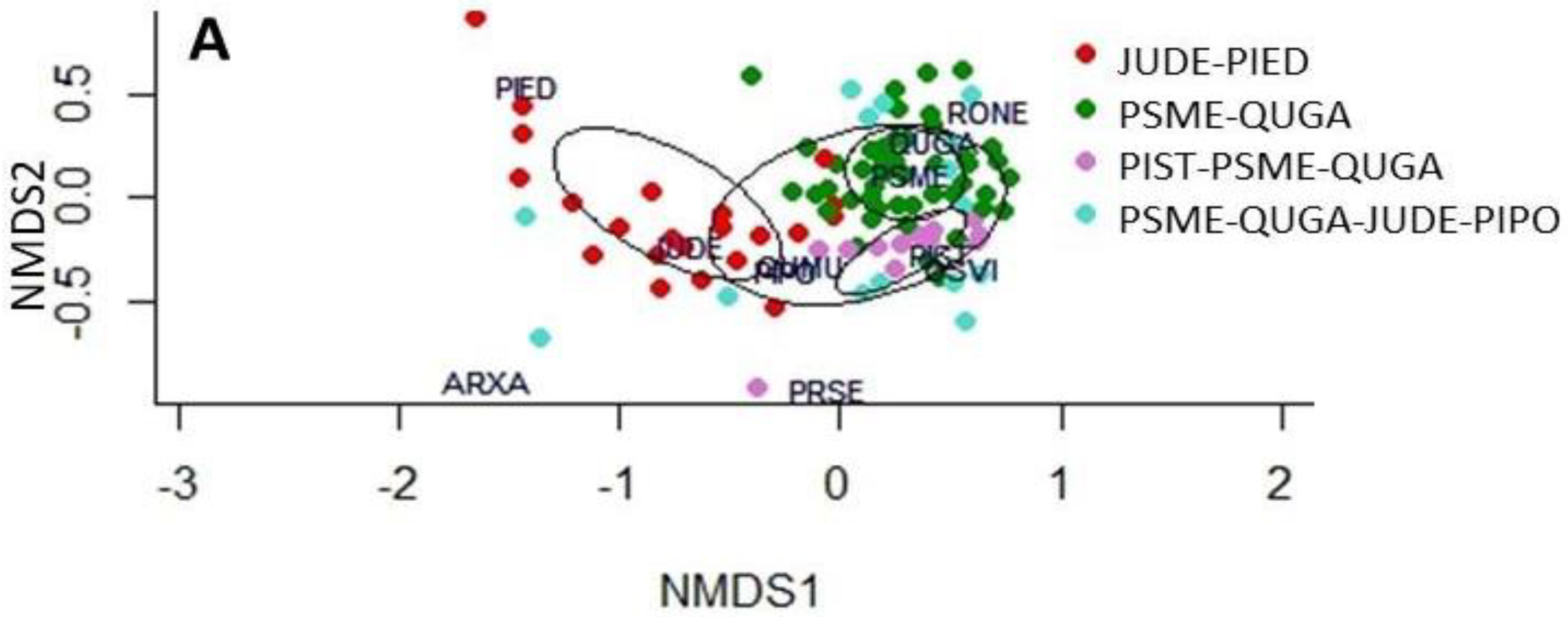
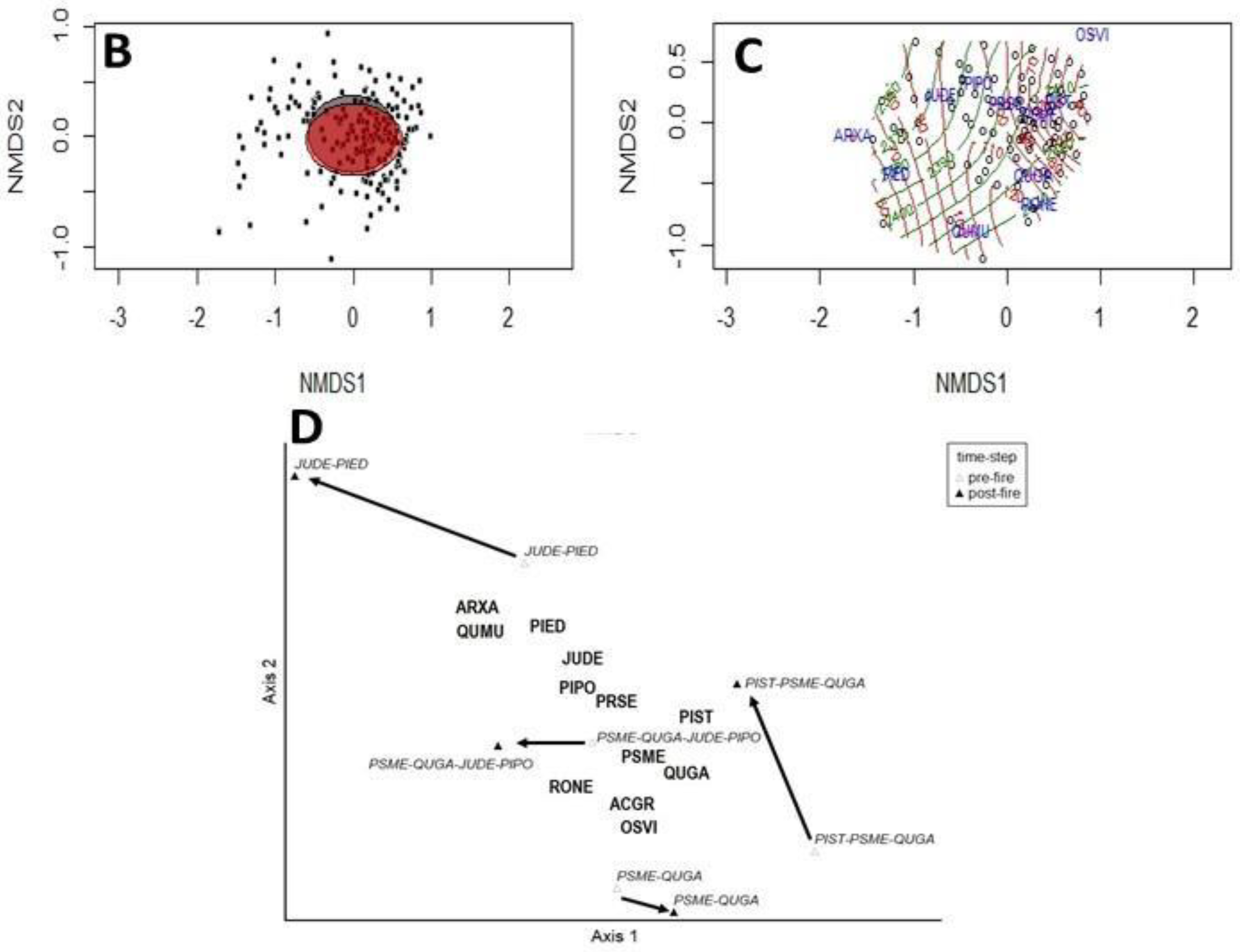
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Westerling, A.L. Increasing western US forest wildfire activity: Sensitivity to changes in the timing of spring. Philos. Trans. R. Soc. B 2016, 371, 20150178. [Google Scholar] [CrossRef] [PubMed]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Hurteau, M.D.; Bradford, J.B.; Fulé, P.Z.; Taylor, A.H.; Martin, K.L. Climate change, fire management, and ecological services in the southwestern US. For. Ecol. Manag. 2014, 327, 280–289. [Google Scholar] [CrossRef]
- Nagy, R.; Fusco, E.; Bradley, B.; Abatzoglou, J.T.; Balch, J. Human-related ignitions increase the number of large wildfires across US ecoregions. Fire 2018, 1, 4. [Google Scholar] [CrossRef]
- Covington, W.W.; Moore, M.M. Southwestern Ponderosa Forest Structure—Changes since Euro-American Settlement. J. For. 1994, 92, 39–47. [Google Scholar]
- Poulos, H.M.; Villanueva Díaz, J.; Cerano Paredes, J.; Camp, A.E.; Gatewood, R.G. Human influences on fire regimes and forest structure in the Chihuahuan Desert Borderlands. For. Ecol. Manag. 2013, 298, 1–11. [Google Scholar] [CrossRef]
- Poulos, H.; Gatewood, R.; Camp, A. Fire regimes of the piñon-juniper woodlands of Big Bend National Park and the Davis Mountains, west Texas, USA. Can. J. For. Res. 2009, 39, 1236–1246. [Google Scholar] [CrossRef]
- Sakulich, J.; Taylor, A.H. Fire regimes and forest structure in a sky island mixed conifer forest, Guadalupe Mountains National Park, Texas, USA. For. Ecol. Manag. 2007, 241, 62–73. [Google Scholar] [CrossRef]
- Savage, M.; Swetnam, T.W.J.E. Early 19th-century fire decline following sheep pasturing in a Navajo ponderosa pine forest. Ecology 1990, 71, 2374–2378. [Google Scholar] [CrossRef]
- Coop, J.D.; Parks, S.A.; McClernan, S.R.; Holsinger, L.M. Influences of prior wildfires on vegetation response to subsequent fire in a reburned southwestern landscape. Ecol. Appl. 2016, 26, 346–354. [Google Scholar] [CrossRef]
- Covington, W.W.; Fule, P.Z.; Moore, M.M.; Hart, S.C.; Kolb, T.E.; Mast, J.N.; Sackett, S.S.; Wagner, M.R. Restoring ecosystem health in ponderosa pine forests of the Southwest. J. For. 1997, 95, 23. [Google Scholar]
- O’Connor, C.D.; Falk, D.A.; Lynch, A.M.; Swetnam, T.W. Fire severity, size, and climate associations diverge from historical precedent along an ecological gradient in the Pinaleño Mountains, Arizona, USA. For. Ecol. Manag. 2014, 329, 264–278. [Google Scholar] [CrossRef]
- Barton, A.M.; Poulos, H.M. Pine vs. oaks revisited: Conversion of Madrean pine-oak forest to oak shrubland after high-severity wildfire in the Sky Islands of Arizona. For. Ecol. Manag. 2018, 414, 28–40. [Google Scholar] [CrossRef]
- Stephens, S.L.; Collins, B.M.; Fettig, C.J.; Finney, M.A.; Hoffman, C.M.; Knapp, E.E.; North, M.P.; Safford, H.; Wayman, R.B. Drought, Tree Mortality, and Wildfire in Forests Adapted to Frequent Fire. BioScience 2018, 68, 77–88. [Google Scholar] [CrossRef]
- Moritz, M.A.; Batllori, E.; Bradstock, R.A.; Gill, A.M.; Handmer, J.; Hessburg, P.F.; Leonard, J.; McCaffrey, S.; Odion, D.C.; Schoennagel, T. Learning to coexist with wildfire. Nature 2014, 515, 58–66. [Google Scholar] [CrossRef]
- Schoennagel, T.; Balch, J.K.; Brenkert-Smith, H.; Dennison, P.E.; Harvey, B.J.; Krawchuk, M.A.; Mietkiewicz, N.; Morgan, P.; Moritz, M.A.; Rasker, R.; et al. Adapt to more wildfire in western North American forests as climate changes. Proc. Natl. Acad. Sci. USA 2017, 114, 4582–4590. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Conifer regeneration in stand-replacement portions of a large mixed-severity wildfire in the Klamath–Siskiyou Mountains. Can. J. For. Res. 2009, 39, 823–838. [Google Scholar] [CrossRef]
- Haffey, C.; Sisk, T.D.; Allen, C.D.; Thode, A.E.; Margolis, E.Q. Limits to Ponderosa Pine Regeneration following Large High-Severity Forest Fires in the United States Southwest. Fire Ecol. 2018, 14, 143–163. [Google Scholar] [CrossRef]
- Halofsky, J.; Donato, D.; Hibbs, D.; Campbell, J.; Cannon, M.D.; Fontaine, J.; Thompson, J.R.; Anthony, R.; Bormann, B.; Kayes, L. Mixed-severity fire regimes: Lessons and hypotheses from the Klamath-Siskiyou Ecoregion. Ecosphere 2011, 2, 1–19. [Google Scholar] [CrossRef]
- Keyser, T.L.; Lentile, L.B.; Smith, F.W.; Shepperd, W.D. Changes in forest structure after a large, mixed-severity wildfire in ponderosa pine forests of the Black Hills, South Dakota, USA. For. Sci. 2008, 54, 328–338. [Google Scholar]
- Lydersen, J.M.; Collins, B.M.; Hunsaker, C.T. Implementation constraints limit benefits of restoration treatments in mixed-conifer forests. Int. J. Wildland Fire 2019, 28, 495–511. [Google Scholar] [CrossRef]
- Owen, S.M.; Sieg, C.H.; Meador, A.J.S.; Fulé, P.Z.; Iniguez, J.M.; Baggett, L.S.; Fornwalt, P.J.; Battaglia, M.A. Spatial patterns of ponderosa pine regeneration in high-severity burn patches. For. Ecol. Manag. 2017, 405, 134–149. [Google Scholar] [CrossRef]
- Parks, S.A.; Dobrowski, S.Z.; Panunto, M.H. What drives low-severity fire in the Southwestern USA? Forests 2018, 9, 165. [Google Scholar] [CrossRef]
- Poulos, H.M.; Reemts, C.M.; Wogan, K.A.; Karges, J.P.; Gatewood, R.G. Multiple wildfires with minimal consequences: Low-severity wildfire effects on West Texas piñon-juniper woodlands. For. Ecol. Manag. 2020, 473, 118293. [Google Scholar] [CrossRef]
- Romme, W.H.; Allen, C.D.; Bailey, J.D.; Baker, W.L.; Bestelmeyer, B.T.; Brown, P.M.; Eisenhart, K.S.; Floyd, M.L.; Huffman, D.W.; Jacobs, B.F. Historical and modern disturbance regimes, stand structures, and landscape dynamics in piñon–juniper vegetation of the western United States. Rangel. Ecol. Manag. 2009, 62, 203–222. [Google Scholar] [CrossRef]
- Strom, B.A.; Fulé, P.Z. Pre-wildfire fuel treatments affect long-term ponderosa pine forest dynamics. Int. J. Wildland Fire 2007, 16, 128–138. [Google Scholar] [CrossRef]
- Coppoletta, M.; Merriam, K.E.; Collins, B.M. Post-fire vegetation and fuel development influences fire severity patterns in reburns. Ecol. Appl. 2016, 26, 686–699. [Google Scholar] [CrossRef]
- Harris, L.; Taylor, A.H. Topography, fuels, and fire exclusion drive fire severity of the Rim Fire in an old-growth mixed-conifer forest, Yosemite National Park, USA. Ecosystems 2015, 18, 1192–1208. [Google Scholar] [CrossRef]
- Ahlstrand, G.M. Fire History of a Mixed Conifer Forest in Guadalupe Mountains National Park. In Proceedings of the Fire History Workshop, Tucson, Arizona, 20–24 October 1980; p. 4. [Google Scholar]
- Kittams, W.H. Effect of fire on vegetation of the Chihuahuan Desert region. In Proceedings of the 13th Tall Timbers Fire Ecology Conference, Tallahassee, FL, USA, 1 December 1973. [Google Scholar]
- Fabry, J.K. Guadalupe Mountains National Park: An Administrative History; Division of History, Southwest Cultural Resources Center: Santa Fe, NM, USA, 1990. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Thomas, J.W. Wildlife Habitats in Managed Forests: The Blue Mountains of Oregon and Washington; Wildlife Management Institute: Washington, DC, USA, 1979. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- Rogers, J.J. ECOSIM: A System for Projecting Multiresource Outputs under Alternative Forest Management Regimes Administrative Report. 1984. Available online: https://agris.fao.org/agris-search/search.do?recordID=US8919234 (accessed on 8 July 2021).
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A.; Charrad, M.M. Package ‘NbClust’. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Packag. Version 2013, 2, 1–295. [Google Scholar]
- Stoddard, M.T.; Sánchez Meador, A.J.; Fulé, P.Z.; Korb, J.E. Five-year post-restoration conditions and simulated climate-change trajectories in a warm/dry mixed-conifer forest, southwestern Colorado, USA. For. Ecol. Manag. 2015, 356, 253–261. [Google Scholar] [CrossRef]
- Higgins, A.M.; Waring, K.M.; Thode, A.E. The effects of burn entry and burn severity on ponderosa pine and mixed conifer forests in Grand Canyon National Park. Int. J. Wildland Fire 2015, 24, 495–506. [Google Scholar] [CrossRef]
- Erdman, J.A. Pinyon-juniper succession after natural fires on residual soils of Mesa Verde, Colorado. Brigh. Young Univ. Sci. Bull. Biol. Ser. 1970, 11, 1. [Google Scholar]
- Kaufmann, M.R.; Huisjen, D.W.; Kitchen, S.; Babler, M.; Abella, S.R.; Gardiner, T.S.; McAvoy, D.; Howie, J.; Douglas, H.P. Gambel Oak Ecology and Management in the Southern Rockies: The Status of Our Knowledge; Colorado State University, Southern Rockies Fire Sciences Network: Fort Collins, CO, USA, 2016; 16p. [Google Scholar]
- Poulos, H.; Gatewood, R. Effectiveness of Thinning and Prescribed Fire Fuel Treatments in Piñon-Juniper Woodlands of the Davis Mountains, West Texas, USA. J. Sustain. For. 2013, 32, 806–821. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Anderson, R.S.; Jass, R.B.; Toney, J.L.; Allen, C.D.; Cisneros-Dozal, L.M.; Hess, M.; Heikoop, J.; Fessenden, J. Development of the mixed conifer forest in northern New Mexico and its relationship to Holocene environmental change. Quat. Res. 2008, 69, 263–275. [Google Scholar] [CrossRef]
- Malone, S.L.; Fornwalt, P.J.; Battaglia, M.A.; Chambers, M.E.; Iniguez, J.M.; Sieg, C.H. Mixed-severity fire fosters heterogeneous spatial patterns of conifer regeneration in a dry conifer forest. Forests 2018, 9, 45. [Google Scholar] [CrossRef]
- Korb, J.E.; Fornwalt, P.J.; Stevens-Rumann, C.S. What drives ponderosa pine regeneration following wildfire in the western United States? For. Ecol. Manag. 2019, 454, 117663. [Google Scholar] [CrossRef]
- Parsons, D.J.; Keane, R.E. Mixed-severity fire regimes in the northern Rocky Mountains: Consequences of fire exclusion and options for the future. In Wilderness Science in a Time of Change Conference: Wilderness Ecosystems, Threats, and Management; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; Volume 5. [Google Scholar]
- Perry, D.A.; Hessburg, P.F.; Skinner, C.N.; Spies, T.A.; Stephens, S.L.; Taylor, A.H.; Franklin, J.F.; McComb, B.; Riegel, G. The ecology of mixed severity fire regimes in Washington, Oregon, and Northern California. For. Ecol. Manag. 2011, 262, 703–717. [Google Scholar] [CrossRef]
- Fulé, P. Effects of an intense wildfire in a Mexican oak-pine forest. For. Sci. 2000, 46, 52–61. [Google Scholar]
- Barton, A.M.; Poulos, H. Wildfire and topography drive woody plant diversity in a Sky Island mountain range in the Southwest USA. Ecol. Evol. 2021, 11, 14715–14732. [Google Scholar] [CrossRef]
- Poulos, H.M.; Freiburger, M.R.; Barton, A.M.; Taylor, A.H.J.F. Mixed-Severity Wildfire as a Driver of Vegetation Change in an Arizona Madrean Sky Island System, USA. Fire 2021, 4, 78. [Google Scholar] [CrossRef]
- Lydersen, J.M.; North, M.P.; Collins, B.M. Severity of an uncharacteristically large wildfire, the Rim Fire, in forests with relatively restored frequent fire regimes. For. Ecol. Manag. 2014, 328, 326–334. [Google Scholar] [CrossRef]
- McIver, J.D.; Stephens, S.L.; Agee, J.K.; Barbour, J.; Boerner, R.E.; Edminster, C.B.; Erickson, K.L.; Farris, K.L.; Fettig, C.J.; Fiedler, C.E.J.I.J.o.W.F. Ecological effects of alternative fuel-reduction treatments: Highlights of the National Fire and Fire Surrogate study (FFS). Int. J. Wildland Fire 2012, 22, 63–82. [Google Scholar] [CrossRef]
- Stephens, S.L.; Moghaddas, J.J.; Edminster, C.; Fiedler, C.E.; Haase, S.; Harrington, M.; Keeley, J.E.; Knapp, E.E.; McIver, J.D.; Metlen, K. Fire treatment effects on vegetation structure, fuels, and potential fire severity in western US forests. Ecol. Appl. 2009, 19, 305–320. [Google Scholar] [CrossRef]
- Addington, R.N.; Aplet, G.H.; Battaglia, M.A.; Briggs, J.S.; Brown, P.M.; Cheng, A.S.; Dickinson, Y.; Feinstein, J.A.; Pelz, K.A.; Regan, C.M.; et al. Principles and Practices for the Restoration of Ponderosa Pine and Dry Mixed-Conifer Forests of the Colorado Front Range; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2018; 121p.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Year | Acer grandidentata | Juniperus deppeana | Pinus edulis | Pinus ponderosa | Pinus strobiformis | Pseudotsuga menzesii | Quercus gambelii | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| tree density (# ha−2) | 2004 | 89.2 ± 36.3 | * | 1234.4 ± 200.6 | ns | 684.0 ± 180.9 | * | 1435.9 ± 247.3 | ns | 2748.6 ± 458.4 | ** | 5289.0 ± 578.1 | ** | 4129.2 ± 429.8 | ns | 16,053.0 ± 200.6 | ** |
| 2018 | 23.6 ± 13.0 | 1121.7 ± 178.9 | 475.5 ± 134.3 | 1045.8 ± 181.5 | 2043.0 ± 351.7 | 2814.2 ± 340.7 | 4959.1 ± 653.4 | 12,904.2 ± 178.9 | |||||||||
| basal area (m2 ha−2) | 2004 | 0.1 ± 0.0 | ns | 3.4 ± 0.6 | ns | 96.8 ± 0.3 | ns | 3.3 ± 0.5 | ns | 0.2 ± 1.0 | ns | 8.7 ± 0.0 | ns | 3.4 ± 1.1 | ** | 25.9 ± 0.4 | ** |
| 2018 | 0.1 ± 0.0 | 3.2 ± 0.5 | 88.7 ± 0.2 | 3.0 ± 0.4 | 0.0 ± 0.8 | 7.2 ± 0.0 | 2.1 ± 0.8 | 22.0 ± 0.3 | |||||||||
| Species | Time-Step | JUDE-PIED | PSME-QUGA | PIST-PSME-QUGA | PSME-QUGA-JUDE-PIPO |
|---|---|---|---|---|---|
| Acer grandidentata | pre-fire | 0.0 | 1.3 | 0.1 | 1.3 |
| post-fire | 0.0 | 1.1 | 0.3 | 1.0 | |
| Arbutus xalapensis | pre-fire | 0.7 | 0.1 | 0.0 | 0.0 |
| post-fire | 1.2 | 0.5 | 0.0 | 0.0 | |
| Juniperus deppeana | pre-fire | 42.6 | 15.8 | 12.0 | 33.1 |
| post-fire | 63.2 | 15.5 | 32.8 | 31.2 | |
| Ostrya virginiana | pre-fire | 0.3 | 0.6 | 0.0 | 0.6 |
| post-fire | 0.0 | 2.0 | 0.0 | 0.5 | |
| Pinus edulis | pre-fire | 40.6 | 11.2 | 7.5 | 17.9 |
| post-fire | 33.2 | 4.9 | 1.6 | 23.6 | |
| Pinus ponderosa | pre-fire | 27.0 | 22.4 | 13.9 | 22.9 |
| post-fire | 39.5 | 20.6 | 25.8 | 20.5 | |
| Pinus strobiformis | pre-fire | 21.3 | 25.5 | 62.7 | 27.3 |
| post-fire | 11.1 | 34.3 | 50.8 | 22.6 | |
| Prunus serotina | pre-fire | 0.6 | 0.2 | 0.0 | 0.4 |
| post-fire | 0.2 | 0.2 | 0.8 | 0.9 | |
| Pseudotsuga menzeisii | pre-fire | 36.2 | 67.4 | 54.4 | 55.3 |
| post-fire | 28.6 | 64.5 | 48.5 | 37.9 | |
| Quercus gambelii | pre-fire | 30.6 | 52.5 | 48.2 | 37.8 |
| post-fire | 18.4 | 49.4 | 38.3 | 49.2 | |
| Quercus meuhlembergii | pre-fire | 0.0 | 1.1 | 0.0 | 1.4 |
| post-fire | 4.5 | 0.6 | 0.6 | 1.4 | |
| Robinia neomexicana | pre-fire | 0.1 | 0.4 | 0.4 | 0.4 |
| post-fire | 0.0 | 1.0 | 0.0 | 2.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakulich, J.; Poulos, H.M.; Gatewood, R.G.; Wogan, K.A.; Marks, C.; Taylor, A.H. Low-Severity Wildfire Shifts Mixed Conifer Forests toward Historical Stand Structure in Guadalupe Mountains National Park, Texas, USA. Fire 2022, 5, 119. https://doi.org/10.3390/fire5040119
Sakulich J, Poulos HM, Gatewood RG, Wogan KA, Marks C, Taylor AH. Low-Severity Wildfire Shifts Mixed Conifer Forests toward Historical Stand Structure in Guadalupe Mountains National Park, Texas, USA. Fire. 2022; 5(4):119. https://doi.org/10.3390/fire5040119
Chicago/Turabian StyleSakulich, John, Helen M. Poulos, Richard G. Gatewood, Kelsey A. Wogan, Christopher Marks, and Alan H. Taylor. 2022. "Low-Severity Wildfire Shifts Mixed Conifer Forests toward Historical Stand Structure in Guadalupe Mountains National Park, Texas, USA" Fire 5, no. 4: 119. https://doi.org/10.3390/fire5040119
APA StyleSakulich, J., Poulos, H. M., Gatewood, R. G., Wogan, K. A., Marks, C., & Taylor, A. H. (2022). Low-Severity Wildfire Shifts Mixed Conifer Forests toward Historical Stand Structure in Guadalupe Mountains National Park, Texas, USA. Fire, 5(4), 119. https://doi.org/10.3390/fire5040119