
Simulation Study of an Abstract Forest Ecosystem with Multi-Species under Lightning-Caused Fires


Abstract
:1. Introduction
2. Model
2.1. Forest Environment
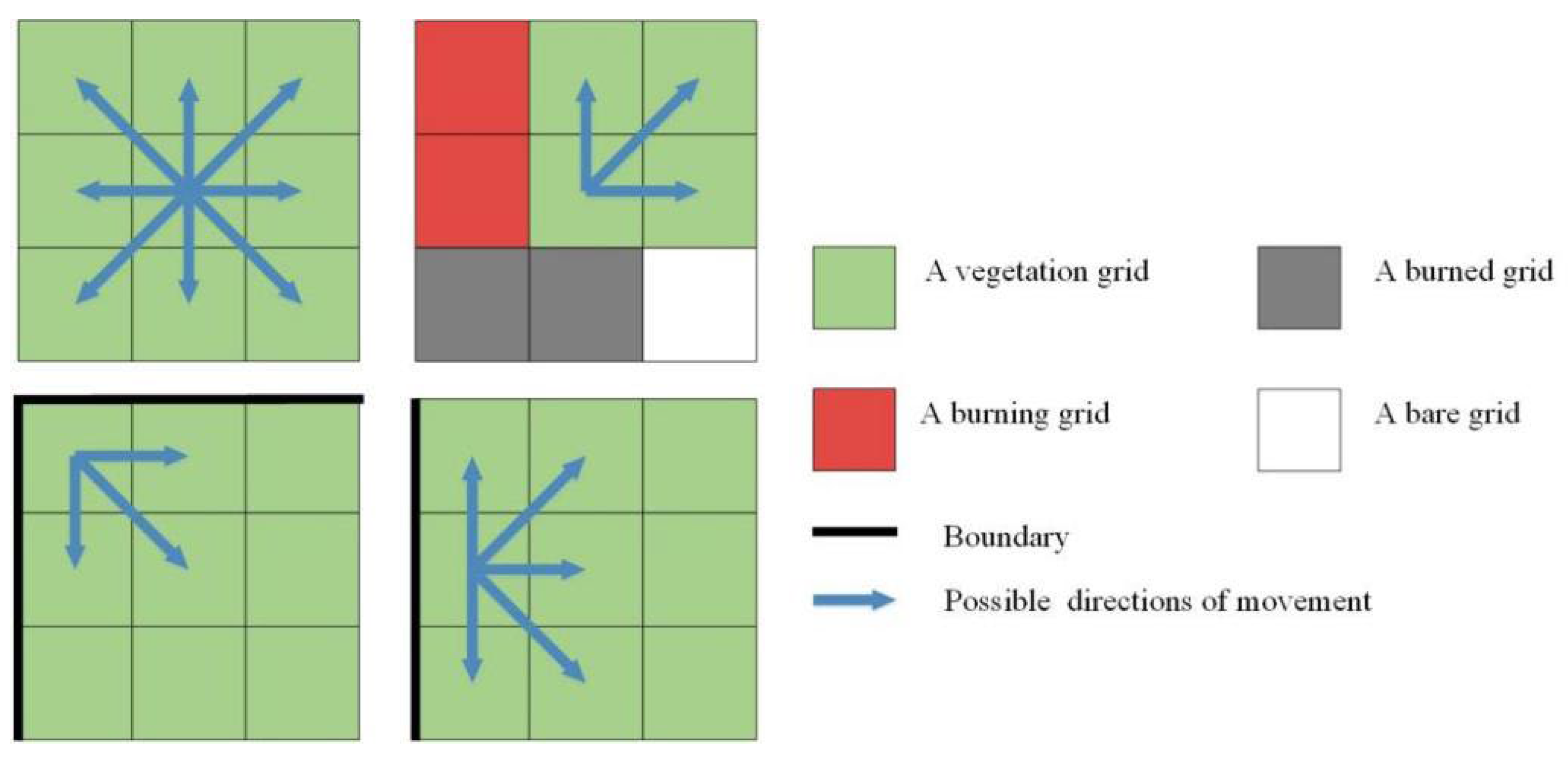
2.2. Lightning-Caused Fire Agents
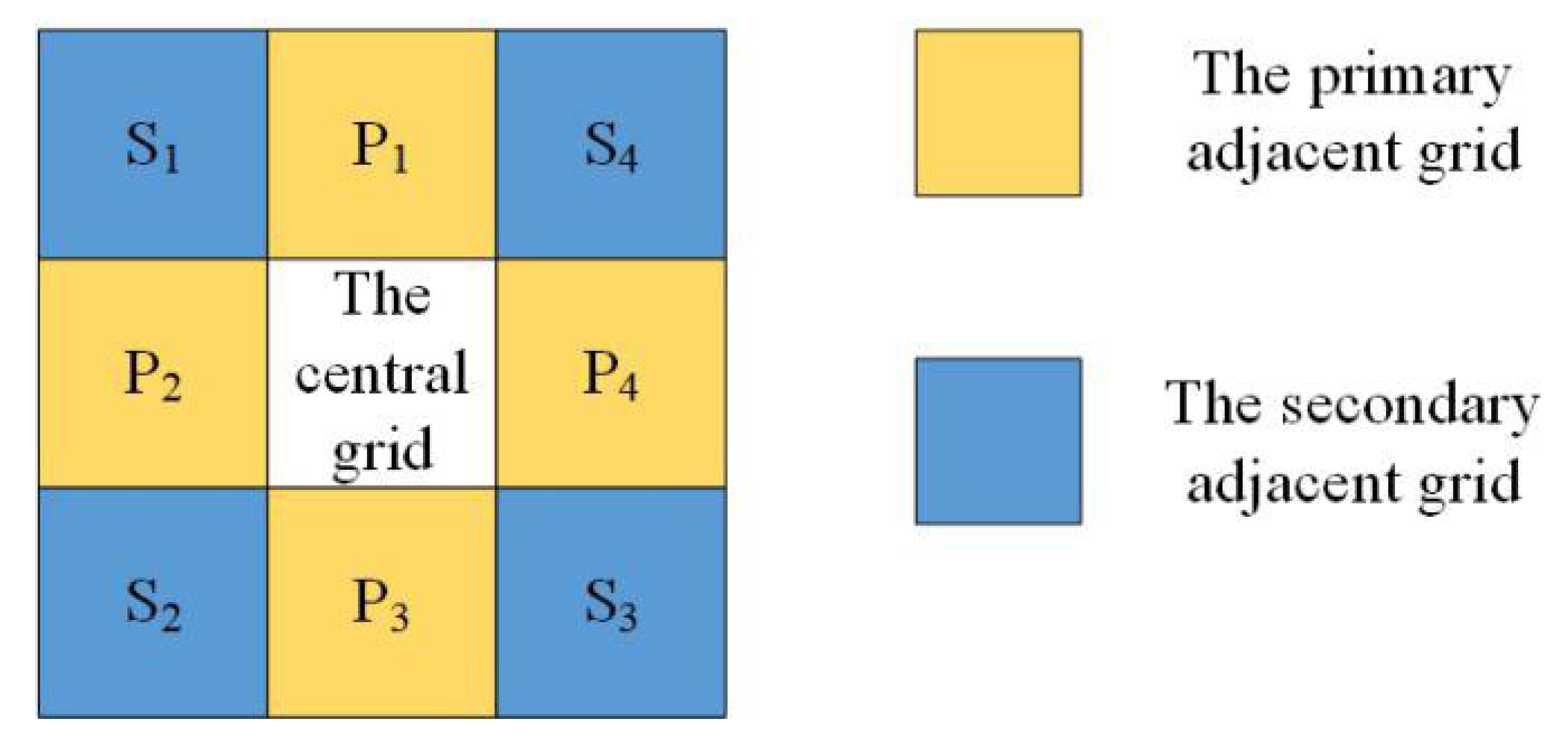
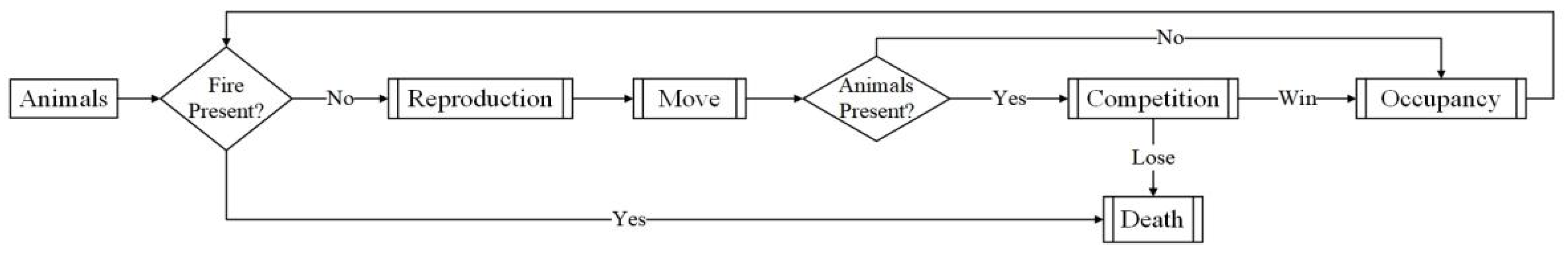
2.3. Species Agents
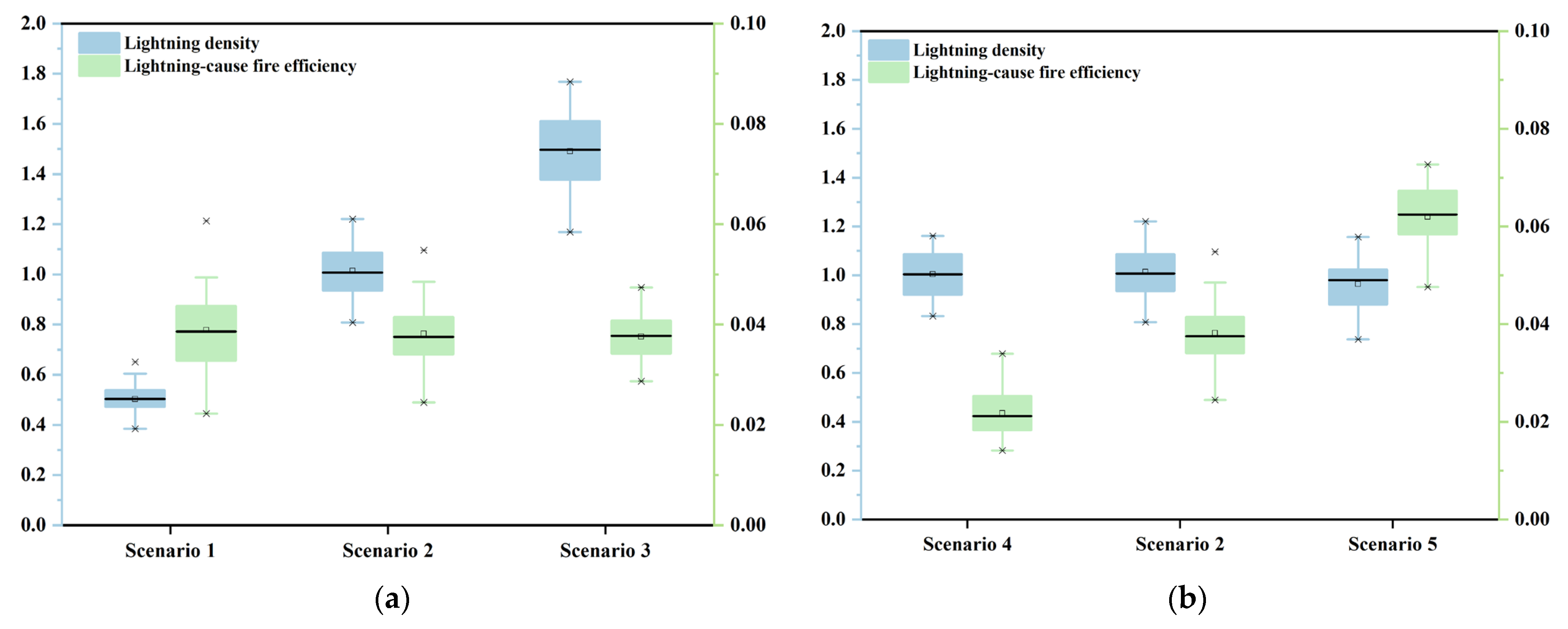
3. Simulation Experiments
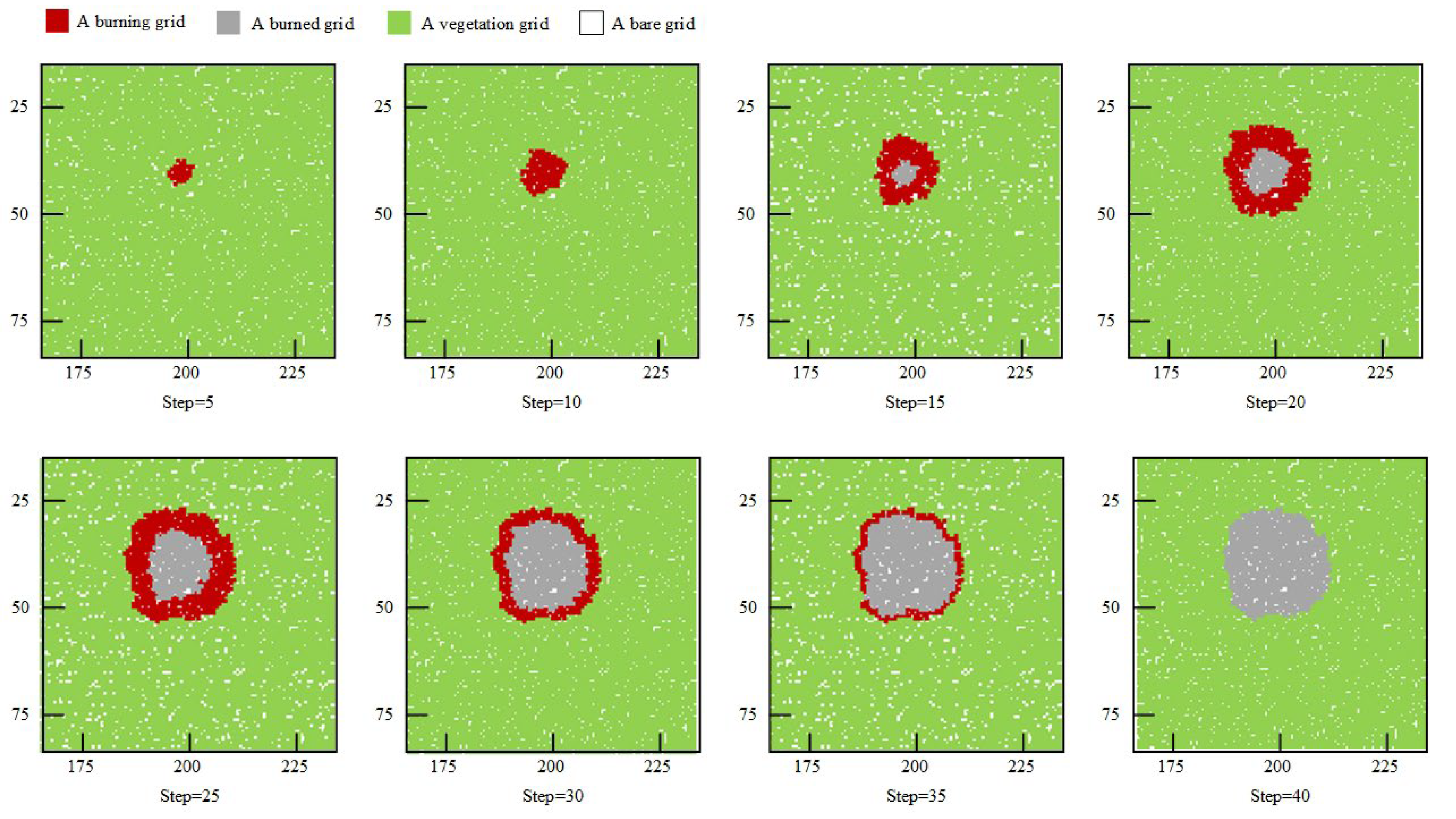
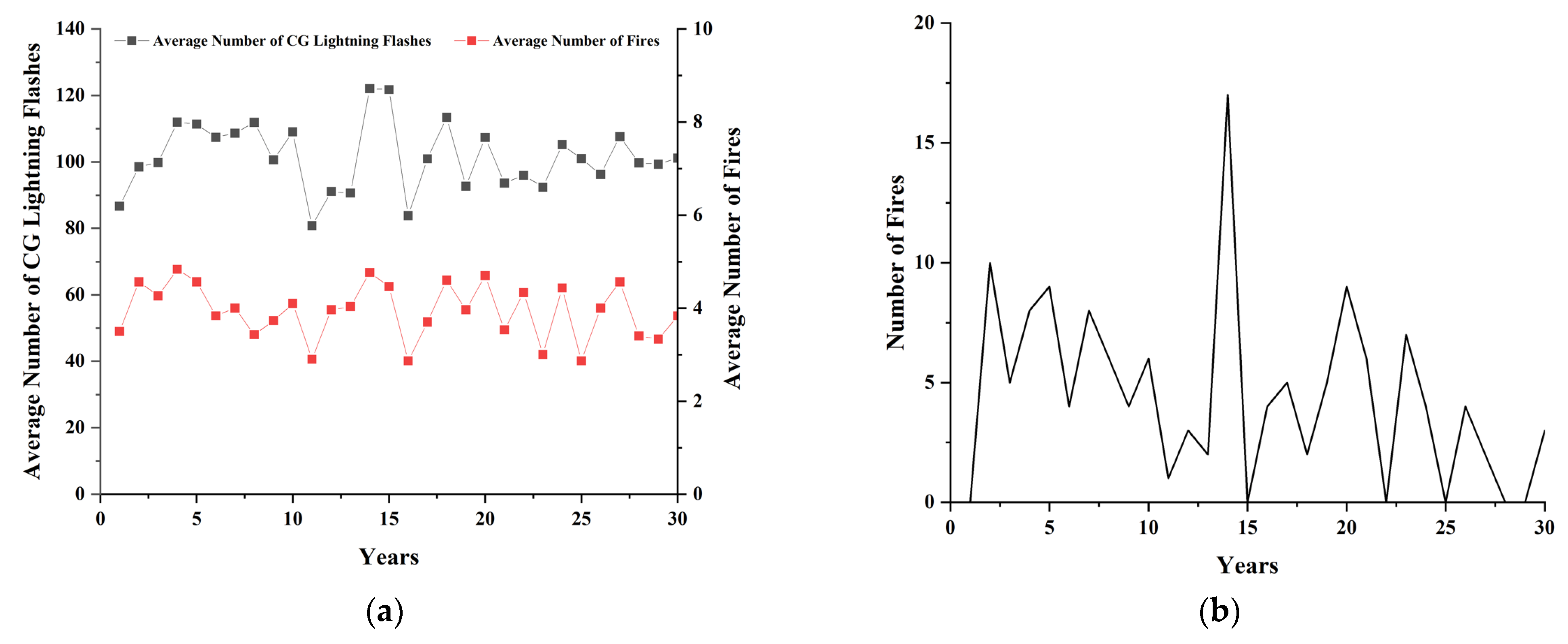
4. Model Validation
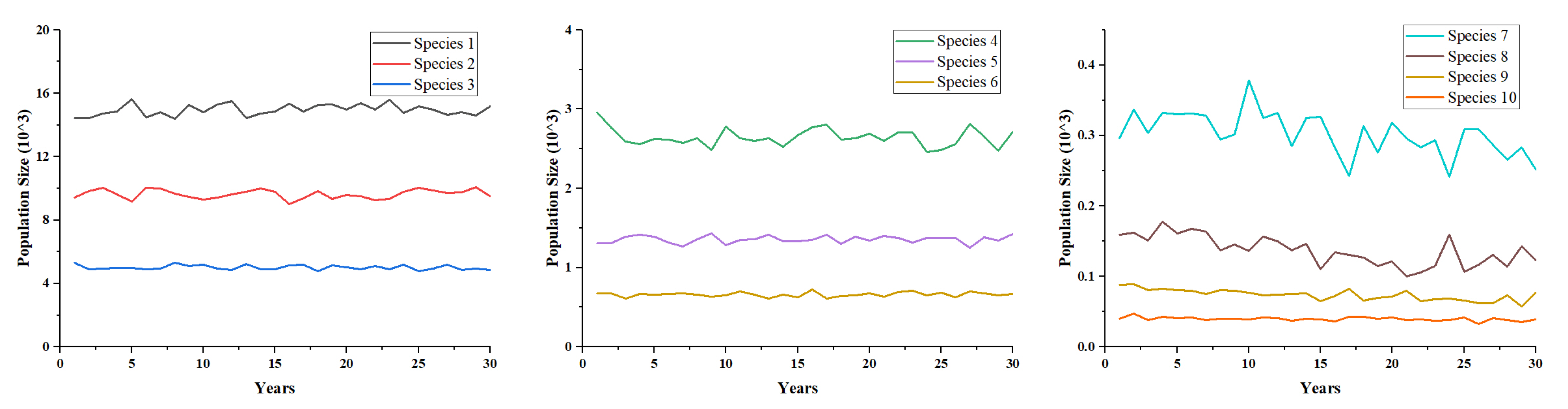
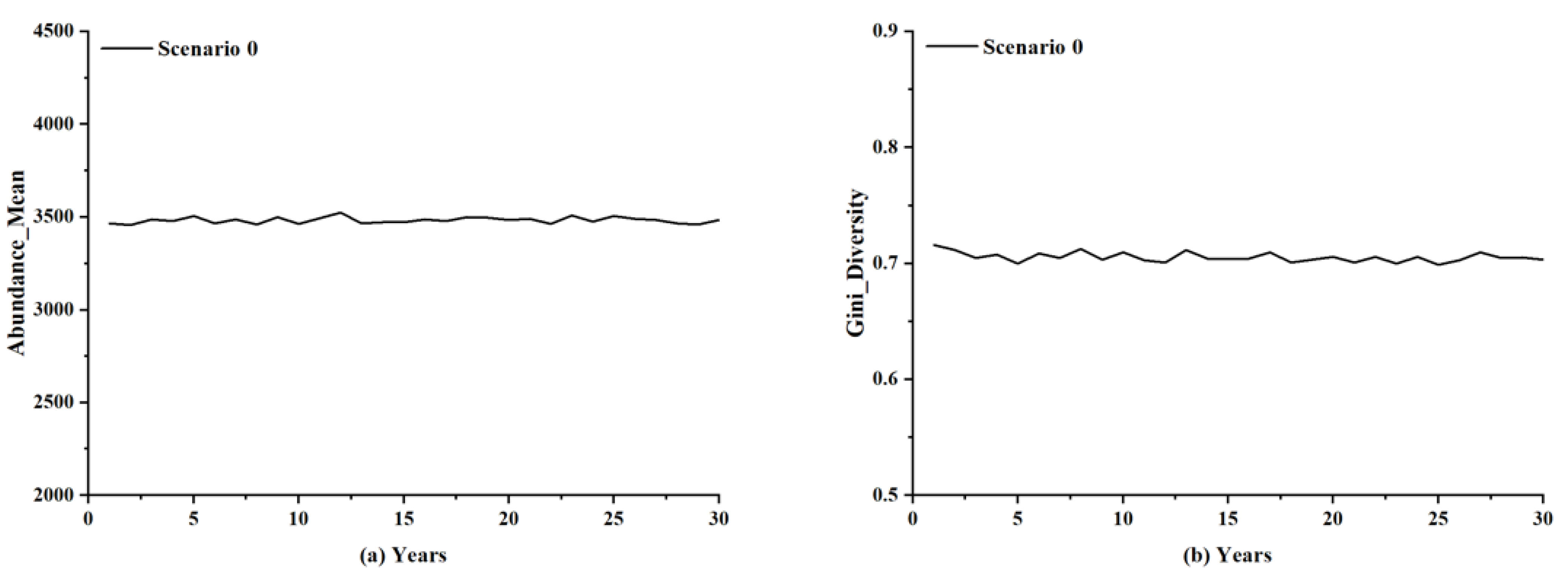
5. Simulation Results
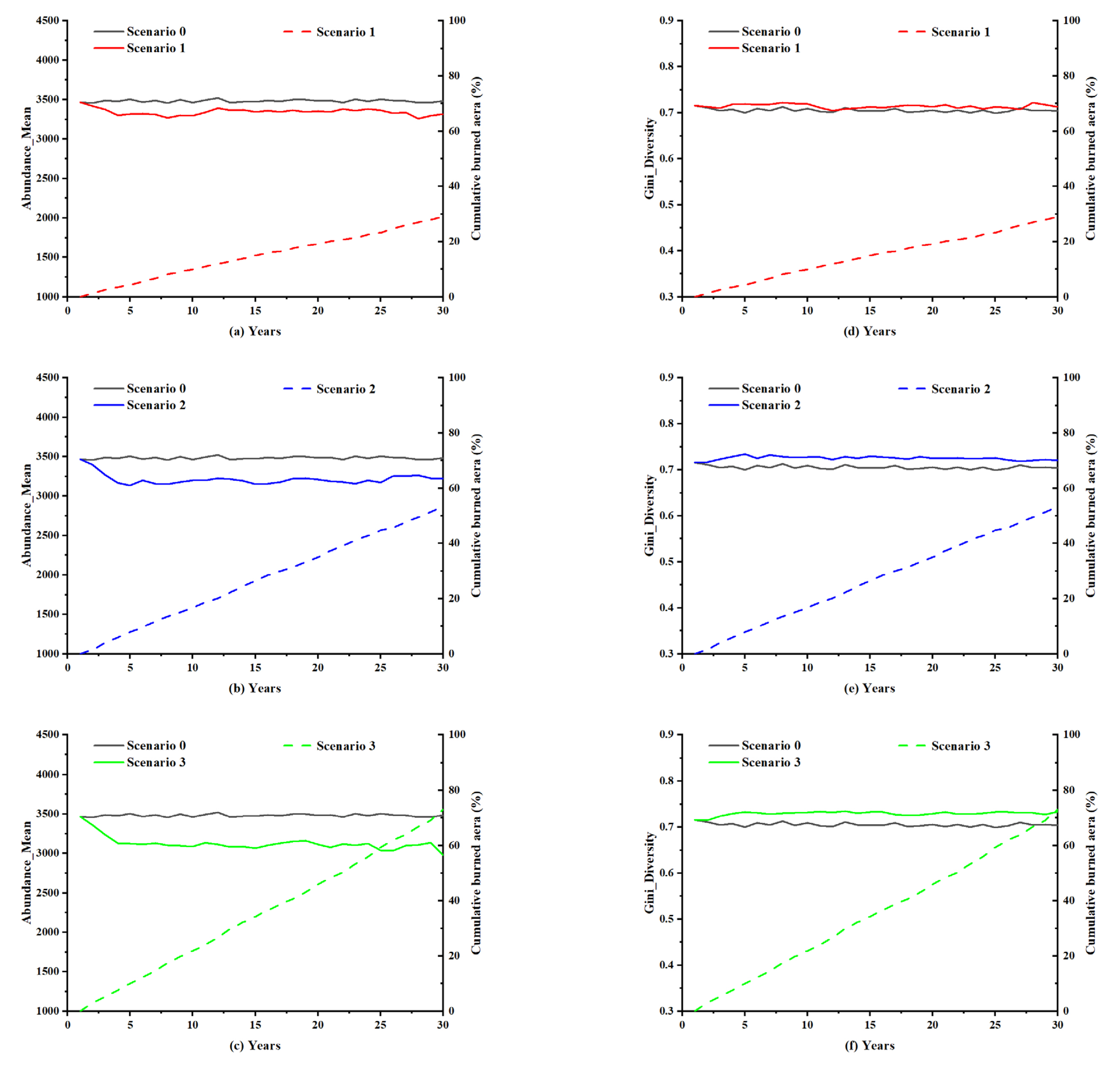
5.1. Insights into the Effect of Lightning-Caused Fires on the Forest Ecosystem under Different Lightning Flash Densities
5.1.1. Burned Area
5.1.2. Species Diversity
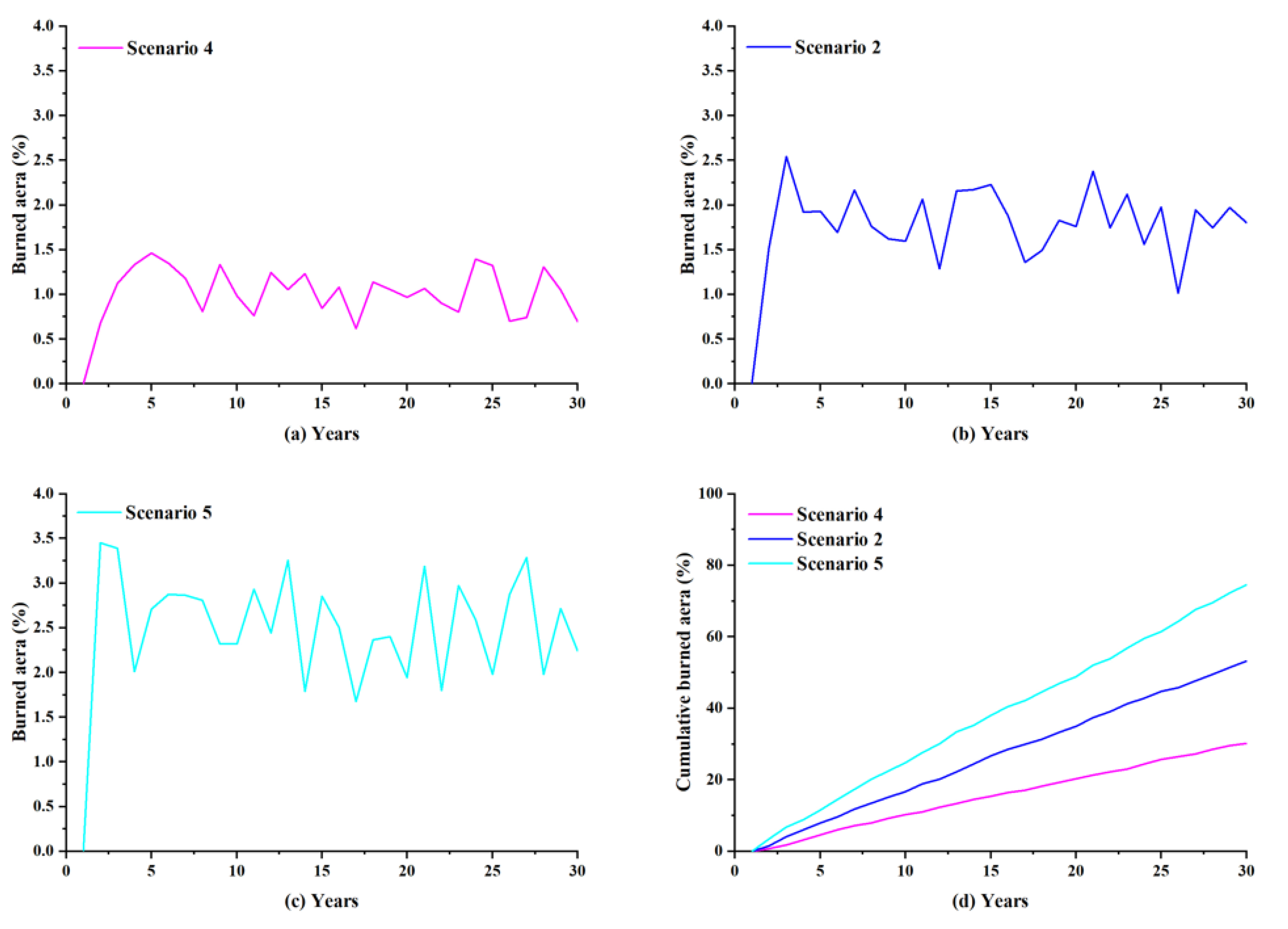
5.2. Insights into the Effect of Lightning-Caused Fires on the Forest Ecosystem under Different Lightning-Caused Fire Efficiencies
5.2.1. Burned Area
5.2.2. Species Diversity
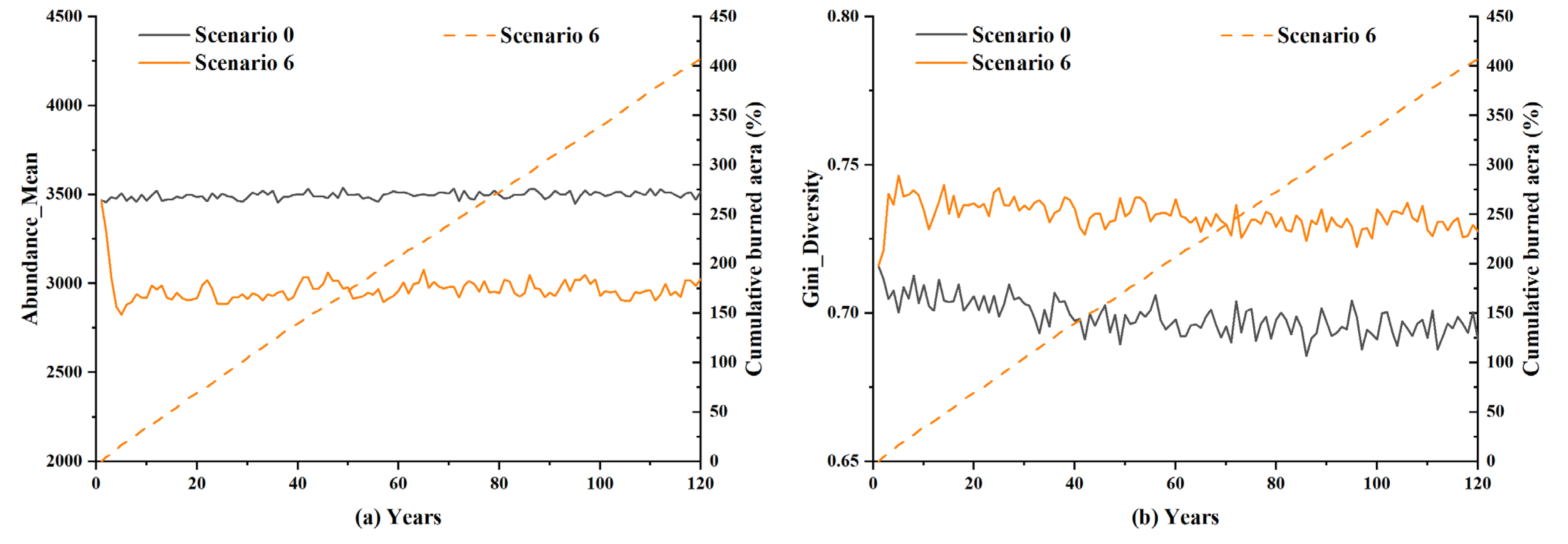
5.3. Insights into the Overall Impact of Lightning-Caused Fires on the Forest Ecosystem
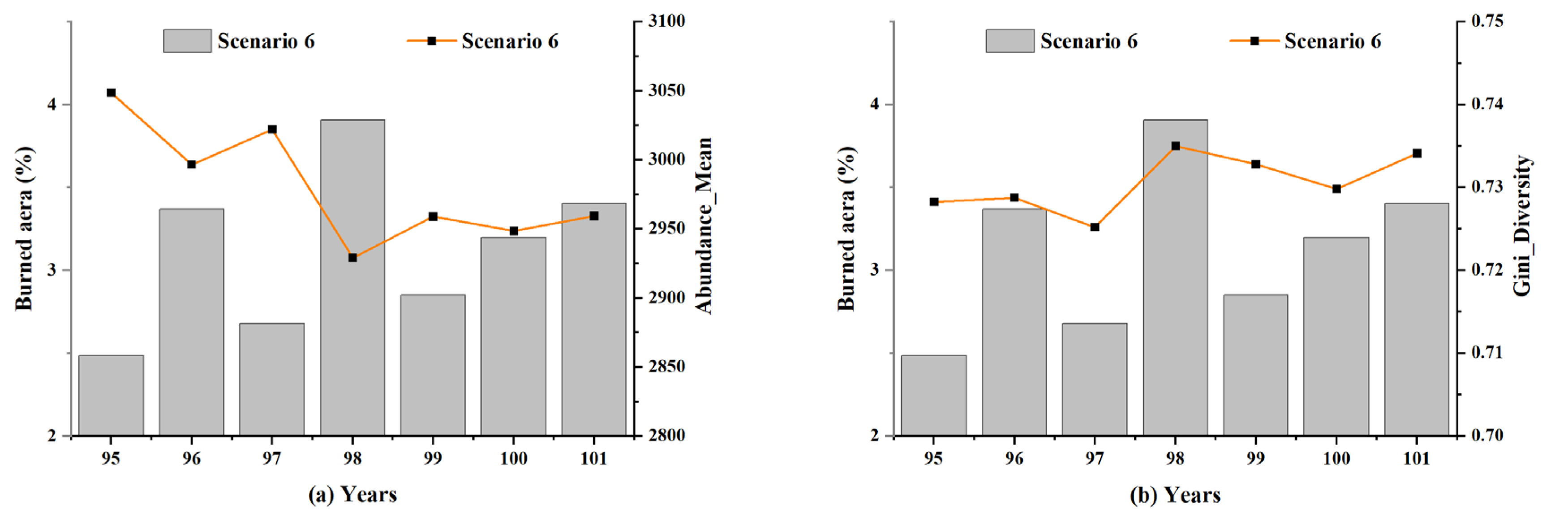
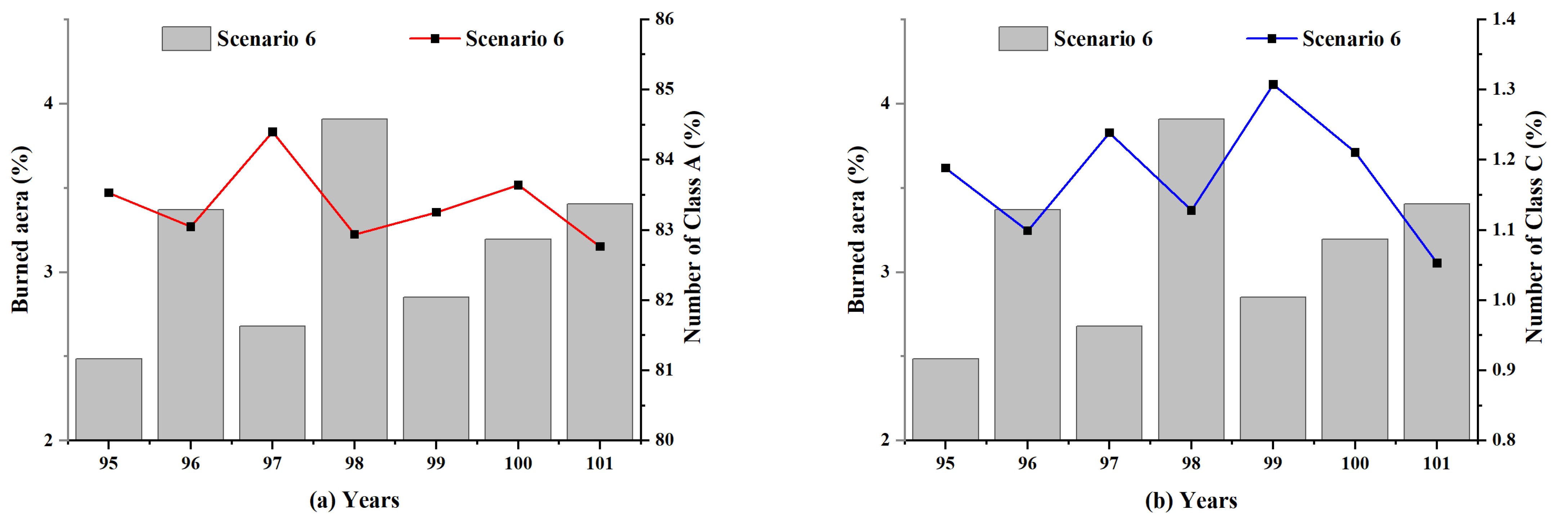
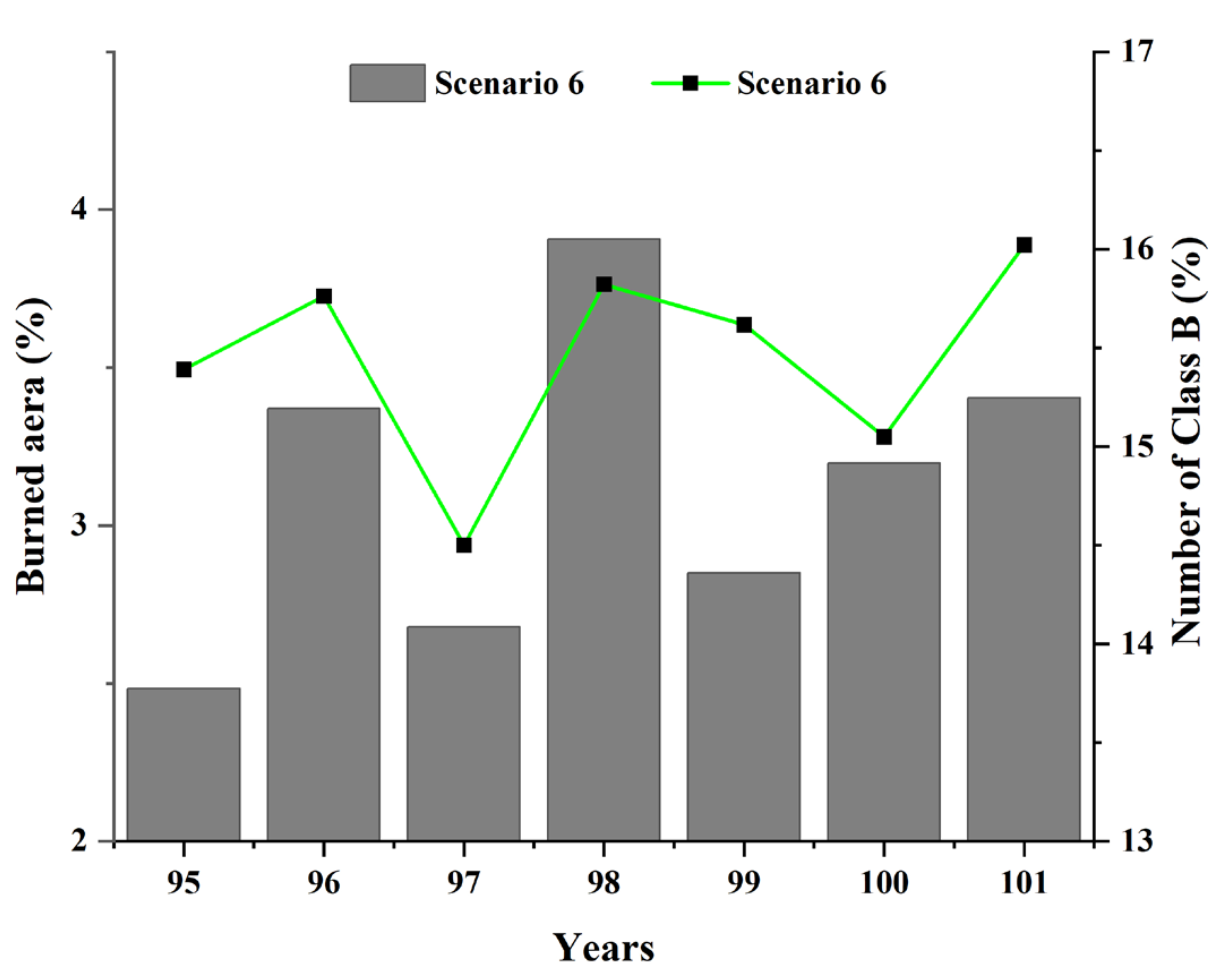
5.4. Local Insight into the Influence of Lightning-Caused Fires on Ecological Diversity
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fahey, T.J. Forest Ecology. In Encyclopedia of Biodiversity; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 528–536. [Google Scholar]
- Oris, F.; Asselin, H.; Ali, A.A.; Finsinger, W.; Bergeron, Y. Effect of increased fire activity on global warming in the boreal forest. Environ. Rev. 2014, 22, 206–219. [Google Scholar] [CrossRef] [Green Version]
- Spies, T.A.; White, E.M.; Kline, J.D.; Fischer, A.P.; Ager, A.; Bailey, J.; Bolte, J.; Koch, J.; Platt, E.; Olsen, C.S. Examining fire-prone forest landscapes as coupled human and natural systems. Ecol. Soc. 2014, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Ganteaume, A.; Syphard, A.D. Ignition Sources. In Encyclopedia of Wildfires and Wildland-Urban Interface (WUI) Fires; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Flannigan, M.D.; Wotton, B.M. Lightning-ignited forest fires in northwestern Ontario. Can. J. For. Res. 1991, 21, 277–287. [Google Scholar] [CrossRef]
- Stocks, B.J.; Mason, J.A.; Todd, J.B.; Bosch, E.M.; Wotton, B.M.; Amiro, B.D.; Flannigan, M.D.; Hirsch, K.G.; Logan, K.A.; Martell, D.L. Large forest fires in Canada, 1959–1997. J. Geophys. Res. Atmos. 2002, 107, 5. [Google Scholar] [CrossRef]
- Van Wagtendonk, J.W. The history and evolution of wildland fire use. Fire Ecol. 2007, 3, 3–17. [Google Scholar] [CrossRef]
- Cohen, D.; Dellinger, B.; Klein, R.; Buchanan, B. Patterns in lightning-caused fires at Great Smoky Mountains National Park. Fire Ecol. 2007, 3, 68–82. [Google Scholar] [CrossRef]
- Anderson, K. A model to predict lightning-caused fire occurrences. Int. J. Wildland Fire 2002, 11, 163–172. [Google Scholar] [CrossRef]
- Soler, A.; Pineda, N.; San Segundo, H.; Bech, J.; Montanyà, J. Characterisation of thunderstorms that caused lightning-ignited wildfires. Int. J. Wildland Fire 2021, 30, 954–970. [Google Scholar] [CrossRef]
- Pineda, N.; Altube, P.; Alcasena, F.J.; Casellas, E.; San Segundo, H.; Montanyà, J. Characterising the holdover phase of lightning-ignited wildfires in Catalonia. Agric. For. Meteorol. 2022, 324, 109–111. [Google Scholar] [CrossRef]
- Read, N.; Duff, T.J.; Taylor, P.G. A lightning-caused wildfire ignition forecasting model for operational use. Agric. For. Meteorol. 2018, 253, 233–246. [Google Scholar] [CrossRef]
- Dissing, D.; Verbyla, D.L. Spatial patterns of lightning strikes in interior Alaska and their relations to elevation and vegetation. Can. J. For. Res. 2003, 33, 770–782. [Google Scholar] [CrossRef]
- Enno, S.; Sugier, J.; Alber, R.; Seltzer, M. Lightning flash density in Europe based on 10 years of ATDnet data. Atmos. Res. 2020, 235, 104769. [Google Scholar] [CrossRef]
- Pineda, N.; Rigo, T. The rainfall factor in lightning-ignited wildfires in Catalonia. Agric. For. Meteorol. 2017, 239, 249–263. [Google Scholar] [CrossRef]
- Duff, T.J.; Keane, R.E.; Penman, T.D.; Tolhurst, K.G. Revisiting wildland fire fuel quantification methods: The challenge of understanding a dynamic, biotic entity. Forest 2017, 8, 351. [Google Scholar]
- Johnson, E.A.; Miyanishi, K. Forest Fires: Behavior and Ecological Effects; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Latham, D.; Williams, E. Lightning and Forest Fires; Elsevier: Amsterdam, The Netherlands, 2001; pp. 375–418. [Google Scholar]
- Núñez-Regueira, L.; Rodríguez-Añón, J.A.; Proupín-Castiñeiras, J. Design of risk index maps as a tool to prevent forest fires in the humid Atlantic zone of Galicia (NW Spain). Thermochim. Acta 2000, 349, 103–119. [Google Scholar] [CrossRef]
- Podur, J.; Martell, D.L.; Csillag, F. Spatial patterns of lightning-caused forest fires in Ontario, 1976–1998. Ecol. Model. 2003, 164, 1–20. [Google Scholar] [CrossRef]
- Pineda, N.; Montanyà, J.; Van der Velde, O.A. Characteristics of lightning related to wildfire ignitions in Catalonia. Atmos. Res. 2014, 135, 380–387. [Google Scholar] [CrossRef]
- Seidl, R.; Fernandes, P.M.; Fonseca, T.F.; Gillet, F.; Jönsson, A.M.; Merganičová, K.; Netherer, S.; Arpaci, A.; Bontemps, J.; Bugmann, H. Modelling natural disturbances in forest ecosystems: A review. Ecol. Model. 2011, 222, 903–924. [Google Scholar]
- Hulse, D.; Branscomb, A.; Enright, C.; Johnson, B.; Evers, C.; Bolte, J.; Ager, A. Anticipating surprise: Using agent-based alternative futures simulation modeling to identify and map surprising fires in the Willamette Valley, Oregon USA. Landsc. Urban Plann. 2016, 156, 26–43. [Google Scholar]
- Green, D.G.; Gill, A.M.; Noble, I.R. Fire shapes and the adequacy of fire-spread models. Ecol. Model. 1983, 20, 33–45. [Google Scholar] [CrossRef]
- Dong, L.; Leung, L.R.; Qian, Y.; Zou, Y.; Song, F.; Chen, X. Meteorological environments associated with California wildfires and their potential roles in wildfire changes during 1984–2017. J. Geophys. Res. Atmos. 2021, 126, e2020JD033180. [Google Scholar] [CrossRef]
- Abrams, M.D. Ecological and ecophysiological attributes and responses to fire in eastern oak forests. In Fire in Eastern Oak Forests: Delivering Science to Land Managers Conference, Columbus, OH, USA, 15–17 November 2005; The Ohio State University: Columbus, OH, USA, 2005; pp. 74–89. [Google Scholar]
- Rothermel, R.C. A Mathematical Model for Predicting Fire Spread in Wildland Fuels; Intermountain Forest & Range Experiment Station, Forest Service, US: Ogden, UT, USA, 1972; Volume 115. [Google Scholar]
- McArthur, A.G. Fire Behaviour in Eucalypt Forests; Commonwealth of Australia Forestry and Timber Bureau: Canberra, Australia, 1967. [Google Scholar]
- Wang, Z. Current forest fire danger rating system. J. Nat. Disasters 1992, 3, 39–44. [Google Scholar]
- Mandel, J.; Amram, S.; Beezley, J.D.; Kelman, G.; Kochanski, A.K.; Kondratenko, V.Y.; Lynn, B.H.; Regev, B.; Vejmelka, M. Recent advances and applications of WRF&SFIRE. Nat. Hazards Earth Syst. Sci. 2014, 14, 2829–2845. [Google Scholar]
- Tang, W.; Bennett, D.A. Agent-based modeling of animal movement: A review. Geogr. Compass 2010, 4, 682–700. [Google Scholar]
- Hu, X.; Sun, Y. Agent-Based Modeling and Simulation of Wildland Fire Suppression. In Proceedings of the 2007 Winter Simulation Conference, Washington, DC, USA, 9–12 December 2007; IEEE: Piscataway, NJ, USA, 2007; pp. 1275–1283. [Google Scholar]
- Bone, C.; Johnson, B.; Nielsen-Pincus, M.; Sproles, E.; Bolte, J. A Temporal Variant-invariant Validation Approach for Agent-based Models of Landscape Dynamics. Trans. GIS 2014, 18, 161–182. [Google Scholar]
- Ager, A.A.; Barros, A.M.; Day, M.A.; Preisler, H.K.; Spies, T.A.; Bolte, J. Analyzing fine-scale spatiotemporal drivers of wildfire in a forest landscape model. Ecol. Model. 2018, 384, 87–102. [Google Scholar]
- Céspedes, B.; Luna, B.; Pérez, B.; Urbieta, I.R.; Moreno, J.M. Burning season effects on the short-term post-fire vegetation dynamics of a Mediterranean heathland. Appl. Veg. Sci. 2014, 17, 86–96. [Google Scholar] [CrossRef]
- Luo, C.; Shen, Z.; Li, Y.; Han, J.; Pausas, J.G.; Xu, Q.; Zhang, Z.; Jiang, Y. Determinants of post–fire regeneration demography in a subtropical monsoon–climate forest in Southwest China. Sci. Total Environ. 2021, 766, 142605. [Google Scholar] [CrossRef]
- Ganey, J.L.; Wan, H.Y.; Cushman, S.A.; Vojta, C.D. Conflicting perspectives on spotted owls, wildfire, and forest restoration. Fire Ecol. 2017, 13, 146–165. [Google Scholar]
- Hutchen, J.; Hodges, K.E. Impact of wildfire size on snowshoe hare relative abundance in southern British Columbia, Canada. Fire Ecol. 2019, 15, 37. [Google Scholar]
- Morales, A.M.; Politi, N.; Rivera, L.O.; Vivanco, C.G.; Defossé, G.E. Fire and distance from unburned forest influence bird assemblages in Southern Andean Yungas of Northwest Argentina: A case study. Fire Ecol. 2020, 16, 10. [Google Scholar]
- Ferrenberg, S.M.; Schwilk, D.W.; Knapp, E.E.; Groth, E.; Keeley, J.E. Fire decreases arthropod abundance but increases diversity: Early and late season prescribed fire effects in a Sierra Nevada mixed-conifer forest. Fire Ecol. 2006, 2, 79–102. [Google Scholar]
- Karsai, I.; Roland, B.; Kampis, G. The effect of fire on an abstract forest ecosystem: An agent based study. Ecol. Complex 2016, 28, 12–23. [Google Scholar]
- Karsai, I.; Schmickl, T.; Kampis, G.; Karsai, I.; Schmickl, T.; Kampis, G. Forest Fires: Fire Management and the Power Law. In Resilience and Stability of Ecological and Social Systems; Springer: Cham, Switzerland, 2020; pp. 63–77. [Google Scholar]
- Foster, C.N.; Banks, S.C.; Cary, G.J.; Johnson, C.N.; Lindenmayer, D.B.; Valentine, L.E. Animals as agents in fire regimes. Trends Ecol. Evol. 2020, 35, 346–356. [Google Scholar] [PubMed]
- Tesson, S.V.; Edelaar, P. Dispersal in a changing world: Opportunities, insights and challenges. Mov. Ecol. 2013, 1, 10. [Google Scholar]
- Okubo, A.; Levin, S.A. Diffusion and Ecological Problems: Modern Perspectives; Springer: New York, NY, USA, 2001. [Google Scholar]
- Bennett, D.A.; Tang, W. Modelling adaptive, spatially aware, and mobile agents: Elk migration in Yellowstone. Int. J. Geogr. Inform. Sci 2006, 20, 1039–1066. [Google Scholar]
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar]
- Dennhardt, A.J.; Duerr, A.E.; Brandes, D.; Katzner, T.E. Modeling autumn migration of a rare soaring raptor identifies new movement corridors in central Appalachia. Ecol. Model. 2015, 303, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Schoon, M.; Baggio, J.A.; Salau, K.R.; Janssen, M. Insights for managers from modeling species interactions across multiple scales in an idealized landscape. Environ. Model. Softw. 2014, 54, 53–59. [Google Scholar] [CrossRef]
- Hidalgo, J.; Suweis, S.; Maritan, A. Species coexistence in a neutral dynamics with environmental noise. J. Theor. Biol. 2017, 413, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Haugo, R.; Zanger, C.; DeMeo, T.; Ringo, C.; Shlisky, A.; Blankenship, K.; Simpson, M.; Mellen-McLean, K.; Kertis, J.; Stern, M. A new approach to evaluate forest structure restoration needs across Oregon and Washington, USA. For. Ecol. Manag. 2015, 335, 37–50. [Google Scholar]
- Trunfio, G.A. Predicting Wildfire Spreading through a Hexagonal Cellular Automata Model. In Proceedings of the 6th International Conference on Cellular Automata for Research and Industry, ACRI 2004, Amsterdam, The Netherlands, 25–28 October 2004; Springer: Berlin/Heidelberg, Germany, 2004; pp. 385–394. [Google Scholar]
- Almeida, R.M.; Macau, E.E. Stochastic Cellular Automata Model for Wildland fire SPREAD Dynamics. J. Phys. Conf. Ser. 2011, 285, 12038. [Google Scholar] [CrossRef]
- Yang, X.; Sun, J.; Li, W. An analysis of cloud-to-ground lightning in China during 2010–13. Weather Forecast 2015, 30, 1537–1550. [Google Scholar] [CrossRef]
- Koehler, T.L. Cloud-to-ground lightning flash density and thunderstorm day distributions over the contiguous United States derived from NLDN measurements: 1993–2018. Mon. Weather Rev. 2020, 148, 313–332. [Google Scholar]
- Hessilt, T.D.; Abatzoglou, J.T.; Chen, Y.; Randerson, J.T.; Scholten, R.C.; Van Der Werf, G.; Veraverbeke, S. Future increases in lightning ignition efficiency and wildfire occurrence expected from drier fuels in boreal forest ecosystems of western North America. Environ. Res. Lett. 2022, 17, 54008. [Google Scholar]
- Green, D.G.; House, A.; House, S.M. Simulating spatial patterns in forest ecosystems. Math. Comput. Simulat. 1985, 27, 191–198. [Google Scholar] [CrossRef]
- Wang, X.; Liu, C.; Liu, J.; Qin, X.; Wang, N.; Zhou, W. A Cellular Automata Model for Forest Fire Spreading Simulation. In Proceedings of the 2016 IEEE Symposium Series on Computational Intelligence (SSCI), Athens, Greece, 6–9 December 2016; IEEE: Piscataway, NJ, USA, 2016; pp. 1–6. [Google Scholar]
- Ramsay, P.M.; Oxley, E. Fire temperatures and postfire plant community dynamics in Ecuadorian grass páramo. Vegetatio 1996, 124, 129–144. [Google Scholar] [CrossRef]
- Konstantinidis, P.; Tsiourlis, G.; Xofis, P. Effect of fire season, aspect and pre-fire plant size on the growth of Arbutus unedo L. (strawberry tree) resprouts. For. Ecol. Manag. 2006, 225, 359–367. [Google Scholar] [CrossRef]
- Couto, F.T.; Iakunin, M.; Salgado, R.; Pinto, P.; Viegas, T.; Pinty, J. Lightning modelling for the research of forest fire ignition in Portugal. Atmos. Res. 2020, 242, 104993. [Google Scholar] [CrossRef]
- Los Huertos, M. Ecology and Management of Inland Waters: A Californian Perspective with Global Applications; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Bampoh, D.; Earl, J.E.; Zollner, P.A. Examining the relative influence of animal movement patterns and mortality models on the distribution of animal transported subsidies. Ecol. Model. 2019, 412, 108824. [Google Scholar]
- Peet, R.K. The measurement of species diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Lan, B.L.; Yeoh, E.V.; Ng, J.A. Distribution of detrended stock market data. Fluct. Noise Lett. 2010, 9, 245–257. [Google Scholar] [CrossRef]
- Looman, J. Biological equilibrium in ecosystems 1. A theory of biological equilibrium. Folia Geobot. Phytotaxon. 1976, 11, 1–21. [Google Scholar] [CrossRef]
- Ramos-Neto, M.B.; Pivello, V.R. Lightning fires in a Brazilian savanna national park: Rethinking management strategies. Environ. Manag. 2000, 26, 675–684. [Google Scholar]
- Granström, A. Spatial and temporal variation in lightning ignitions in Sweden. J. Veg. Sci. 1993, 4, 737–744. [Google Scholar] [CrossRef]
- Wotton, B.M.; Martell, D.L. A lightning fire occurrence model for Ontario. Can. J. For. Res. 2005, 35, 1389–1401. [Google Scholar] [CrossRef]
- Wierzchowski, J.; Heathcott, M.; Flannigan, M.D. Lightning and lightning fire, central cordillera, Canada. Int. J. Wildland Fire 2002, 11, 41–51. [Google Scholar] [CrossRef]
- Apigian, K.O.; Dahlsten, D.L.; Stephens, S.L. Fire and fire surrogate treatment effects on leaf litter arthropods in a western Sierra Nevada mixed-conifer forest. For. Ecol. Manag. 2006, 221, 110–122. [Google Scholar] [CrossRef]
- Pausas, J.G.; Parr, C.L. Towards an understanding of the evolutionary role of fire in animals. Evol. Ecol. 2018, 32, 113–125. [Google Scholar] [CrossRef]
- Bouchard, M.; Kneeshaw, D.; Bergeron, Y. Ecosystem management based on large-scale disturbance pulses: A case study from sub-boreal forests of western Quebec (Canada). For. Ecol. Manag. 2008, 256, 1734–1742. [Google Scholar]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar]
- Guo, F.; Su, Z.; Wang, G.; Sun, L.; Lin, F.; Liu, A. Wildfire ignition in the forests of southeast China: Identifying drivers and spatial distribution to predict wildfire likelihood. Appl. Geogr. 2016, 66, 12–21. [Google Scholar]
- Baranovskiy, N. Deterministic-Probabilistic Approach to Predict Lightning-Caused Forest Fires in Mounting Areas. Forecasting 2021, 3, 695–715. [Google Scholar] [CrossRef]
- Ganteaume, A.; Camia, A.; Jappiot, M.; San-Miguel-Ayanz, J.; Long-Fournel, M.; Lampin, C. A review of the main driving factors of forest fire ignition over Europe. Env. Manag. 2013, 51, 651–662. [Google Scholar]
- Díaz-Avalos, C.; Peterson, D.L.; Alvarado, E.; Ferguson, S.A.; Besag, J.E. Space time modelling of lightning-caused ignitions in the Blue Mountains, Oregon. Can. J. For. Res. 2001, 31, 1579–1593. [Google Scholar]
- Knapp, E.E.; Keeley, J.E. Heterogeneity in fire severity within early season and late season prescribed burns in a mixed-conifer forest. Int. J. Wildland Fire 2006, 15, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.M.; Vacik, H. Characteristics of lightnings igniting forest fires in Austria. Agric. For. Meteorol. 2017, 240, 26–34. [Google Scholar] [CrossRef]
- Vecín-Arias, D.; Castedo-Dorado, F.; Ordóñez, C.; Rodríguez-Pérez, J.R. Biophysical and lightning characteristics drive lightning-induced fire occurrence in the central plateau of the Iberian Peninsula. Agric. For. Meteorol. 2016, 225, 36–47. [Google Scholar]
- Dowdy, A.J.; Fromm, M.D.; McCarthy, N. Pyrocumulonimbus lightning and fire ignition on Black Saturday in southeast Australia. J. Geophys. Res. Atmos. 2017, 122, 7342–7354. [Google Scholar] [CrossRef]
- Stawski, C.; Körtner, G.; Nowack, J.; Geiser, F. The importance of mammalian torpor for survival in a post-fire landscape. Biol. Lett. 2015, 11, 20150134. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scenario | Explanation | Forest Scenario Represented |
|---|---|---|
| Scenario 0 | Lightning density = 0, lightning-caused fire efficiency = 0 | No fire |
| Scenario 1 | Lightning density U(0,1), lightning-caused fire efficiency U(0.03, 0.05) | Low lightning susceptibility, Medium flammability; |
| Scenario 2 | Lightning density U(0,2), lightning-caused fire efficiency U(0.03, 0.05) | Medium lightning susceptibility, Medium flammability; |
| Scenario 3 | Lightning density U(0,3), lightning-caused fire efficiency U(0.03, 0.05) | High lightning susceptibility, Medium flammability; |
| Scenario 4 | Lightning density U(0,2), lightning-caused fire efficiency U(0.01, 0.03) | Medium lightning susceptibility, Low flammability; |
| Scenario 5 | Lightning density U(0,2), lightning-caused fire efficiency U(0.05, 0.07) | Medium lightning susceptibility, High flammability; |
| Scenario 6 | Lightning density U(0,3), lightning-caused fire efficiency U(0.05, 0.07) | Medium lightning susceptibility, High flammability; |
| Contants | Values | Contants | Values |
|---|---|---|---|
| 1.25 | |||
| 0.75 | 0.5 | ||
| 0.7 | 0.4 | ||
| 1 | 1000 | ||
| 0.7 | 1000 | ||
| 0.4 |
| Classification | Species | Quantitative Population |
|---|---|---|
| Class A | 1, 2, 3 | 87.6% |
| Class B | 4, 5, 6 | 11% |
| Class C | 7, 8, 9, 10 | 1.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, Z.; Wang, S.; Du, N. Simulation Study of an Abstract Forest Ecosystem with Multi-Species under Lightning-Caused Fires. Fire 2023, 6, 308. https://doi.org/10.3390/fire6080308
Ouyang Z, Wang S, Du N. Simulation Study of an Abstract Forest Ecosystem with Multi-Species under Lightning-Caused Fires. Fire. 2023; 6(8):308. https://doi.org/10.3390/fire6080308
Chicago/Turabian StyleOuyang, Zhi, Shiying Wang, and Nisuo Du. 2023. "Simulation Study of an Abstract Forest Ecosystem with Multi-Species under Lightning-Caused Fires" Fire 6, no. 8: 308. https://doi.org/10.3390/fire6080308
APA StyleOuyang, Z., Wang, S., & Du, N. (2023). Simulation Study of an Abstract Forest Ecosystem with Multi-Species under Lightning-Caused Fires. Fire, 6(8), 308. https://doi.org/10.3390/fire6080308