Cultured Microfungal Communities in Biological Soil Crusts and Bare Soils at the Tabernas Desert, Spain

Abstract
:1. Introduction
2. Material and Methods
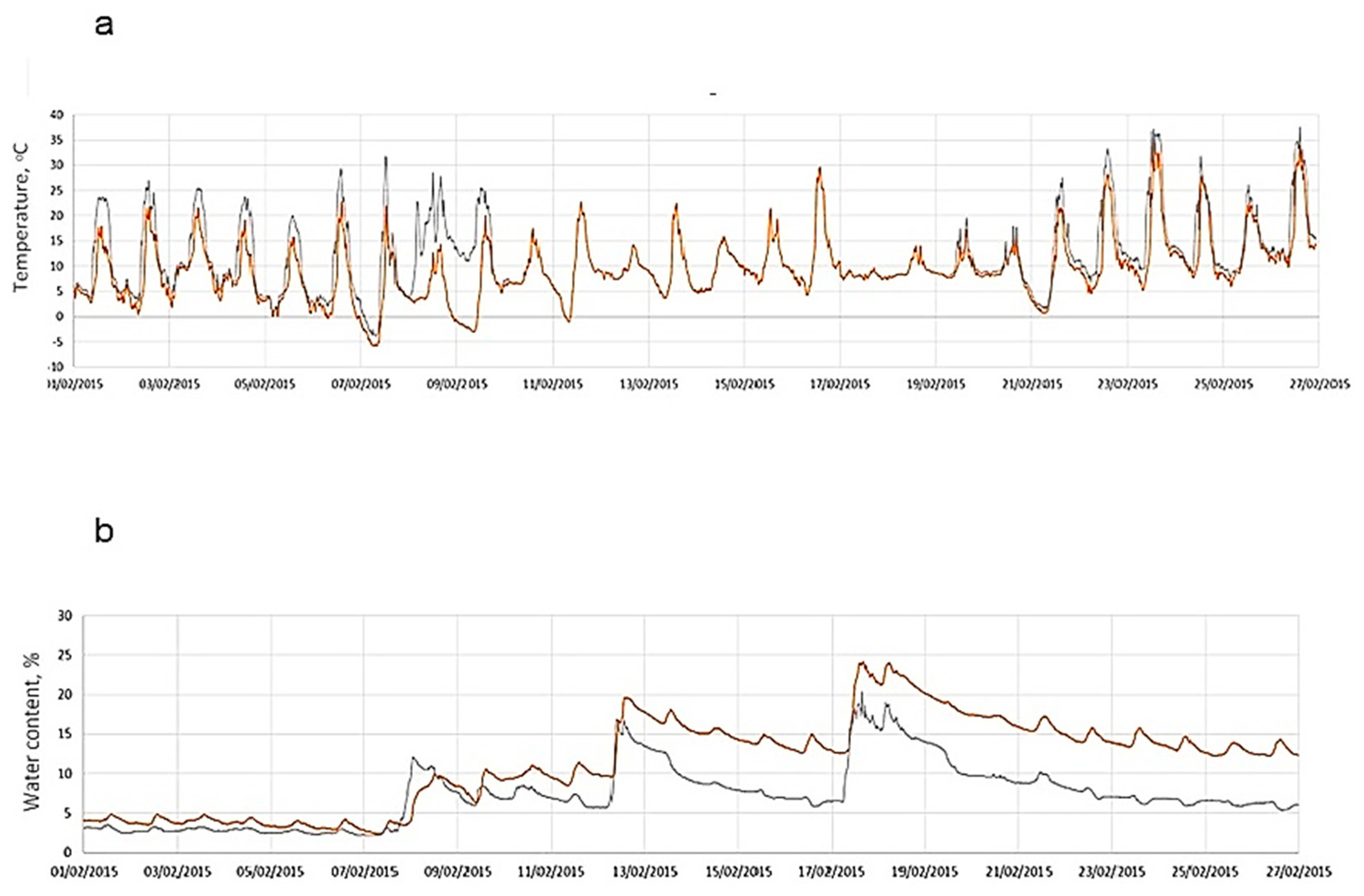
2.1. Site Description
2.2. Sampling
2.3. Characterization of Fungal Communities
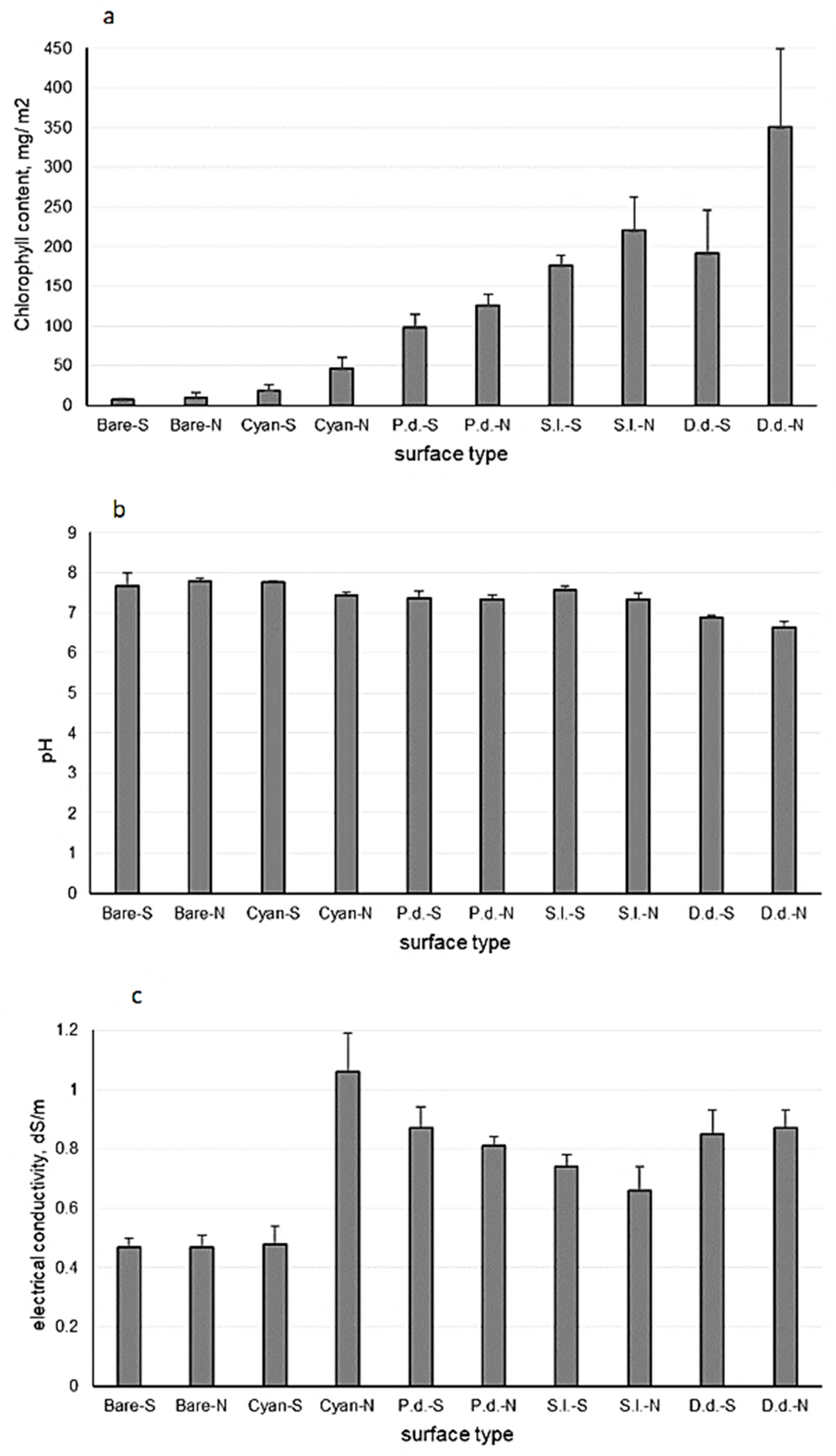
2.4. Measurement of Chlorophyll Content, pH, and Electrical Conductivity (EC)
2.5. Data Analyses
3. Results
3.1. Edaphic Characteristics
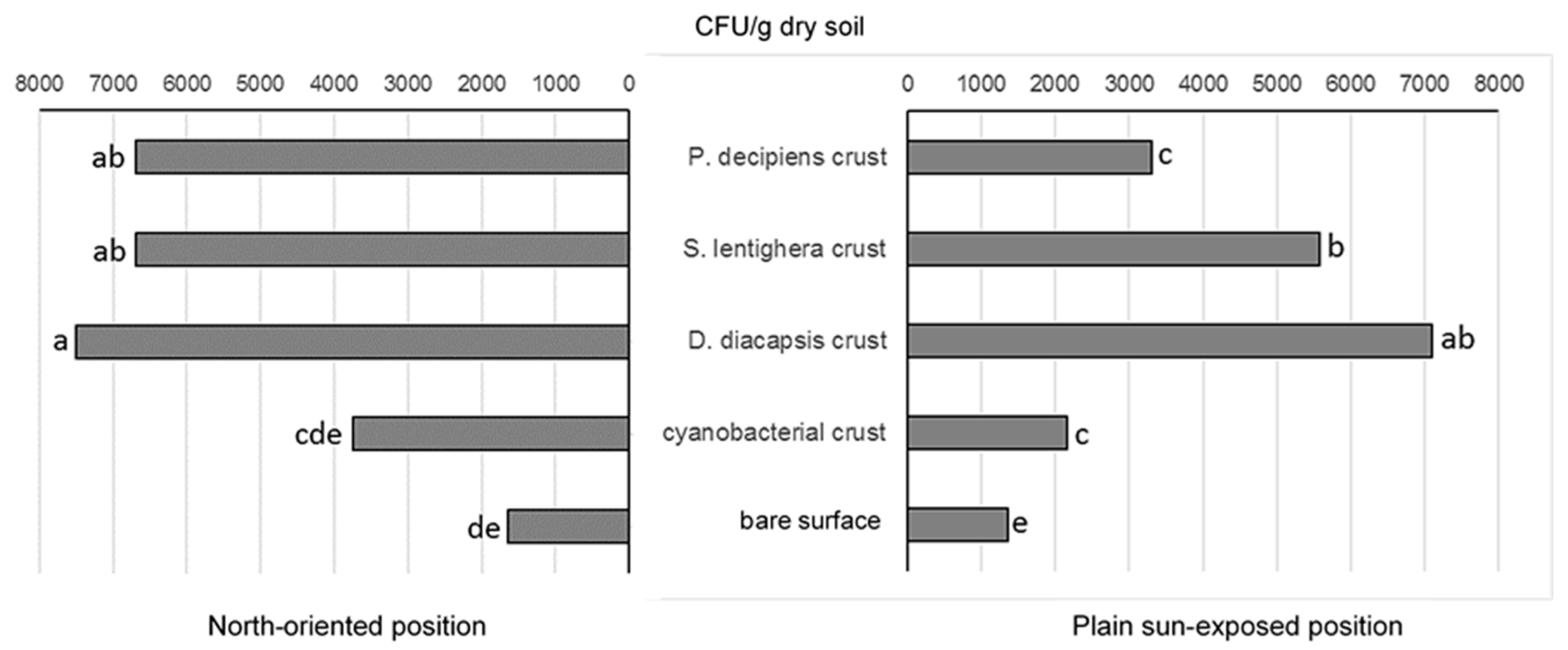
3.2. Density of Microfungal Isolates
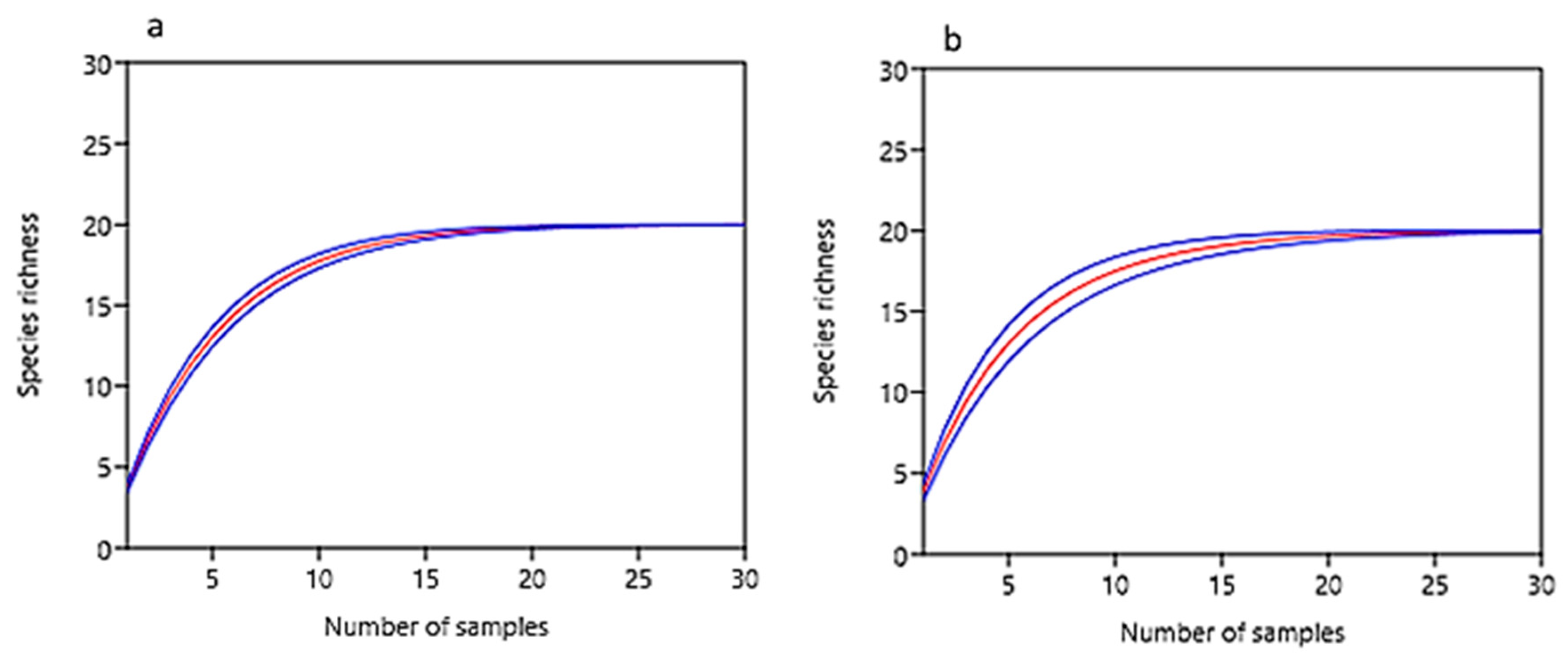
3.3. Composition and Diversity of Microfungal Communities
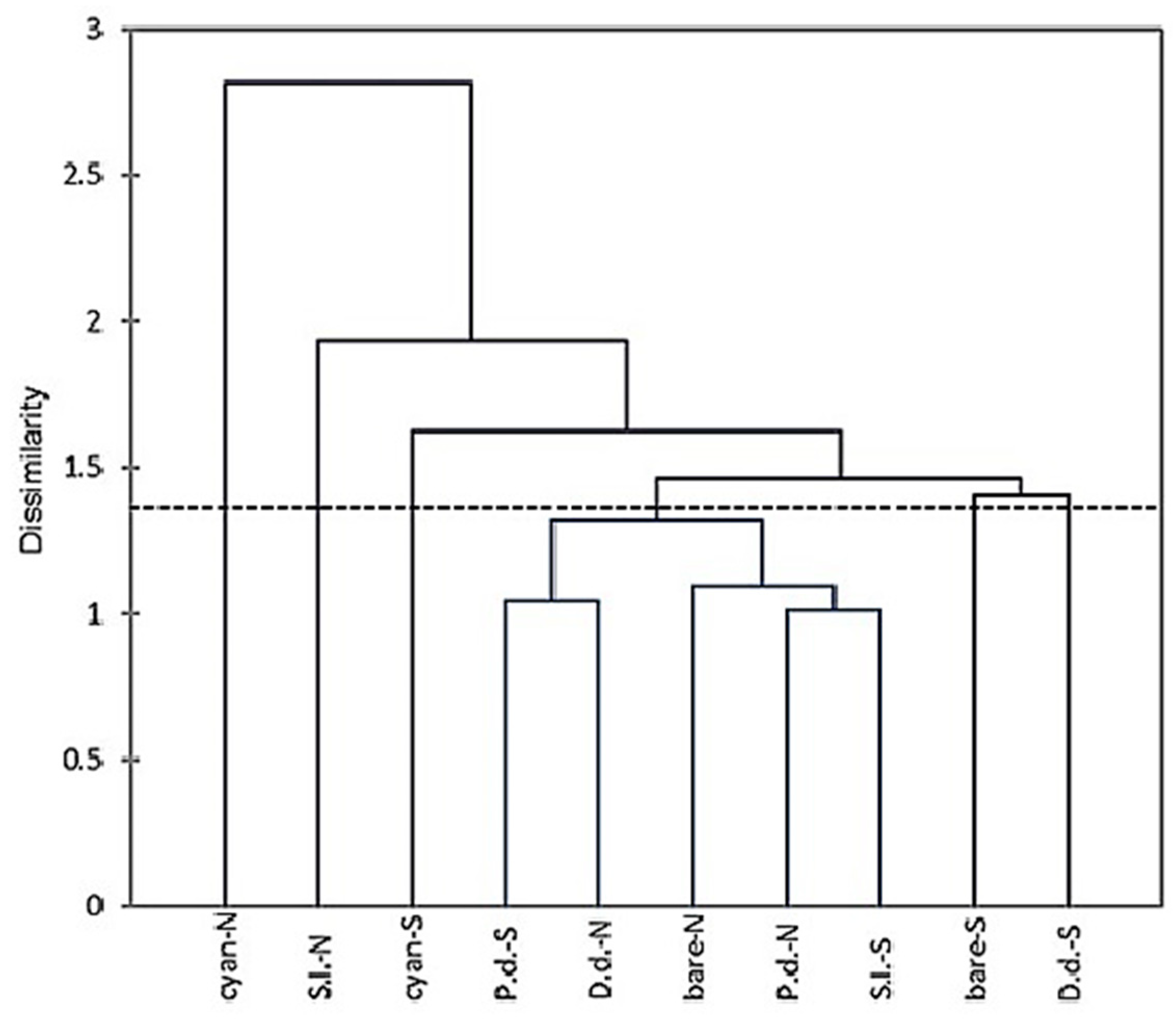
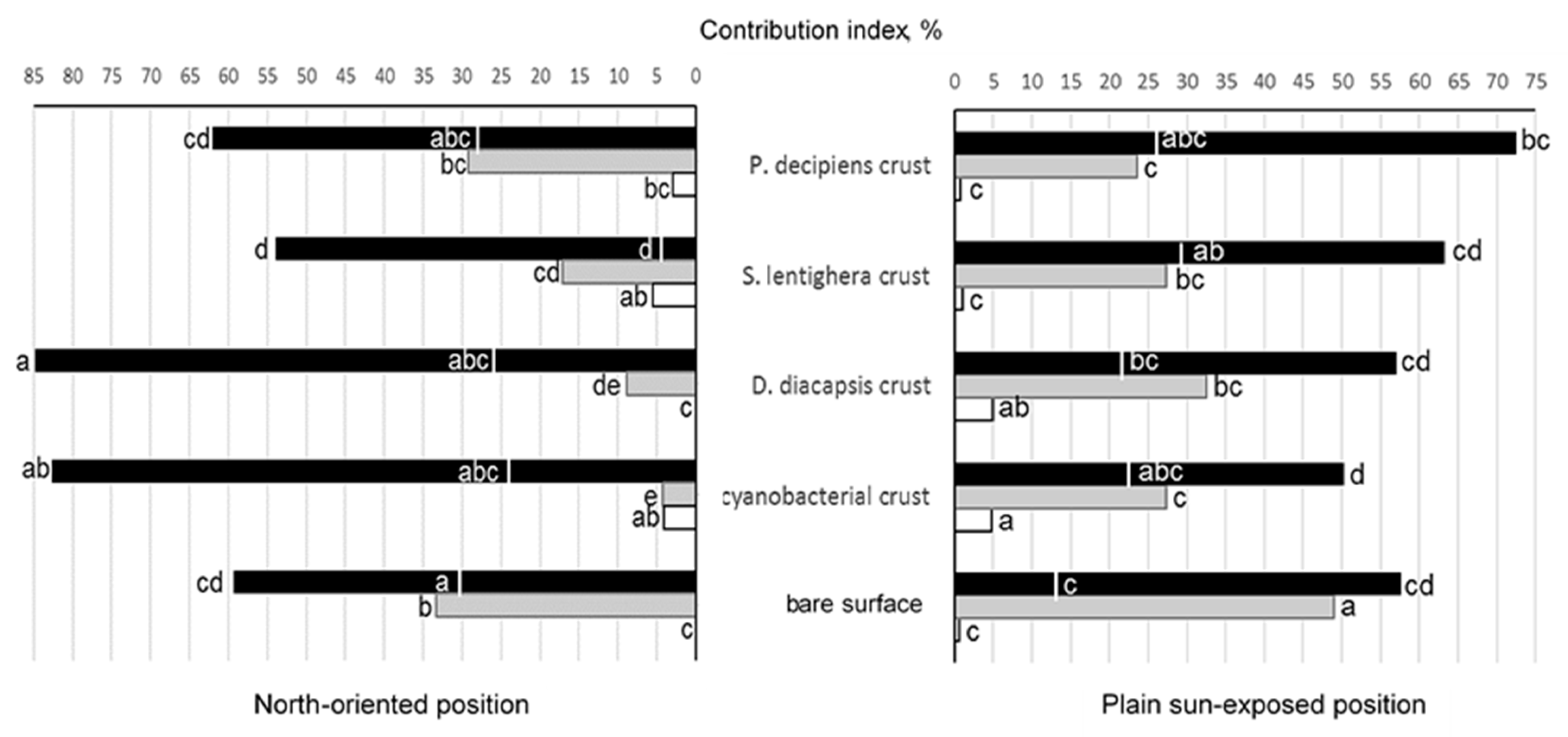
3.4. Effect of Surface Type and Orientation on Characteristics of Microfungal Communities
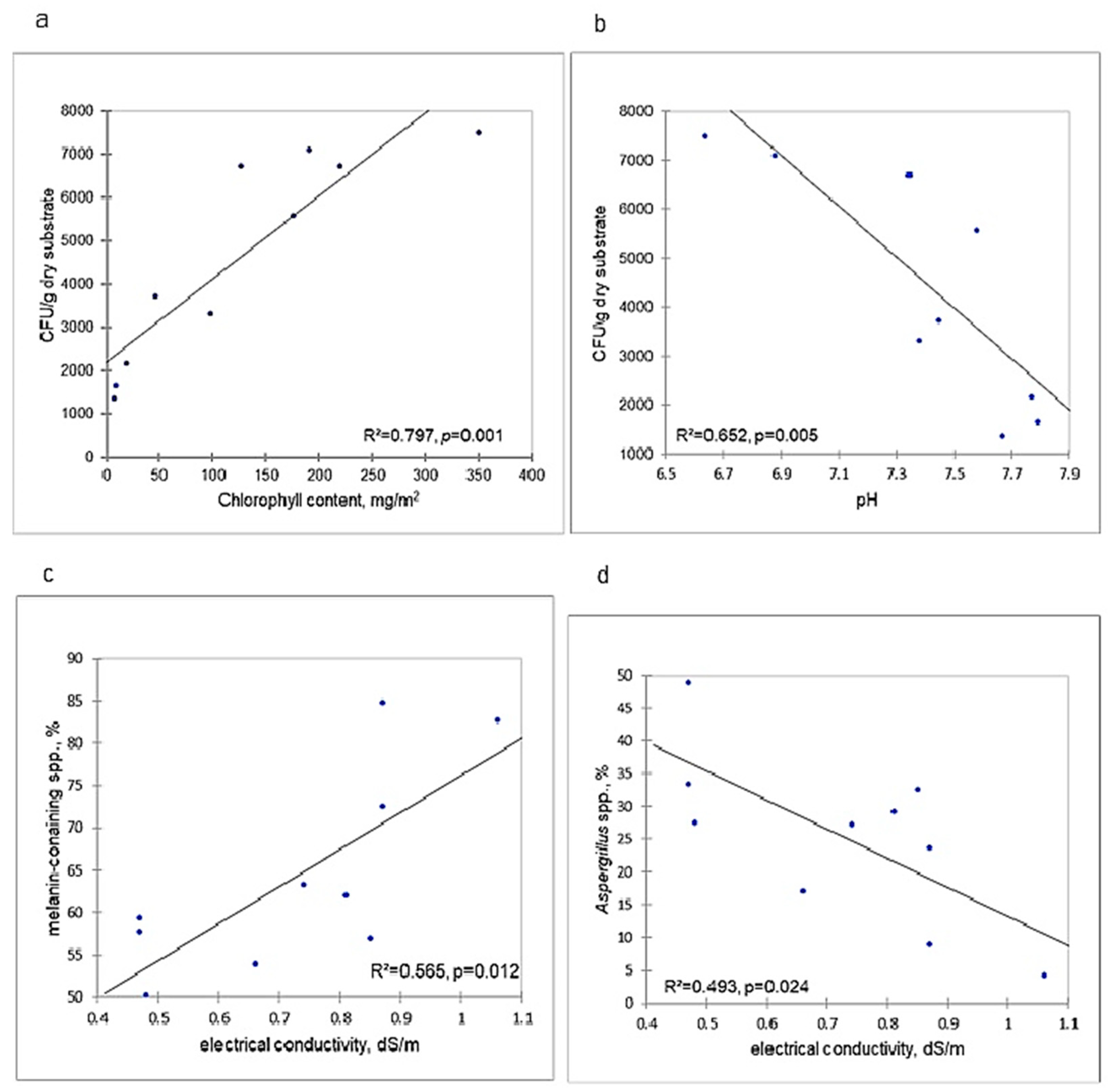
3.5. Relationships of Mycobiotic Characteristics with Edaphic Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belnap, J.; Lange, O. Preface. In Biological Soil Crusts: Structure, Function, and Management; Belnap, J., Lange, O., Eds.; Springer: New York, NY, USA, 2001; pp. V–IX. [Google Scholar]
- Kidron, G.J. Differential water distribution over dune slopes as affected by slope position and microbiotic crust, Negev Desert, Israel. Hydrol. Process. 1999, 13, 1665–1682. [Google Scholar] [CrossRef]
- Kidron, G.J.; Yair, A. Rainfall-runoff relationship over encrusted dune surfaces, Nizzana, western Negev, Israel. Earth Surf. Process. Landf. 1997, 22, 1169–1184. [Google Scholar] [CrossRef]
- Warren, S.D. Ecological role of microphytic soil crusts in arid environments. In Microbial Diversity and Function; Allsopp, D., Caldwell, R.R., Hawksworth, D.L., Eds.; CAB International: Wallingford, UK, 1995; pp. 199–209. [Google Scholar]
- Lázaro, R.; Cantón, Y.; Solé-Benet, A.; Bevan, J.; Alexander, R.; Sancho, L.G.; Puigdefábregas, J. The influence of competition between lichen colonization and erosion on the evolution of soil surfaces in the badlands (SE Spain) and its landscape effects. Geomorphology 2008, 102, 252–266. [Google Scholar] [CrossRef]
- Büdel, B.; Darienko, T.; Deutschewitz, K.; Dojani, S.; Friedl, T.; Mohr, K.I.; Salisch, M.; Reisser, W.; Weber, B. Southern African biological soil crusts are ubiquitous and highly diverse in drylands, being restricted by rainfall frequency. Microb. Ecol. 2009, 57, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Kidron, G.J.; Barinova, S.; Vonchak, A. The effects of heavy winter rains and rare summer rains on biological soil crusts in the Negev Desert. Catena 2012, 95, 6–11. [Google Scholar] [CrossRef]
- Kidron, G.J.; Vonshak, A.; Abeliovich, A. Microbiotic crusts as biomarkers for surface stability and wetness duration in the Negev Desert. Earth Surf. Proc. Land. 2009, 34, 1594–1604. [Google Scholar] [CrossRef]
- States, J.S.; Christensen, M. Fungi associated with biological soil crusts in desert grasslands of Utah and Wyoming. Mycologia 2001, 93, 432–439. [Google Scholar] [CrossRef] [Green Version]
- States, J.S.; Christensen, M.; Kinter, C.K. Soil fungi as components of biological soil crusts. In Biological Soil Crusts: Structure, Function, and Management; Belnap, J., Lange, O., Eds.; Springer: Berlin, Germany; New York, NY, USA, 2001; pp. 155–166. [Google Scholar]
- Bates, S.T.; Garcia-Pichel, F. A culture independent study of free-living fungi in biological soil crusts of the Colorado Plateau: their diversity and relative contribution to microbial biomass. Environ. Microbiol. 2009, 11, 56–67. [Google Scholar] [CrossRef]
- Bates, S.T.; Nash, T.H.; Sweat, K.G.; Garcia-Pichel, F. Fungal communities of lichen-dominated biological soil crusts: diversity, relative microbial biomass, and their relationship to disturbance and crust cover. J. Arid Environ. 2010, 74, 1192–1199. [Google Scholar] [CrossRef]
- Bates, S.T.; Nash, T.H., III; Garcia-Pichel, F. Patterns of diversity for fungal assemblages of biological soil crusts from the southwestern United States. Mycologia 2012, 104, 353–361. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Herrera, J.; Navig, D.O.; Lipinski, K.; Sinsabaugh, R.L. Diversity and distribution of soil fungal communities in a semiarid grassland. Mycologia 2011, 103, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Jia, R.-L.; Yu, L.-Y. Diversity and distribution of soil fungal communities associated with biological soil crusts in the southeastern Tengger Desert (China) as revealed by 454 pyrosequencing. Fung. Ecol. 2016, 23, 156–163. [Google Scholar] [CrossRef]
- Abed, R.M.M.; Al-Sadi, A.M.; Al-Shehi, M.; Al-Hinai, S.; Robinson, M.D. Diversity of free-living and lichenized fungal communities in biological soil crusts of the Sultanate of Oman and their role in improving soil properties. Soil Biol. Biochem. 2013, 57, 695–705. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Li, X.; Zhang, Y. Successional changes of fungal communities along the biocrust development stages. Biol. Fertil. Soils 2018, 54, 285–294. [Google Scholar] [CrossRef]
- Grishkan, I.; Zaady, E.; Nevo, E. Soil crust microfungi along a southward rainfall aridity gradient in the Negev Desert, Israel. Eur. J. Soil Biol. 2006, 42, 33–42. [Google Scholar] [CrossRef]
- Grishkan, I.; Kidron, G.J. Biocrust-inhabiting cultured microfungi along a dune catena in the western Negev Desert, Israel. Eur. J. Soil Biol. 2013, 56, 107–114. [Google Scholar] [CrossRef]
- Kidron, G.J.; Vonshak, A.; Dor, I.; Barinova, S.; Abeliovich, A. Properties and spatial distribution of microbiotic crusts in the Negev Desert, Israel. Catena 2010, 82, 92–101. [Google Scholar] [CrossRef]
- Kidron, G.J.; Li, X.R.; Jia, R.L.; Gao, Y.H.; Zhang, R. Assessment of carbon gains from biocrusts inhabiting a dunefield in the Negev Desert. Geoderma 2015, 253–254, 102–110. [Google Scholar] [CrossRef]
- Grishkan, I.; Rong-Liang, J.I.A.; Kidron, G.J.; Xin-Rong, L. Cultivable microfungal communities inhabiting biological soil crusts in the Tengger Desert, China. Pedosphere 2015, 25, 351–363. [Google Scholar] [CrossRef]
- Cantón, Y.; Solé-Benet, A.; Lázaro, R. Soil-geomorphology relations in gypsiferous materials of the Tabernas Desert (Almería, SE Spain). Geoderma 2003, 115, 193–222. [Google Scholar] [CrossRef]
- Lázaro, R.; Alexander, R.W.; Puigdefabregas, J. Cover distribution patterns of lichens, annuals and shrubs in the Tabernas Desert, Almería, Spain. In Vegetation Mapping: from Patch to Planet; Alexander, R.W., Millington, A.C., Eds.; Wiley: Chichester, UK, 2000; pp. 19–40. [Google Scholar]
- Bevan, J.L. Dynamics of Lichen Dominated Biological Soil Crusts in the El Cautivo Badlands, Southeast Spain. Ph.D. Thesis, University of Liverpool, Liverpool, UK, 2009. [Google Scholar]
- Alexander, R.W.; Calvo-Cases, A.; Arnau-Rosalén, E.; Mather, A.E.; Lázaro-Suau, R. Erosion and stabilization sequences in relation to base level changes in the El Cautivo badlands, SE Spain. Geomorphology 2008, 100, 83–90. [Google Scholar] [CrossRef]
- Lázaro, R.; Rodrigo, F.S.; Gutiérrez, L.; Domingo, F.; Puigdefábregas, J. Analysis of a thirty-year rainfall record (1967–1997) from semi-arid SE Spain for implications on vegetation. J. Arid Environ. 2001, 48, 373–395. [Google Scholar] [CrossRef]
- Lazaro, R.; Rodriguez-Tamayo, M.L.; Ordiales, R. El Clima. In Subdesiertos de Aimeria: Naturaleza de Cine; Pandalai, S.G., Mota, J., Cabello, J., Cerrillo, M.I., Rodriguez Tamayo, M.L., Eds.; Consejeria de Medio: Seville, Spain, 2004; pp. 63–79. [Google Scholar]
- Cantón, Y.; Del Barrio, G.; Solé-Benet, A.; Lázaro, L. Topographic controls on the spatial distribution of ground cover in a semiarid badlands area. Catena 2004, 55, 341–365. [Google Scholar] [CrossRef]
- Miralles, I.; Domingo, F.; Cantón, Y.; Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolase enzyme activities in a successional gradient of biological soil crusts in arid and semi-arid zones. Soil Biol. Biochem. 2012, 53, 124–132. [Google Scholar] [CrossRef]
- Lázaro, R.; Calvo-Cases, A.; Lázaro, A.; Molina, I. Effective run-off flow length over biological soil crusts on silty loam soils in drylands. Hydrol. Process. 2015, 29, 2534–2544. [Google Scholar] [CrossRef]
- Davet, P.; Rouxel, F. Detection and Isolation of Soil Fungi; Science Publisher Inc.: Enfield, UK; Plymouth, UK, 2000. [Google Scholar]
- Jeewon, R.; Hyde, K.D. Detection and diversity of fungi from environmental samples: traditional versus molecular approaches. In Soil Biology. Vol. 11. Advanced Techniques in Soil Microbiology; Varma, A., Oelmuller, R., Eds.; Springler: Berlin/Heidelberg, Germany, 2007; pp. 1–15. [Google Scholar]
- Bills, G.F.; Christensen, M.; Powell, M.; Thorn, G. Saprobic soil fungi. In Biodiversity of Fungi. Inventory and Monitoring Methods; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 271–302. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd revised ed.; Academic Press: New York, NY, USA, 2007. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; CAB International: Wallingford, UK, 1971. [Google Scholar]
- Klick, M.A. Identification of Common Aspegillus Species; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002. [Google Scholar]
- Graham, G.C.; Mayers, P.; Henry, R.J. A simplified method for the preparation of fungal genomic DNA for PCR and RAPD analysis. Biotechniques 1994, 16, 48–49. [Google Scholar] [PubMed]
- Altschul, S.F.; Madden, T.J.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.G.; Westlake, D.F. Periphyton. In A Manual on Methods for Measuring Primary Production in Aquatic Environments; Vollenweider, R.A., Ed.; Blackwell: Oxford, UK, 1969; pp. 33–40. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Addison Wesley Longman: Redwood City, CA, USA, 1999. [Google Scholar]
- Ranzoni, F.V. Fungi isolated in culture from soils of the Sonoran Desert. Mycologia 1968, 60, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M. Species diversity and dominance in fungal community. In The Fungal Community, Its Organization and Role in the Ecosystem; Carroll, G.W., Wicklow, D.T., Eds.; Marcell Dekker: New York, NY, USA, 1981; pp. 201–232. [Google Scholar]
- Halwagy, R.; Moustafa, A.F.; Kamel, S. Ecology of the soil mycoflora in the desert of Kuwait. J. Arid Environ. 1982, 5, 109–125. [Google Scholar] [CrossRef]
- Abdullah, S.K.; Al-Khesraji, T.O.; Al-Edany, T.Y. Soil mycoflora of the Southern Desert of Iraq. Sydowia 1986, 39, 8–16. [Google Scholar]
- Ciccarone, C.; Rambelli, A. A study on micro-fungi in arid areas. Notes on stress-tolerant fungi. Plant Biosyst. 1998, 132, 17–20. [Google Scholar] [CrossRef]
- Skujins, J. Microbial ecology of desert soils. Adv. Microb. Ecol. 1984, 7, 49–91. [Google Scholar]
- Zak, J. Fungal communities of desert ecosystems: links to climate change. In The Fungal Community, Its Organization and Role in the Ecosystem; Dighton, J., White, J.F., Jr., Oudemans, P., Eds.; CRC Press: London, UK, 2005; pp. 659–681. [Google Scholar]
- Eigenberg, R.A.; Doran, J.W.; Nienaber, J.A.; Ferguson, R.B.; Woodbury, B.L. Electrical conductivity monitoring of soil condition and available N with animal manure and a cover crop. Agric. Ecosyst. Environ. 2002, 88, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Grishkan, I.; Beharav, A.; Kirzhner, V.; Nevo, E. Adaptive spatiotemporal distribution of soil microfungi in “Evolution Canyon” III, Nahal Shaharut, extreme Southern Negev desert, Israel. Biol. J. Linnean Soc. 2007, 90, 263–277. [Google Scholar] [CrossRef]
- Grishkan, I.; Nevo, E. Spatiotemporal distribution of soil microfungi in the Makhtesh Ramon area, central Negev desert, Israel. Fung. Ecol. 2010, 3, 326–337. [Google Scholar] [CrossRef]
- Kay, B.D. Soil structure and organic carbon: A review. In Soil Processes and the Carbon Cycle; Lal, R., Kimble, J.M., Follet, R.F., Stewart, R.A., Eds.; CRC Press: London, UK, 1997; pp. 169–197. [Google Scholar]
- Hibbett, D.; Taylor, J.W. Fungal systematics: Is a new age of enlightenment at hand. Nat. Rev. Microbiol. 2013, 11, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Peay, K.G. Back to the future: natural history and the way forward in modern fungal ecology. Fung. Ecol. 2014, 12, 4–9. [Google Scholar] [CrossRef]






 —melanin-containing microfungi;
—melanin-containing microfungi;  —Aspergillus spp.;
—Aspergillus spp.;  —Penicillium spp.; the area left (plain sun-exposed position) or right (north-oriented position) from the line on the bars of melanin-containing microfungi indicates contributions of species with large multicellular spores. Means with the same letters are not significantly different (a one-way ANOVA, the Fisher test, at the 5% level).
—melanin-containing microfungi; —Aspergillus spp.; —Penicillium spp.; the area left (plain sun-exposed position) or right (north-oriented position) from the line on the bars of melanin-containing microfungi indicates contributions of species with large multicellular spores. Means with the same letters are not significantly different (a one-way ANOVA, the Fisher test, at the 5% level).
—Penicillium spp.; the area left (plain sun-exposed position) or right (north-oriented position) from the line on the bars of melanin-containing microfungi indicates contributions of species with large multicellular spores. Means with the same letters are not significantly different (a one-way ANOVA, the Fisher test, at the 5% level).
—melanin-containing microfungi; —Aspergillus spp.; —Penicillium spp.; the area left (plain sun-exposed position) or right (north-oriented position) from the line on the bars of melanin-containing microfungi indicates contributions of species with large multicellular spores. Means with the same letters are not significantly different (a one-way ANOVA, the Fisher test, at the 5% level).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | North-Oriented Positon | Plain Sun-Exposed Position | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bare | Cyan (a) | P.d. | S.l. | D.d. | Bare | Cyan | P.d. | S.l. | D.d. | |
| Zygomycota | ||||||||||
| Absidia corymbifera | - | - | - | - | 0.3 | - | - | - | - | - |
| Actinomucor elegans | - | - | - | - | - | - | - | - | - | 0.1 |
| Mortierella humilis | 0.5 | - | - | - | - | - | - | - | - | - |
| Mucor hiemalis | - | - | - | - | - | - | - | - | 0.15 | - |
| M. plumbeus | - | - | - | - | - | - | - | - | - | 0.1 |
| Rhizopus arrhizus | 0.5 | 0.5 | 2 | - | - | 0.9 | 0.6 | 0.75 | 0.45 | 1.4 |
| teleomorphic Ascomycota | ||||||||||
| Aspergillus nidulans | 0.5 | - | - | - | - | - | 0.3 | - | 1 | - |
| A. rugulosus | - | - | - | 0.12 | - | - | - | - | - | - |
| Canariomyces notabilis | 0.5 | - | - | - | - | - | - | - | - | - |
| Chaetomidium subfimeti | - | - | - | 0.12 | 0.1 | - | - | - | - | - |
| Chaetomium strumarium | - | - | - | - | 0.4 | - | - | - | - | - |
| Ch. succineum | - | 0.35 | 0.2 | - | - | 0.3 | 4.5 | - | - | - |
| Chaetomium sp. | 1.5 | - | 0.45 | 0.12 | 0.1 | 0.6 | 1.2 | - | 0.15 | - |
| Chaetomium sp.1 | - | 0.17 | 0.2 | - | - | - | 0.3 | 0.6 | - | - |
| Sordaria fimicola | - | - | - | - | 1.1 | - | - | 0.2 | - | - |
| Sporomiella minima | 1.5 | 0.7 | - | - | - | - | 0.6 | - | - | - |
| Immature fruit bodies | - | - | - | - | - | - | 0.3 | 0.2 | - | - |
| anamorphic Ascomycota | ||||||||||
| Acremonium charticola | - | - | 0.2 | - | - | - | - | - | - | - |
| Alternaria alternata | 2.1 | 0.5 | 0.35 | 0.25 | 0.1 | 5.2 | 0.3 | 0.6 | 0.6 | 0.9 |
| A. atra | 0.5 | 2.2 | 0.2 | 1.2 | 2.0 | 1.5 | 1.2 | 3.4 | 1.5 | 1.4 |
| A. phragmospora | 29.5 | 7.4 | 21.9 | 1.3 | 20.9 | 3.1 | 20.2 | 23.2 | 33.2 | 6.5 |
| A. raphani | - | - | - | - | - | 0.3 | 0.9 | - | - | 0.1 |
| Amerosporium concinnum | - | - | - | - | - | - | - | - | 0.15 | - |
| Aphanocladium album | - | 1.3 | 0 | 0.25 | - | - | 0.6 | 0.4 | - | - |
| Aspergillus alliaceus | - | - | - | - | - | - | - | - | 0.3 | - |
| A. flavus | - | 0.5 | - | 0.12 | - | - | - | 0.2 | - | 0.1 |
| A. fumigatus | 42.4 | 2.2 | 48 | 13.4 | 6.4 | 58.7 | 37.5 | 22.8 | 29.5 | 43.6 |
| A. niger | - | - | 0.35 | 0.25 | - | 7.4 | - | - | 0.45 | 0.5 |
| A. niveus | - | - | - | 0.12 | - | - | - | - | 0.3 | - |
| A. puniceus | - | - | - | - | - | - | - | 0.2 | - | - |
| A. terreus | - | - | - | - | - | - | - | - | - | 0.1 |
| A. versicolor | 0.5 | - | - | - | - | - | - | - | - | - |
| Aspergillus sp. | - | - | - | - | - | - | - | - | 0.3 | - |
| Aureobasidium pullulans | - | - | - | - | 0.1 | - | - | - | - | - |
| Beauveria bassiana | - | - | - | 0.25 | - | - | - | - | - | - |
| Boeremia exigua | 7.3 | 5.9 | 14 | 46.6 | 48.3 | 7.1 | 3.9 | 23.2 | 8.2 | 11.4 |
| Camarosporium aequivocum | 2.6 | - | - | - | - | 0.3 | 0.3 | 2.6 | - | - |
| Cladosporium cladosporioides | - | - | - | - | - | 0.6 | - | - | - | - |
| Coleophoma empetri | 0.5 | - | 6.3 | 4.6 | 13.4 | 10.4 | 0.6 | 14.5 | 7 | 16.1 |
| Curvularia inaequales | - | 6.4 | 1.6 | - | - | - | - | - | - | 0.9 |
| Engyodontium album | - | 0.8 | - | - | - | - | - | - | - | - |
| Epicoccum nigrum | 0.5 | - | - | - | - | 0.3 | - | - | - | - |
| Fusarium gibbosum | - | 1.3 | - | 4.1 | 1.7 | 0.3 | 2.1 | 1.3 | 0.45 | 1.4 |
| F. oxysporum | 4.1 | 0.35 | - | 0.5 | - | 0.9 | 8.4 | - | - | - |
| F. solani | 0.5 | |||||||||
| Lecanicillium psaliotae | - | - | - | - | - | - | - | 0.6 | - | - |
| Malbranchea pulchella | 0.5 | - | - | - | - | - | - | - | - | - |
| Metarhizium marquandii | - | - | - | - | - | - | - | 0.2 | 0.6 | - |
| Neoscytalidium dimidiatum | - | 0.17 | - | - | - | - | - | - | - | - |
| Papulaspora pannosa | 1.0 | - | - | 0.4 | 0.6 | - | - | - | - | - |
| Paraboeremia putaminum | - | 53.9 | - | 0.25 | - | - | 1.2 | - | 1 | - |
| Penicillium aurantiogriseum | - | 2.9 | 0.2 | 2 | - | - | 0.9 | - | 0.15 | - |
| P. brevicompactum | - | - | - | - | - | - | 0.6 | - | - | 0.1 |
| P. glabrum | - | - | - | - | - | - | - | - | - | 0.9 |
| P. griseoroseum | - | - | - | - | - | - | - | - | - | 1.7 |
| P. corylophylum | - | - | - | - | - | - | - | 0.2 | - | - |
| P. herquei | - | - | - | - | - | - | - | - | - | 0.2 |
| P. janczewskii | - | - | - | - | - | - | 0.3 | - | - | - |
| P. lividum | - | - | - | 0.12 | - | - | - | - | - | - |
| P. simplicissimum | - | - | 0.8 | - | - | 0.3 | 0.9 | - | 0.45 | - |
| P. waksmani | - | - | - | - | - | - | 0.3 | - | - | - |
| Phoma medicaginis | - | - | - | 0.25 | - | - | - | - | - | 0.2 |
| Phomatodes nebulosa | - | - | 0.35 | - | - | - | - | - | - | - |
| Pithomyces africanus | - | 1.2 | - | - | - | - | - | - | - | - |
| Plenodomus tracheiphilus | - | - | - | 1.1 | - | - | - | - | - | - |
| Pleospora tarda | 0.5 | 0.35 | - | - | - | - | 0.3 | - | - | 6.9 |
| Pseudogymnoascus pannorum | - | 0.7 | - | - | - | - | - | - | - | - |
| Pyrenochaeta cava | 1.0 | 0.8 | - | 1.5 | 0.3 | - | 2.7 | 1.1 | 1.5 | - |
| Schizothecium inaequale(b) | 1.5 | 6.9 | 1.4 | 0.75 | - | - | 2.7 | 0.9 | 6 | 2.7 |
| Setophoma terrestris (b) | 0.5 | 1.0 | - | - | - | 0.9 | 1.5 | - | - | - |
| Stachybotrys chartarum | - | 0.17 | - | 1.1 | - | 0.6 | 0.3 | 0.75 | 0.75 | - |
| Talaromyces purpurogenus | - | - | - | 0.75 | - | - | - | - | - | 0.1 |
| T. variabilis | - | - | - | - | 0.1 | - | - | 0.2 | - | - |
| Trichoderma koningii | - | 0.8 | - | - | 0.4 | - | - | - | - | - |
| T. viride | - | - | - | 0.6 | - | - | - | - | - | - |
| Xepicula leucotricha | - | - | - | 16 | - | - | - | - | 4.6 | - |
| Basidiomycota | ||||||||||
| Disporotrichum dimorphosporum | - | - | - | - | - | - | 2.7 | - | - | - |
| Non-sporulating strains | ||||||||||
| light-colored | - | 0.5 | - | - | - | - | 1.8 | 0.6 | 0.6 | 0.9 |
| dark-colored | - | - | 1.5 | 1.7 | 3.2 | 0.3 | - | 1.3 | 0.6 | 1.7 |
| Surface Type | North-Oriented Position | Plain Sun-Exposed Position | ||||
|---|---|---|---|---|---|---|
| R | H | J | R | H | J | |
| P. decipiens crust | 19 c | 1.56 c | 0.53 ab | 24 ab | 2.02 ab | 0.63 a |
| S. lentighera crust | 29 a | 1.86 abc | 0.55 ab | 27 a | 2.0 ab | 0.61 a |
| D. diacapsis crust | 19 c | 1.53 c | 0.52 ab | 25 ab | 1.95 ab | 0.61 a |
| cyanobacterial crust | 27 ab | 1.92 ab | 0.58 a | 31 a | 2.27 a | 0.66 a |
| bare surface | 22 b | 1.79 bc | 0.59 a | 20 c | 1.59 c | 0.53 ab |
| Parameter | Surface Type | Orientation | Type × Orientation |
|---|---|---|---|
| Species richness | 3.17 @ | NS | 4.05 * |
| Shannon index | NS | NS | NS |
| Evenness | NS | NS | NS |
| Melanin-containing spp. | 2.98 @ | 4.29 @ | 3.52 @ |
| Melanics with multicellular spores | NS | NS | 4.38 * |
| Penicillium spp. | 3.95@ | NS | 3.44 @ |
| Aspergillus spp. | 7.71 *** | 8.96 ** | NS |
| Isolate density | 16.6 **** | 6.9 * | NS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grishkan, I.; Lázaro, R.; Kidron, G.J. Cultured Microfungal Communities in Biological Soil Crusts and Bare Soils at the Tabernas Desert, Spain. Soil Syst. 2019, 3, 36. https://doi.org/10.3390/soilsystems3020036
Grishkan I, Lázaro R, Kidron GJ. Cultured Microfungal Communities in Biological Soil Crusts and Bare Soils at the Tabernas Desert, Spain. Soil Systems. 2019; 3(2):36. https://doi.org/10.3390/soilsystems3020036
Chicago/Turabian StyleGrishkan, Isabella, Roberto Lázaro, and Giora J. Kidron. 2019. "Cultured Microfungal Communities in Biological Soil Crusts and Bare Soils at the Tabernas Desert, Spain" Soil Systems 3, no. 2: 36. https://doi.org/10.3390/soilsystems3020036
APA StyleGrishkan, I., Lázaro, R., & Kidron, G. J. (2019). Cultured Microfungal Communities in Biological Soil Crusts and Bare Soils at the Tabernas Desert, Spain. Soil Systems, 3(2), 36. https://doi.org/10.3390/soilsystems3020036