Molecular Scale Studies of Phosphorus Speciation and Transformation in Manure Amended and Microdose Fertilized Indigenous Vegetable Production Systems of Nigeria and Republic of Benin

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phosphorus K-Edge XAS
2.2. Statistical Analysis
3. Results
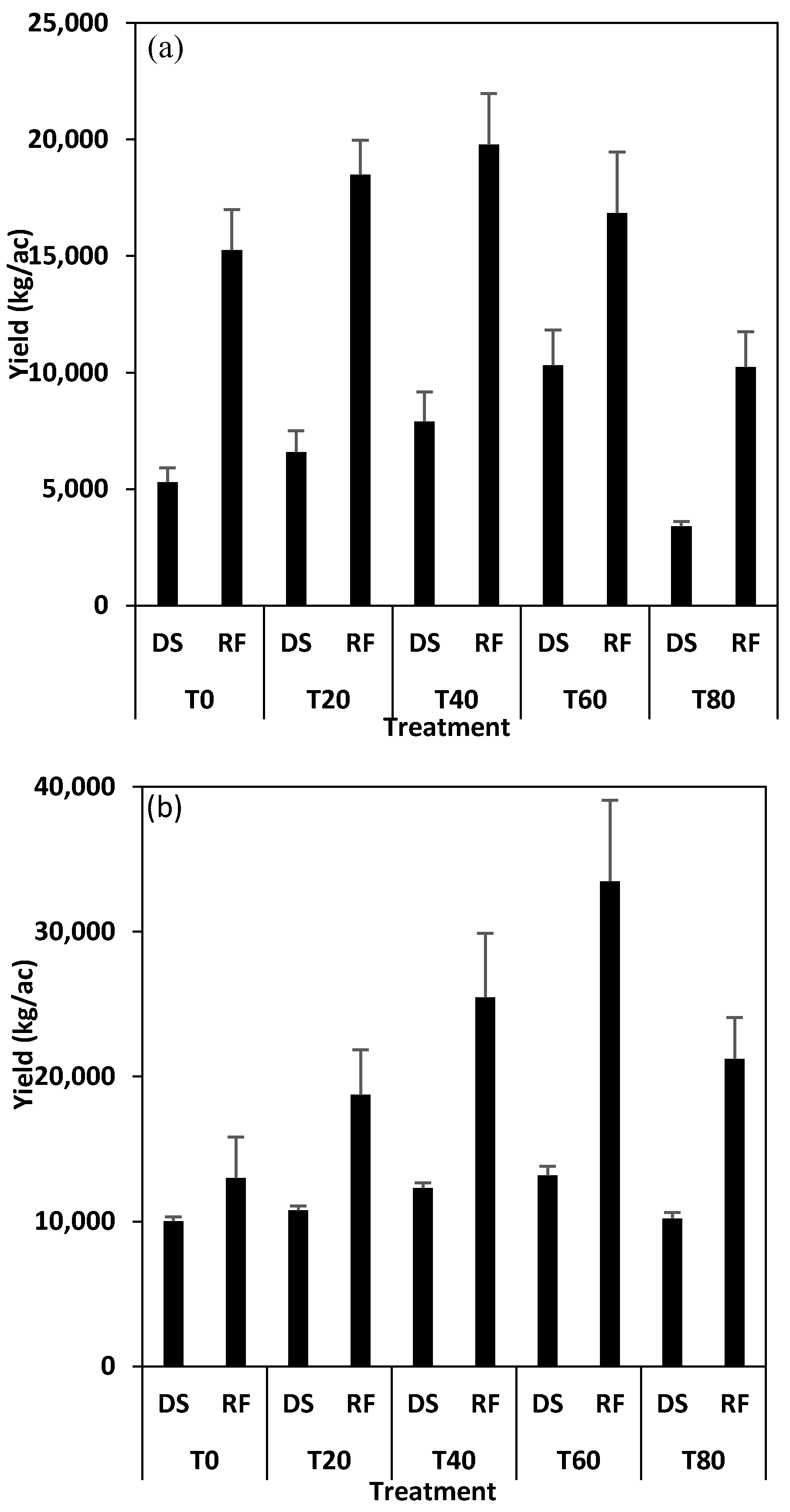
3.1. Yields
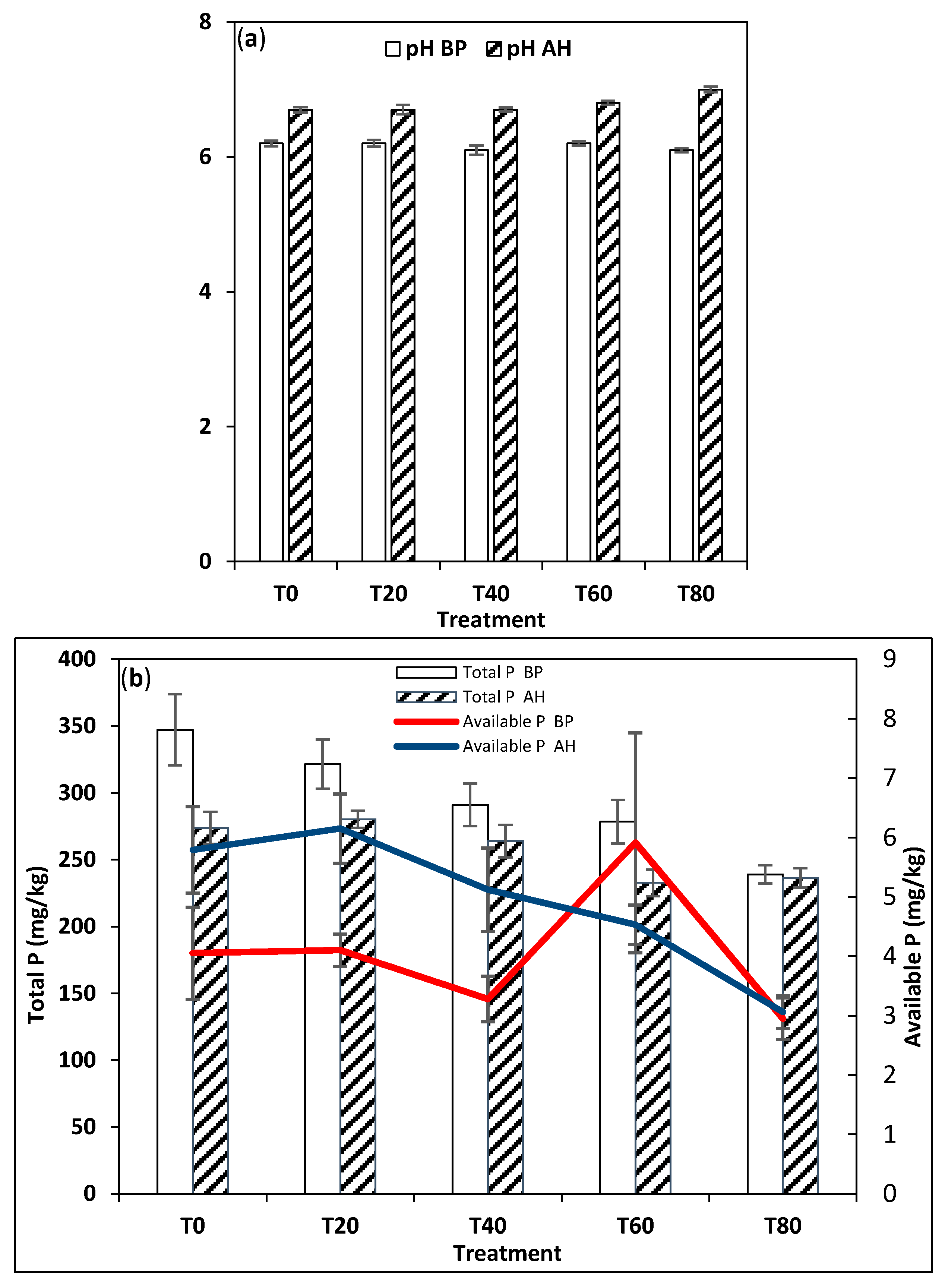
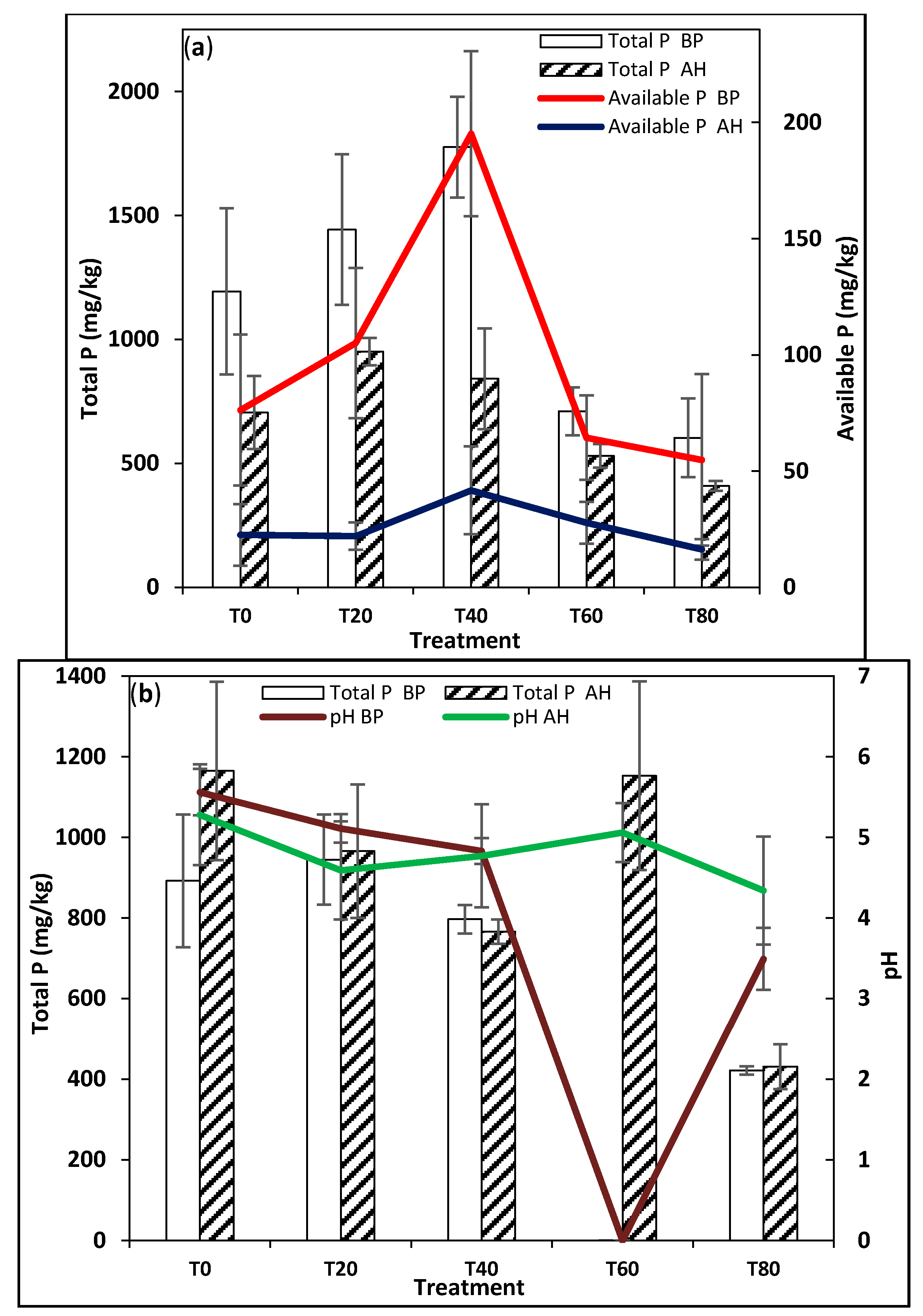
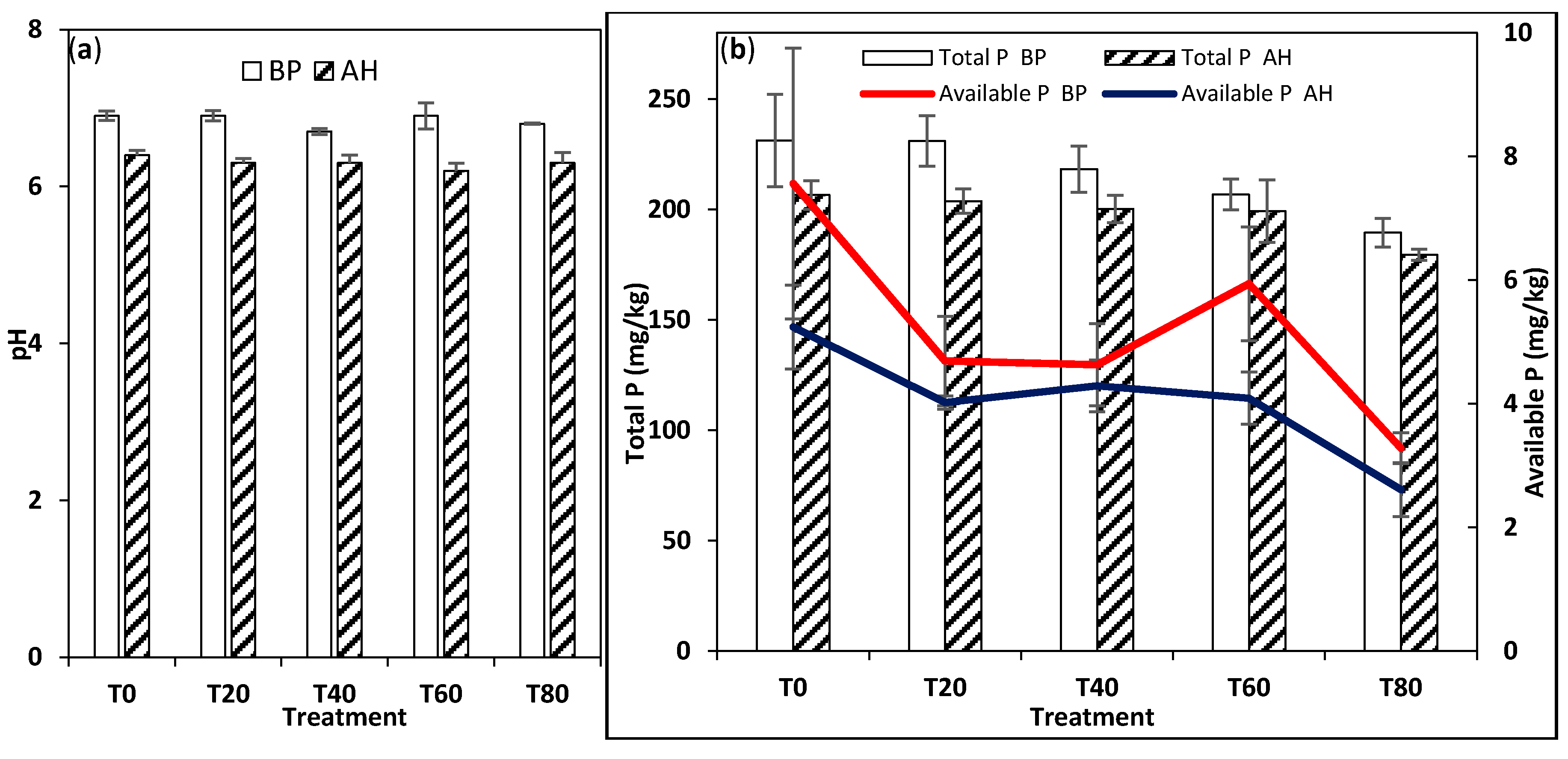
3.2. Soil Physico-Chemical Analysis
3.3. P XANES Analysis
3.4. X-Ray Fluorescence Imaging
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zingore, S.; Delve, R.J.; Nyamangara, J.; Giller, K.E. Multiple benefits of manure: The key to maintenance of soil fertility and restoration of depleted sandy soils on African smallholder farms. Nutr. Cycl. Agroecosyst. 2007, 80, 267–282. [Google Scholar] [CrossRef]
- Twomlow, S.; Mugabe, F.T.; Mwale, M.; Delve, R.; Nanja, D.; Carberry, P.; Howden, M. Building adaptive capacity to cope with increasing vulnerability due to climatic change in Africa- a new approach. Phys. Chem. Earth 2008, 33, 780–787. [Google Scholar] [CrossRef]
- United Nations (UN). World Population Prospects: The 2000 Revision; Department of Economic and Social Affairs: New York, NY, USA, 2001. [Google Scholar]
- Bationo, A.B.; Waswa, B.; Abdou, A.; Bado, B.V.; Bonzi, M.; Iwuafor, E.; Kibunja, C.; Kihara, J.; Mucheru, M.; Mugendi, D.; et al. Lessons Learned from Long-Term Soil Fertility Management Experiments in Africa; Springer: Dordrecht, The Netherlands, 2012; pp. 1–26. [Google Scholar]
- Callo-Concha, D.; Gaiser, T.; Webber, H.; Tischbein, B.; Muller, M.; Ewert, F. Farming in the West African Sudan Savanna: Insights in the context of climate change. Afr. J. Agric. Res. 2013, 8, 4693–4705. [Google Scholar] [CrossRef] [Green Version]
- Alliance for a Green Revolution in Africa (AGRA). Building on the New Momentum in African Agriculture: AGRA in 2008; Alliance for a Green Revolution in Africa: Nairobi, Kenya, 2009. [Google Scholar]
- Vanlauwe, B.; Kihara, J.; Chivenge, P.; Pypers, P.; Coe, R.; Six, J. Agronomic use efficiency of N fertilizer in maize-based systems in sub-Saharan Africa within the context of integrated soil fertility management. Plant Soil 2010, 339, 35–50. [Google Scholar] [CrossRef]
- International Crops Research Institute for the Semi-Arid Tropics (ICRISAT). Fertilizer Microdosing; ICRISAT Publ.: Patancheru, India, 2009; Available online: www.icrisat.org/fertilizermicrodosing (accessed on 3 May 2018).
- Adebooye, O.C. Proximate composition and nutrient analysis of six selected leaf vegetables of southwest Nigeria. IFE J. Agric. 1996, 18, 56–62. [Google Scholar]
- Adebooye, O.C. Wild Plants for Medicinal and Culinary Uses: Nigeria. Sharing Innovative Experiences; TWAS: Trieste, Italy, 2001; Volume 2, pp. 69–78. [Google Scholar]
- United States Department of Agriculture (USDA). USDA Nutrient Database for Standard Reference; Release 16; US. Department of Agriculture, Agriculture Research Service, Nutrient Data Laboratory: Beltsville, MD, USA, 2003. Available online: www.nal.usda.gov/fnic/foodcomp (accessed on 12 January 2016).
- Smith, I.F.; Eyzaguirre, P. African leafy vegetables: Their role in the world health organization’s global fruit and vegetables initiative. Afr. J. Food Agric. Nutr. 2007, 7, 3. [Google Scholar]
- Adams, A.M.; Gillespie, A.W.; Kar, G.; Koala, S.; Ouattara, B.; Kimaro, A.A.; Bationo, A.; Akponikpe, P.B.I.; Schoenau, J.J.; Peak, D. Long term effects of reduced fertilizer rates on millet yields and soil properties in the West-African Sahel. Nutr. Cycl. Agroecosyst. 2016, 106, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Hammond, J.P.; White, P.J. Sucrose transport in the phloem: Integrating root responses to phosphorus starvation. J. Exp. Bot. 2008, 59, 93–109. [Google Scholar] [CrossRef]
- Pierzynski, G.M.; Sims, J.T.; Vance, G.F. Soils and Environmental Quality, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Richardson, A.E. Utilization of Soil Organic Phosphorus by Higher Plants; Organic Phosphorus in the Environment; CABI Publishing: Oxfordshire, UK, 2005; pp. 165–184. [Google Scholar]
- Castillo, A.E.; Benito, S.G.; Fernandez, J.A. Using organic manures as liming materials. Agrochimica 2003, 47, 14–20. [Google Scholar]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Nitrogen. In Principles of Plant Nutrition; Springer: Dordrecht, The Netherland, 2001; pp. 397–434. [Google Scholar]
- Cooperband, L.; Bollero, G.; Coale, F. Effect of poultry litter and composts on soil nitrogen and phosphorus availability and corn production. Nutr. Cycl. Agroecosyst. 2002, 62, 185–194. [Google Scholar] [CrossRef]
- Kashem, M.A.; Akinremi, O.O.; Racz, G.J. Phosphorus fractions in soil amended with organic and inorganic phosphorus sources. Can. J. Soil Sci. 2004, 84, 83–90. [Google Scholar] [CrossRef]
- Montgomery, D.R. Dirt the Erosion of Civilizations; University of California Press: Berkeley, CA, USA, 2007. [Google Scholar]
- Schoenau, J.J.; Carley, C.; Stumborg, C.; Malhi, S.S. Strategies for maximizing crop recovery of applied manure nitrogen in the Northern Great Plains of North America. In Recent Trends in Soil Science and Agronomy Research in the Northern Great Plains of North America; Malhi, S.S., Gan, Y., Schoenau, J.J., Lemke, R.L., Mark, A., Eds.; Liebig. Research Signpost: Kerala, India, 2010; pp. 95–108. [Google Scholar]
- Lemke, R.L.; Farrell, R.E.; Malhi, S.S. Greenhouse gas emissions from crop production systems in the Northern Great Plains of North America. In Recent Trends in Soil Science and Agronomy Research in the Northern Great Plains of North America; Malhi, S.S., Gan, Y., Schoenau, J.J., Lemke, R.L., Mark, A., Eds.; Liebig. Research Signpost: Kerala, India, 2010; pp. 95–108. [Google Scholar]
- Campbell, C.A.; Janzen, H.H.; Juma, N.G. Case studies of soil quality in the Canadian prairies: Long-term field experiments. Dev. Soil Sci. 1997, 25, 351–397. [Google Scholar]
- Assefa, B.A.; Schoenau, J.J.; Grevers, M.C.J. Effects of four annual applications of manure on Black Chernozemic soils. Can. Biosyst. Eng. 2004, 46, 39–46. [Google Scholar]
- Kar, G.; Schoenau, J.J.; Hilger, D.; Peak, D. Direct chemical speciation of soil Phosphorus in a Saskatchewan Chernozem after long- and short-term manure amendments. Can. J. Soil Sci. 2017, 97, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Dormaar, J.F.; Chang, C. Effects of 20 annual applications of excess feedlot manure on labile soil phosphorus. Can. J. Soil Sci. 1995, 75, 507–512. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Racz, G.J. Laboratory characterization of P in fresh and oven-dried organic amendments. J. Environ. Qual. 2004, 33, 1062–1069. [Google Scholar] [CrossRef]
- Schoenau, J.J.; Davis, J.G. Optimizing soil and plant responses to land-applied manure nutrients in the Great Plains of North America. Can. J. Soil Sci. 2006, 86, 587–595. [Google Scholar] [CrossRef]
- Hesterberg, D.; Weiqing, Z.; Hutchison, K.J.; Beauchemin, S.; Sayers, D.E. XAFS study of adsorbed and mineral forms of phosphate. J. Synchrotron Rad. 1999, 6, 636–638. [Google Scholar] [CrossRef]
- Beauchemin, S.; Hesterberg, D.; Chou, J.; Beauchemin, M.; Simard, R.R.; Sayers, D.E. Speciation of phosphorus in phosphorus-enriched agricultural soils using X-ray absorption near-edge structure spectroscopy and chemical fractionation. J. Environ. Qual. 2003, 32, 1809–1819. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Hu, Y.; Flaten, D.N. Phosphorus speciation of sequential extracts of organic amendments using nuclear magnetic resonance and X-ray absorption near-edge structure spectroscopies. J. Environ. Qual. 2007, 36, 1563–1576. [Google Scholar] [CrossRef]
- Ajiboye, B.; Akinremi, O.O.; Hu, Y.; Jurgensen, A. XANES Speciation of Phosphorus in Organically Amended and Fertilized Vertisol and Mollisol. Soil Sci. Soc. Am. J. 2008, 72, 1256–1262. [Google Scholar] [CrossRef]
- Kar, G.; Peak, D.; Schoenau, J.J. Spatial distribution and chemical speciation of soil phosphorus in a band application. Soil Sci. Soc. Am. J. 2012, 76, 2297–2306. [Google Scholar] [CrossRef]
- Prietzel, J.; Dümig, A.; Wu, Y.; Zhou, J.; Klysubun, W. Synchrotron-based P K-edge XANES spectroscopy reveals rapid changes of phosphorus speciation in the topsoil of two glacier foreland chronosequences. Geochim. Cosmochim. Acta 2013, 108, 154–171. [Google Scholar] [CrossRef]
- Alotaibi, K.D.; Schoenau, J.J.; Kar, G.; Peak, D.; Fonstad, T. Phosphorus speciation in a prairie soil amended with MBM and DDG ash: Sequential chemical extraction and synchrotron-based XANES spectroscopy investigations. Sci. Rep. 2018, 8, 3617. [Google Scholar] [CrossRef]
- Hamilton, J.G.; Grosskleg, J.; Hilger, D.; Bradshaw, K.; Carlson, T.; Siciliano, S.D.; Peak, D. Chemical speciation and fate of tripolyphosphate after application to a calcerous soil. Geochem. Trans. 2018, 19, 1. [Google Scholar] [CrossRef] [Green Version]
- Ingall, E.D.; Brandes, J.A.; Diaz, J.M.; de Jonge, M.D.; Paterson, D.; McNulty, I.; Elliott, W.C.; Northrup, P. Phosphorus Kedge XANES spectroscopy of mineral standards. J. Synchrotron Radiat. 2011, 18, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Negassa, W.; Kruse, J.; Michalik, D.; Appathurai, N.; Zuin, L.; Leinweber, P. Phosphorus Speciation in Agro-Industrial Byproducts: Sequential Fractionation, Solution 31P NMR, and P K-and L2,3-Edge XANES Spectroscopy. Environ. Sci. Technol. 2010, 44, 2092–2097. [Google Scholar] [CrossRef]
- Condron, L.M.; Newman, S. Revisiting the fundamentals of phosphorus fractionation of sediments and soils. J. Soils Sediments 2011, 11, 830–840. [Google Scholar] [CrossRef]
- Khare, N.; Hesterberg, D.; Martin, J.D. XANES investigation of phosphate sorption in single and binary systems of iron and aluminum oxide minerals. Environ. Sci. Technol. 2005, 39, 2152–2160. [Google Scholar] [CrossRef]
- Kelly, S.D.; Hesterberg, D.; Ravel, B. Analysis of soils and minerals using X-ray absorption spectroscopy. Methods of Soil Analysis. Part 5. Mineral. Methods 2008, 5, 387–463. [Google Scholar]
- Peak, D. Solid-state speciation of natural and alum-amended poultry litter using XANES spectroscopy. Environ. Sci. Technol. 2002, 36, 4253–4261. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Solomon, D.; Hyland, C.; Ketterings, Q.M.; Lehmann, J. Phosphorus speciation in manure and manure-amended soils using XANES spectroscopy. Environ. Sci. Technol. 2005, 39, 7485–7491. [Google Scholar] [CrossRef] [PubMed]
- Kruse, J.; Leinweber, P. Phosphorus in sequentially extracted fen peat soils: A K-edge X-ray absorption near-edge structure (XANES) spectroscopy study. Environ. Sci. Technol. 2008, 39, 7485–7491. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil Sampling and Methods of Analysis, 2nd ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2008. [Google Scholar]
- Zuo, M.; Renman, G.; Gustafsson, J.P.; Renman, A. Phosphorus removal performance and speciation in virgin and modified argon oxygen decarburisation slag designed for wastewater treatment. Water Res. 2015, 87, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, K.O.; Tighe, M.K.; Guppy, C.N.; Milham, P.J.; McLaren, T.I.; Schefe, C.R.; Lombi, E. XANES demonstrates the release of calcium phosphates from alkaline vertisols to moderately acidified solution. Environ. Sci. Technol. 2016, 50, 4229–4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, A.K.; Hillier, S.; Hesterberg, D.; Klysubun, W.; Ulén, B.; Gustafsson, J.P. Evolution of phosphorus speciation with depth in an agricultural soil profile. Geoderma 2016, 280, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Klopfenstein, T.J.; Angel, R.; Cromwell, G.; Erickson, G.E.; Fox, D.G.; Parsons, C.; Satter, L.D.; Sutton, A.L.; Baker, D.H.; Lewis, A.; et al. Animal diet modification to decrease the potential for nitrogen and phosphorus pollution. In Faculty Papers and Publications in Animal Science; University of Nebraska Press: Lincoln, NE, USA, 2002; p. 518. Available online: http://digitalcommons.unl.edu/animalscifacpub/518 (accessed on 8 January 2020).
- Toor, G.S.; Peak, D.J.; Sims, J.T. Phosphorus speciation in broiler litter and turkey manure produced from modified diets. J. Environ. Qual. 2005, 34, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Dendougui, F.; Schwedt, G. In vitro analysis of binding capacities of calcium to organic acid in different food samples. Eur. Food Res. Technol. 2004, 219, 409–415. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | Total P | Available P | eCEC | Org C | Sand | Clay | Silt | Texture | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | |
| (mg kg−1) | (cmolc kg−1) | % | ||||||||||||||||
| Amaranthus cruentus | ||||||||||||||||||
| T0 | 6.9 | 6.4 | 231.25 | 206.5 | 7.56 | 5.24 | 42.8 | 40.9 | 0.81 | 0.89 | 75 | 78 | 13 | 11 | 13 | 11 | SL | SL |
| T20 | 6.9 | 6.3 | 231 | 203.75 | 4.69 | 4.02 | 45.1 | 38.5 | 0.82 | 0.8 | 74 | 77 | 14 | 11 | 13 | 11 | SL | SL |
| T40 | 6.7 | 6.3 | 218.25 | 200.25 | 4.63 | 4.29 | 39.3 | 38.9 | 0.73 | 0.82 | 75 | 76 | 11 | 11 | 14 | 13 | SL | SL |
| T60 | 6.9 | 6.2 | 206.75 | 199.25 | 5.94 | 4.09 | 50.7 | 40 | 0.86 | 0.81 | 76 | 76 | 13 | 13 | 11 | 11 | SL | SL |
| T80 | 6.8 | 6.3 | 189.5 | 179.5 | 3.28 | 2.61 | 36.9 | 40.2 | 0.82 | 0.8 | 76 | 80 | 11 | 13 | 13 | 8 | SL | SL |
| Solanum macrocarpon | ||||||||||||||||||
| T0 | 6.2 | 6.7 | 347.3 | 273.8 | 4.05 | 5.79 | 49 | 64 | 1.09 | 1.14 | 75 | 75 | 13 | 14 | 13 | 11 | SL | SL |
| T20 | 6.2 | 6.7 | 321.5 | 280.3 | 4.1 | 6.15 | 48 | 61 | 1.04 | 1.14 | 76 | 79 | 11 | 5 | 13 | 16 | SL | LS |
| T40 | 6.1 | 6.7 | 291 | 264 | 3.28 | 5.12 | 47 | 57 | 1.02 | 1.1 | 75 | 81 | 11 | 7.5 | 14 | 11 | SL | LS |
| T60 | 6.2 | 6.8 | 278.5 | 232.8 | 5.91 | 4.53 | 52 | 60 | 1.03 | 1.06 | 74 | 77 | 13 | 6.3 | 14 | 16 | SL | LS |
| T80 | 6.1 | 7 | 239 | 236.5 | 2.95 | 3.06 | 40 | 51 | 0.98 | 0.94 | 72 | 80 | 14 | 5 | 14 | 15 | SL | LS |
| Vegetables | 0.199 | <0.001 | 0.846 | 0.004 | ||||||||||||||
| Treatments | 0.247 | 0.005 | 0.041 | 0.244 | ||||||||||||||
| Time | 0.555 | 0.009 | 0.665 | 0.082 | ||||||||||||||
| Vegetables * Treatments | 0.431 | 0.013 | 0.473 | 0.255 | ||||||||||||||
| Vegetables * Time | <0.001 | 0.157 | 0.111 | 0.411 | ||||||||||||||
| Treatments * Time | 0.093 | 0.367 | 0.281 | 0.015 | ||||||||||||||
| Vegetables * Treatments * Time | 0.246 | 0.166 | 0.159 | 0.199 | ||||||||||||||
| Treatment | pH | Total P | Available P | eCEC | Org C | Sand | Clay | Silt | Texture | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | BP | AH | |
| (mg kg−1) | (cmolc kg−1) | % | ||||||||||||||||
| Amaranthus cruentus | ||||||||||||||||||
| T0 | 6.7 | 6.3 | 1193.3 | 705.25 | 76.26 | 22.55 | 35.8 | 156 | 3.4 | 2.49 | 69.4 | 70.8 | 17 | 15 | 14 | 14 | SL | SL |
| T20 | 6.6 | 6.1 | 1442.8 | 950.5 | 105.1 | 22.04 | 247 | 39 | 3.8 | 4.21 | 69.3 | 65.5 | 19 | 20 | 12 | 14 | SL | SCL |
| T40 | 6.6 | 6 | 1775.5 | 841.25 | 195.14 | 41.73 | 303 | 217 | 3.4 | 3.57 | 70.8 | 68.9 | 15 | 16 | 14 | 16 | SL | SL |
| T60 | 6.3 | 5.5 | 710 | 530.25 | 64.41 | 27.81 | 150 | 128 | 2.2 | 2.48 | 74.8 | 73 | 8.8 | 13 | 16 | 14 | SL | SL |
| T80 | 5.8 | 5.8 | 603.25 | 409 | 54.85 | 16.39 | 186 | 104 | 2.1 | 2.25 | 73.6 | 74.7 | 8.8 | 8.8 | 18 | 17 | SL | SL |
| Solanum macrocarpon | ||||||||||||||||||
| T0 | 5.6 | 5.3 | 892 | 1164.8 | 54.48 | 64.97 | 172 | 218 | 3 | 3.49 | 63.2 | 56.5 | 19 | 26 | 18 | 18 | SL | SCL |
| T20 | 5.1 | 4.6 | 944.5 | 965.5 | 52.96 | 35.34 | 176 | 157 | 3.6 | 2.92 | 53.5 | 51.4 | 30 | 33 | 16 | 15 | SCL | SCL |
| T40 | 4.8 | 4.8 | 796.75 | 766 | 81.56 | 84.69 | 169 | 207 | 2.8 | 2.53 | 81.1 | 71.9 | 7.6 | 15 | 11 | 13 | LS | SL |
| T60 | - | 5.1 | - | 1152.5 | - | 73.19 | - | 254 | - | 3.78 | - | 67.7 | - | 23 | - | 9 | - | SCL |
| T80 | 3.5 | 4.3 | 421.75 | 431.75 | 18.18 | 26.68 | 91.7 | 61.5 | 1.8 | 1.86 | 79.9 | 85 | 10 | 5 | 10 | 10 | SL | LS |
| Vegetables | 0.001 | 0.030 | 0.110 | 0.304 | ||||||||||||||
| Treatments | 0.043 | <0.001 | 0.095 | <0.001 | ||||||||||||||
| Time | 0.120 | 0.171 | 0.021 | 0.001 | ||||||||||||||
| Vegetables * Treatments | 0.010 | 0.012 | 0.020 | 0.011 | ||||||||||||||
| Vegetables * Time | 0.033 | 0.005 | 0.016 | 0.053 | ||||||||||||||
| Treatments * Time | 0.034 | 0.454 | 0.510 | 0.010 | ||||||||||||||
| Vegetables * Treatments * Time | 0.065 | 0.150 | 0.319 | 0.079 | ||||||||||||||
| Treatment | Goodness of Fit | % | |||||
|---|---|---|---|---|---|---|---|
| R Factor | χ2 | Organic P | Adsorbed P | Apatite P | |||
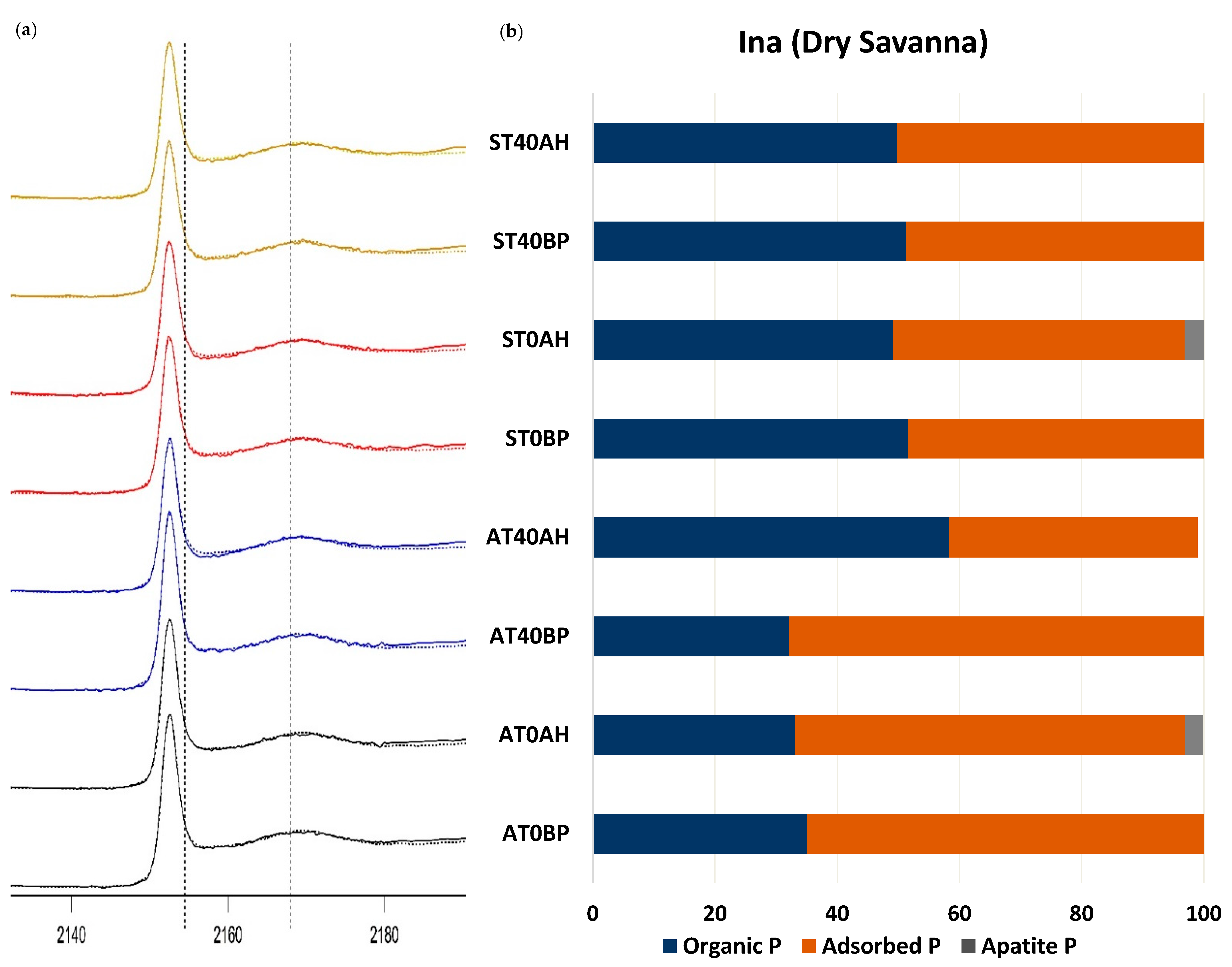
| Ina (dry savanna) | AT0 | BP | 0.00162 | 0.2089 | 35.1 (±1.4) | 64.9 (±0.5) | |
| AH | 0.00202 | 0.2530 | 33.1 (±1.5) | 63.9 (±0.5) | 2.9 (±2.2) | ||
| AT40 | BP | 0.00179 | 0.2382 | 32.1 (±1.5) | 67.9 (±0.5) | ||
| AH | 0.0038 | 0.3863 | 58.3 (±1.9) | 40.7 (±0.6) | |||
| ST0 | BP | 0.00231 | 0.2472 | 51.6 (±1.5) | 48.4 (±0.5) | ||
| AH | 0.00211 | 0.2224 | 49.1 (±1.4) | 47.8 (±0.5) | 3.1 (±2.1) | ||
| ST40 | BP | 0.00233 | 0.2493 | 51.3 (±1.5) | 48.7 (±0.5) | ||
| AH | 0.00233 | 0.2529 | 49.8 (±1.5) | 50.2 (±0.5) | |||
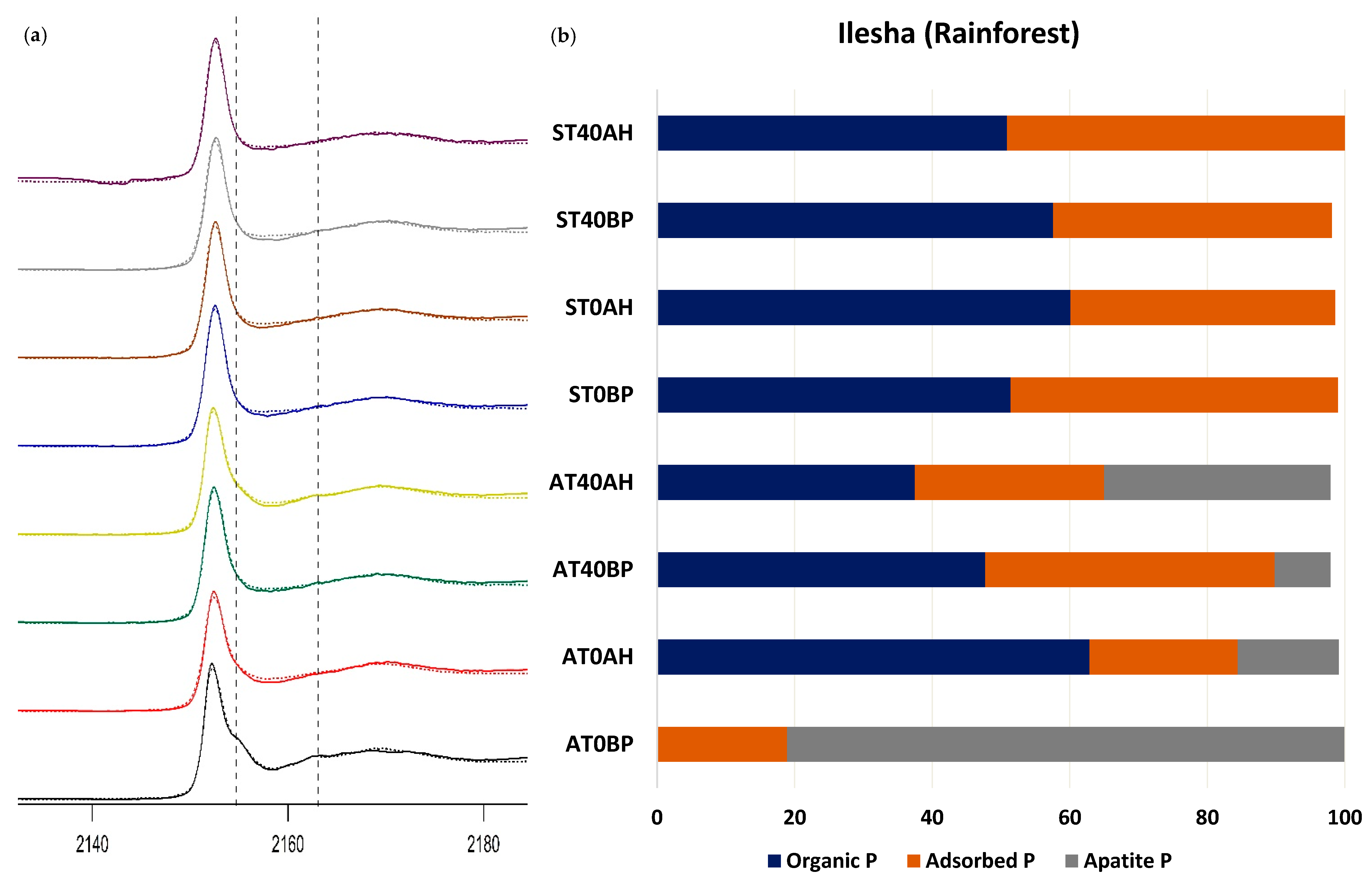
| Ilesha (rainforest) | AT0 | BP | 0.00274 | 0.2861 | 18.9 (±0.6) | 81.0 (±2.2) | |
| AH | 0.00917 | 0.7632 | 62.9 (±2.6) | 21.5 (±0.9) | 14.7 (±3.1) | ||
| AT40 | BP | 0.00273 | 0.2796 | 47.7 (±1.6) | 42.1 (±0.5) | 8.1 (±2.2) | |
| AH | 0.00492 | 0.4532 | 37.5 (±2.0) | 27.5 (±0.7) | 32.9 (±2.6) | ||
| ST0 | BP | 0.00325 | 0.3457 | 51.4 (±1.8) | 47.6 (±0.6) | ||
| AH | 0.00539 | 0.5236 | 60.1 (±2.2) | 38.5 (±0.7) | |||
| ST40 | BP | 0.00373 | 0.3733 | 57.6 (±1.8) | 40.5 (±0.6) | ||
| AH | 0.00316 | 0.3449 | 50.9 (±1.8) | 49.1 (±0.6) | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olaleye, A.; Oyedele, D.; Akponikpe, P.; Kar, G.; Peak, D. Molecular Scale Studies of Phosphorus Speciation and Transformation in Manure Amended and Microdose Fertilized Indigenous Vegetable Production Systems of Nigeria and Republic of Benin. Soil Syst. 2020, 4, 5. https://doi.org/10.3390/soilsystems4010005
Olaleye A, Oyedele D, Akponikpe P, Kar G, Peak D. Molecular Scale Studies of Phosphorus Speciation and Transformation in Manure Amended and Microdose Fertilized Indigenous Vegetable Production Systems of Nigeria and Republic of Benin. Soil Systems. 2020; 4(1):5. https://doi.org/10.3390/soilsystems4010005
Chicago/Turabian StyleOlaleye, Abimfoluwa, Durodoluwa Oyedele, Pierre Akponikpe, Gourango Kar, and Derek Peak. 2020. "Molecular Scale Studies of Phosphorus Speciation and Transformation in Manure Amended and Microdose Fertilized Indigenous Vegetable Production Systems of Nigeria and Republic of Benin" Soil Systems 4, no. 1: 5. https://doi.org/10.3390/soilsystems4010005
APA StyleOlaleye, A., Oyedele, D., Akponikpe, P., Kar, G., & Peak, D. (2020). Molecular Scale Studies of Phosphorus Speciation and Transformation in Manure Amended and Microdose Fertilized Indigenous Vegetable Production Systems of Nigeria and Republic of Benin. Soil Systems, 4(1), 5. https://doi.org/10.3390/soilsystems4010005