

Is the Current Modelling of Litter Decomposition Rates Reliable under Limiting Environmental Conditions Induced by Ongoing Climate Change?



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Climatic Data
2.2. Field Data Collection and Laboratory Analysis
2.3. Decomposition Model
2.3.1. Climatic Module
2.3.2. Hydrological Module
2.3.3. Decomposition Module
3. Results
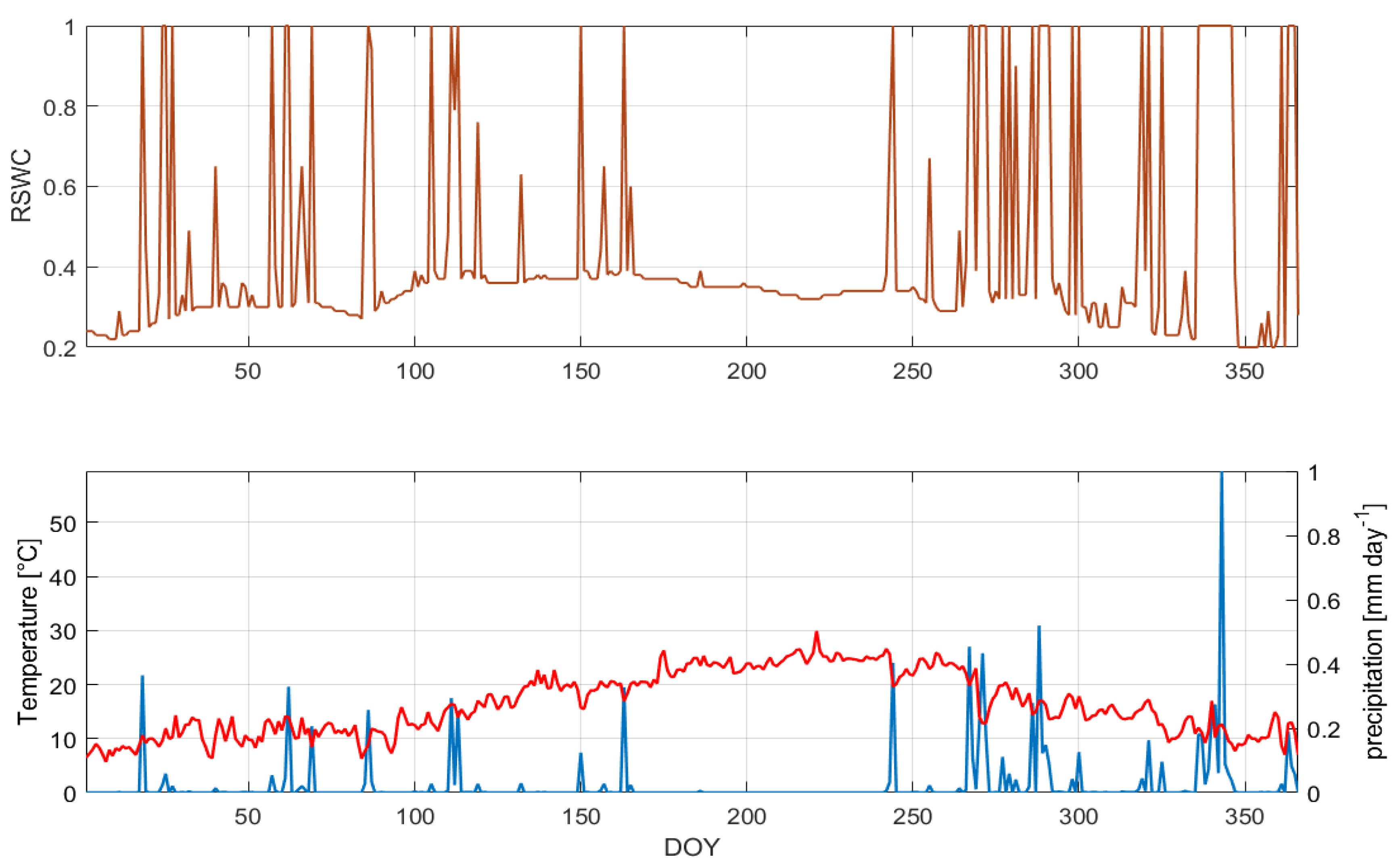
3.1. Climatic Condition and Relative Soil Water Content
3.2. Decomposition Dynamics
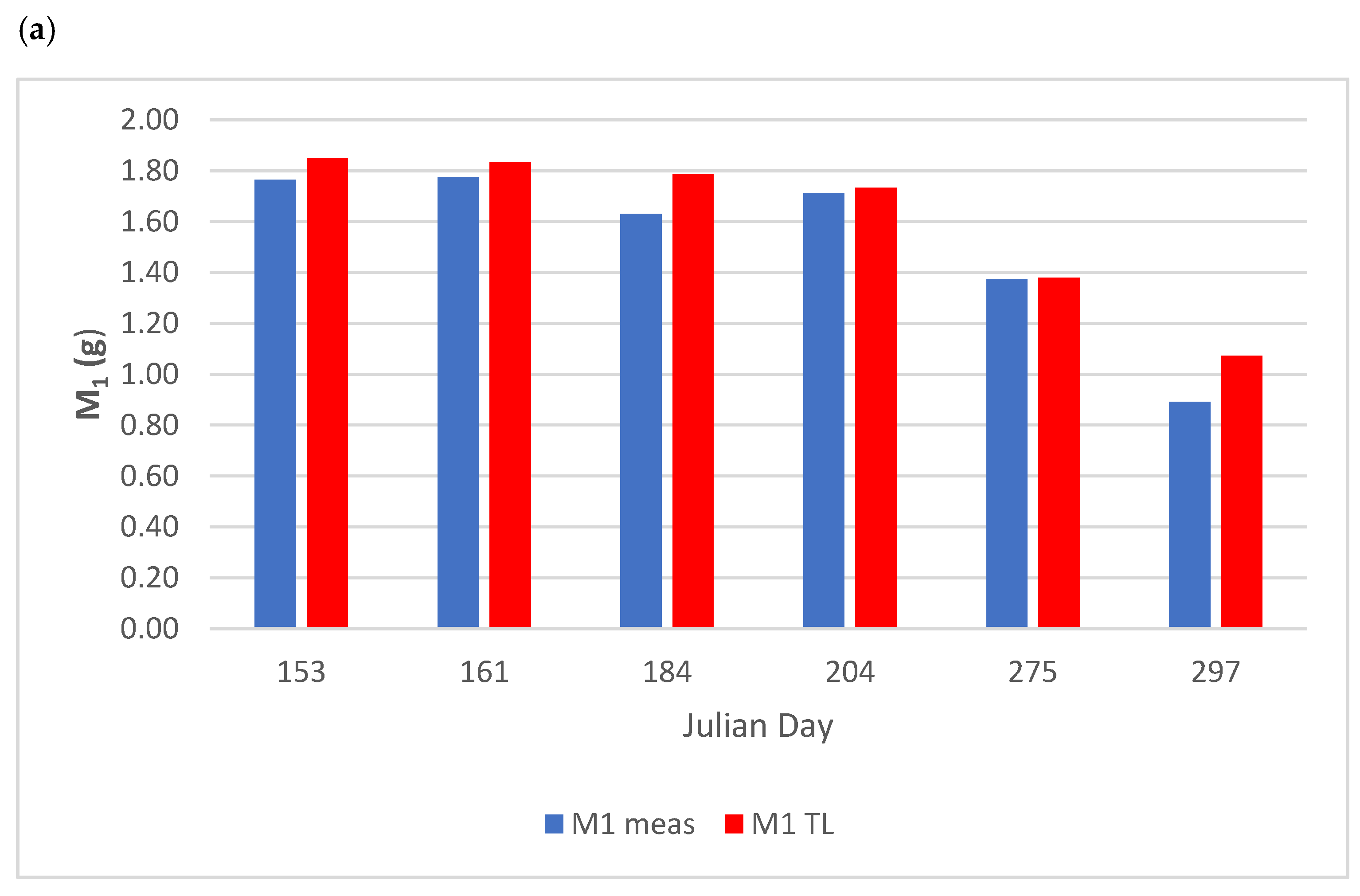
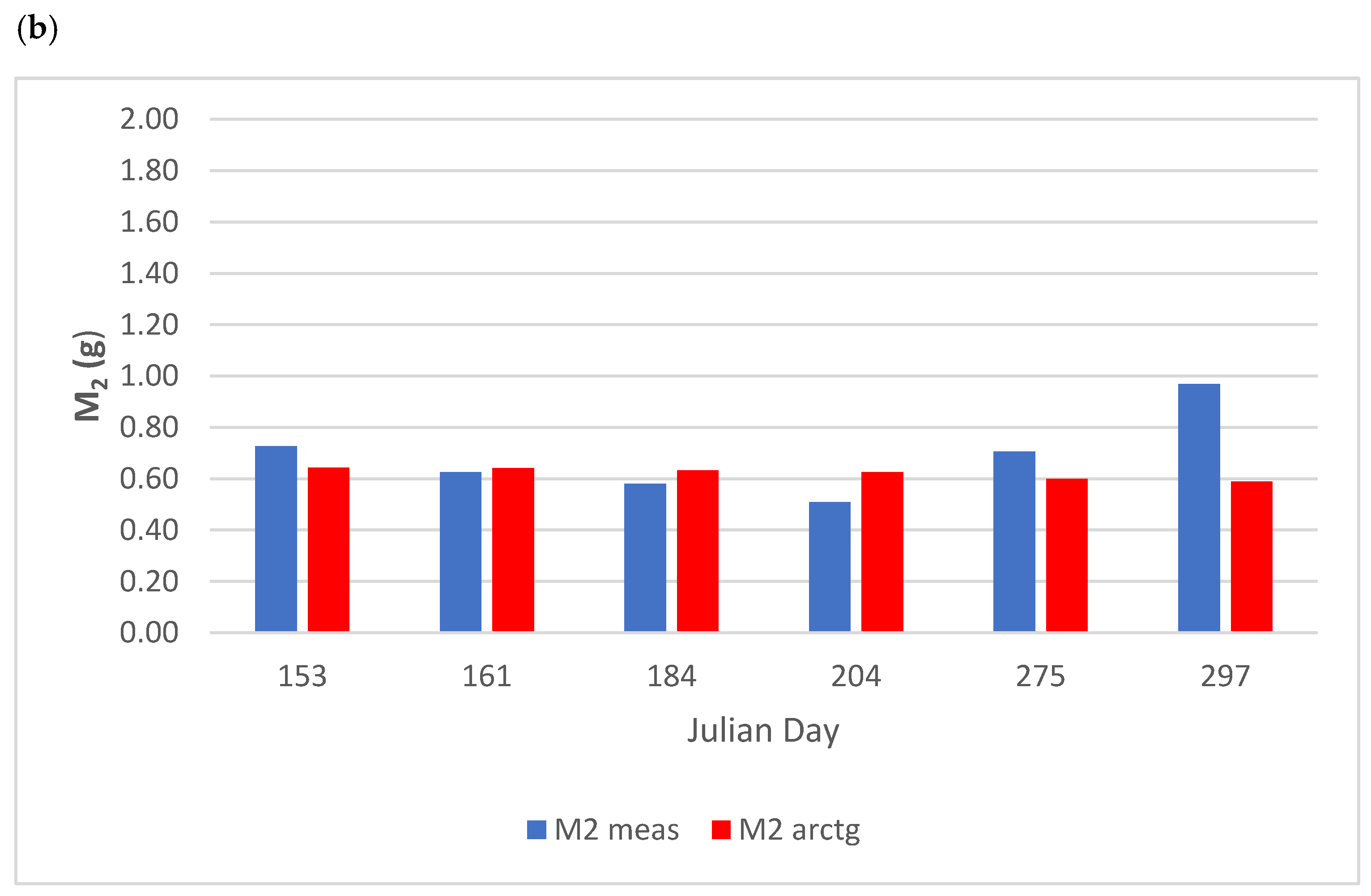
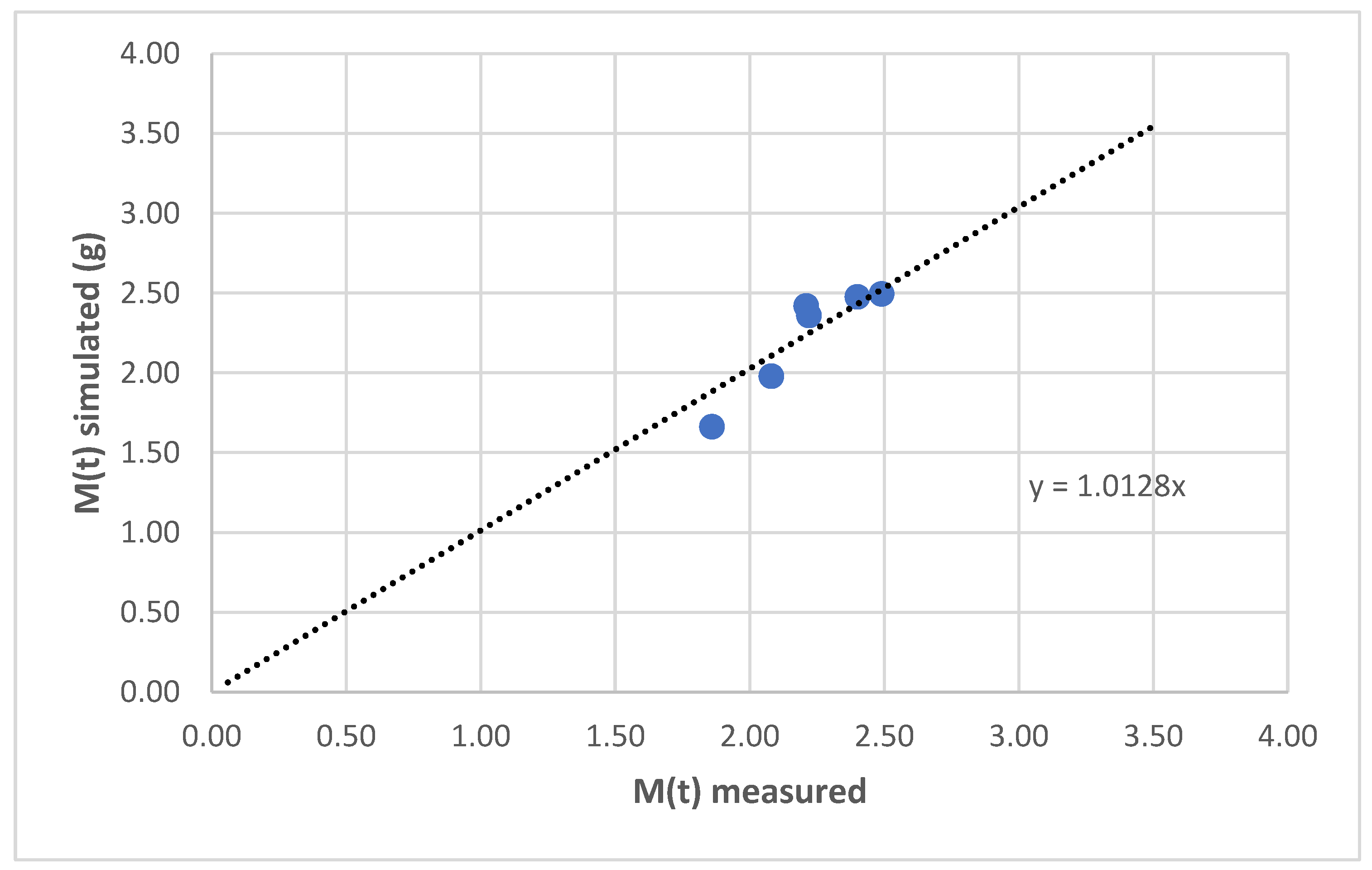
3.2.1. Biomass Variation
3.2.2. ADF, Cellulose, and Lignin Variations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bradford, M.A.; Wieder, W.R.; Bonan, G.B.; Fierer, N.; Raymond, P.A.; Crowther, T.W. Managing Uncertainty in Soil Carbon Feedbacks to Climate Change. Nat. Clim. Chang. 2016, 6, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Luo, Y.; Chen, H.Y.H.; Ruan, H. Responses of Litter Decomposition and Nutrient Release to N Addition: A Meta-Analysis of Terrestrial Ecosystems. Appl. Soil Ecol. 2018, 128, 35–42. [Google Scholar] [CrossRef]
- Zhang, M.; Cheng, X.; Geng, Q.; Shi, Z.; Luo, Y.; Xu, X. Leaf Litter Traits Predominantly Control Litter Decomposition in Streams Worldwide. Glob. Ecol. Biogeogr. 2019, 28, 1469–1486. [Google Scholar] [CrossRef]
- Bonan, G.B.; Hartman, M.D.; Parton, W.J.; Wieder, W.R. Evaluating Litter Decomposition in Earth System Models with Long-Term Litterbag Experiments: An Example Using the Community Land Model Version 4 (CLM4). Glob. Chang. Biol. 2013, 19, 957–974. [Google Scholar] [CrossRef]
- Chapin, F.S.; Folke, C.; Kofinas, G.P. A Framework for Understanding Change. In Principles of Ecosystem Stewardship; Springer: New York, NY, USA, 2009; pp. 3–28. [Google Scholar]
- Cai, A.; Liang, G.; Yang, W.; Zhu, J.; Han, T.; Zhang, W.; Xu, M. Patterns and Driving Factors of Litter Decomposition across Chinese Terrestrial Ecosystems. J. Clean. Prod. 2021, 278, 123964. [Google Scholar] [CrossRef]
- Frøseth, R.B.; Bleken, M.A. Effect of Low Temperature and Soil Type on the Decomposition Rate of Soil Organic Carbon and Clover Leaves, and Related Priming Effect. Soil Biol. Biochem. 2015, 80, 156–166. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter Decomposition: Effects of Temperature Driven by Soil Moisture and Vegetation Type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Bryant, D.M.; Holland, E.A.; Seastedt, T.R.; Walker, M.D. Analysis of Litter Decomposition in an Alpine Tundra. Can. J. Bot. 1998, 76, 1295–1304. [Google Scholar] [CrossRef]
- Knorr, M.; Frey, S.D.; Curtis, P.S. Nitrogen Additions and Litter Decomposition: A Meta-Analysis. Ecology 2005, 86, 3252–3257. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.; Amitrano, W.; Hoshika, Y.; Paoletti, E. Plant Species-Specific Litter Decomposition Rates Are Directly Affected by Tropospheric Ozone: Analysis of Trends and Modelling. Water Air Soil Pollut. 2019, 230, 311. [Google Scholar] [CrossRef]
- Dang, C.K.; Chauvet, E.; Gessner, M.O. Magnitude and Variability of Process Rates in Fungal Diversity-Litter Decomposition Relationships. Ecol. Lett. 2005, 8, 1129–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graça, M.A.; Bärlocher, F.; Gessner, M.O. Methods to Study Litter Decomposition: A Practical Guide; Springer: Cham, Switzerland, 2005. [Google Scholar]
- Hooper, D.U.; Vitousek, P.M. The Effects of Plant Composition and Diversity on Ecosystem Processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef] [Green Version]
- Mclaren, J.R.; Turkington, R. Plant Functional Group Identity Differentially Affects Leaf and Root Decomposition. Glob. Chang. Biol. 2010, 16, 3075–3084. [Google Scholar] [CrossRef]
- Dirks, I.; Navon, Y.; Kanas, D.; Dumbur, R.; Grünzweig, J.M. Atmospheric Water Vapor as Driver of Litter Decomposition in Mediterranean Shrubland and Grassland during Rainless Seasons. Glob. Chang. Biol. 2010, 16, 2799–2812. [Google Scholar] [CrossRef]
- Evans, S.; Todd-Brown, K.E.O.; Jacobson, K.; Jacobson, P. Non-Rainfall Moisture: A Key Driver of Microbial Respiration from Standing Litter in Arid, Semiarid, and Mesic Grasslands. Ecosystems 2020, 23, 1154–1169. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Tenhunen, J.D.; Roupsard, O.; Ourcival, J.-M.; Rambal, S.; Dore, S.; Valentini, R. Ecosystem Respiration in Two Mediterranean Evergreen Holm Oak Forests: Drought Effects and Decomposition Dynamics. Funct. Ecol. 2002, 16, 27–39. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Lin, Y.; Han, G.; Guo, P.; Tian, X. The effect of temperature on decomposition of leaf litter from two tropical forests by a microcosm experiment. Eur. J. Soil Biol. 2010, 46, 200–207. [Google Scholar] [CrossRef]
- Van Meeteren, M.J.M.; Tietema, A.; Westerveld, J.W. Regulation of microbial carbon, nitrogen, and phosphorus transformations by temperature and moisture during decomposition of Calluna vulgaris litter. Biol. Fertil. Soils 2007, 44, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Dijkstra, P.; Kock, G.W.; Peñuelas, J.; Hungate, B.A. Responses of Terrestrial Ecosystems to Temperature and Precipitation Change: A Meta-Analysis of Experimental Manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef] [Green Version]
- Saccone, P.; Morin, S.; Baptist, F.; Bonneville, J.-M.; Colace, M.-P.; Domine, F.; Faure, M.; Geremia, R.; Lochet, J.; Poly, F.; et al. The Effects of Snowpack Properties and Plant Strategies on Litter Decomposition during Winter in Subalpine Meadows. Plant Soil 2013, 363, 215–229. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant Species Traits Are the Predominant Control on Litter Decomposition Rates within Biomes Worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Incerti, G.; Bonanomi, G.; Giannino, F.; Rutigliano, F.A.; Piermatteo, D.; Castaldi, S.; De Marco, A.; Fierro, A.; Fioretto, A.; Maggi, O.; et al. Litter Decomposition in Mediterranean Ecosystems: Modelling the Controlling Role of Climatic Conditions and Litter Quality. Appl. Soil Ecol. 2011, 49, 148–157. [Google Scholar] [CrossRef]
- Jarvis, S.C.; Stockdale, E.A.; Shepherd, M.A.; Powlson, D.S. Nitrogen Mineralization in Temperate Agricultural Soils: Processes and Measurement. Adv. Agron. 1996, 57, 187–235. [Google Scholar] [CrossRef]
- Xu, J.M.; Tang, C.; Chen, Z.L. Chemical Composition Controls Residue Decomposition in Soils Differing in Initial pH. Soil Biol. Biochem. 2006, 38, 544–552. [Google Scholar] [CrossRef]
- Del Grosso, S.J.; Parton, W.J.; Mosier, A.R.; Holland, E.A.; Pendall, E.; Schimel, D.S.; Ojima, D.S. Modeling Soil CO2 Emissions from Ecosystems. Biogeochemistry 2005, 73, 71–91. [Google Scholar] [CrossRef]
- Adair, E.C.; Parton, W.J.; Del Grosso, S.J.; Silver, W.L.; Harmon, M.E.; Hall, S.A.; Burke, I.C.; Hart, S.C. Simple Three-Pool Model Accurately Describes Patterns of Long-Term Litter Decomposition in Diverse Climates. Glob. Chang. Biol. 2008, 14, 2636–2660. [Google Scholar] [CrossRef]
- Sierra, C.A.; Müller, M.; Trumbore, S.E. Models of Soil Organic Matter Decomposition: The SoilR Package, Version 1.0. Geosci. Model Dev. 2012, 5, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Cutini, A. La Stima Del LAI Con Il Metodo Delle Misure Di Trasmittanza in Popolamenti Diradati e Non Diradati Di Cerro. Ann. Ist. Sper. Selv. Arezzo 1994, 23, 167–181. [Google Scholar]
- Gahrooe, F.R. Impacts of Elevated Atmospheric CO2 on Litter Quality, Litter Decomposability and Nitrogen Turnover Rate of Two Oak Species in a Mediterranean Forest Ecosystem. Glob. Chang. Biol. 1998, 4, 667–677. [Google Scholar] [CrossRef]
- Wider, R.K.; Lang, G.E. A Critique of the Analytical Methods Used in Examining Decomposition Data Obtained From Litter Bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef]
- Fioretto, A.; Papa, S.; Fuggi, A. Litter-Fall and Litter Decomposition in a Low Mediterranean Shrubland. Biol. Fertil. Soils 2003, 39, 37–44. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and Litter Decomposition in Terrestrial Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Schneider, T.; Roschitzki, B.; Schmid, E.; Eberl, L.; Hämmerle, I.; Leitner, S.; Richter, A.; Wanek, W.; Riedel, K.; et al. Effects of Stoichiometry and Temperature Perturbations on Beech Leaf Litter Decomposition, Enzyme Activities and Protein Expression. Biogeosciences 2012, 9, 4537–4551. [Google Scholar] [CrossRef] [Green Version]
- Van Soest, P.J.; Wine, R.H. Determination of Lignin and Cellulose in Acid-Detergent Fiber with Permanganate. J. AOAC Int. 1968, 51, 780–785. [Google Scholar] [CrossRef]
- Fioretto, A.; Di Nardo, C.; Papa, S.; Fuggi, A. Lignin and Cellulose Degradation and Nitrogen Dynamics during Decomposition of Three Leaf Litter Species in a Mediterranean Ecosystem. Soil Biol. Biochem. 2005, 37, 1083–1091. [Google Scholar] [CrossRef]
- Vitale, M.; Savi, F.; Baldantoni, D.; Attorre, F. Modeling of Early Stage Litter Decomposition in Mediterranean Mixed Forests: Functional Aspects Affected by Local Climate. iFor.—Biogeosci. For. 2014, 8, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Chiesi, M.; Angeli, L.; Battista, P.; Bottai, L.; Fibbi, L.; Gardin, L.; Gozzini, B.; Rapi, B.; Romani, M.; Sabatini, F.; et al. Bilancio Idrico Multiscala Di Aree Forestali e Agricole. In Sistemi Integrati per il Monitoraggio Ambientale e il Supporto alla Gestione delle Risorse. Componenti, Procedure e Prospettive; Raschi, A., Conese, C., Battista, P., Eds.; CNR-IBIMET: Florence, Italy, 2016; pp. 131–138. [Google Scholar]
- Gardin, L.; Chiesi, M.; Fibbi, L.; Angeli, L.; Rapi, B.; Battista, P.; Maselli, F. Simulation of Soil Water Content through the Combination of Meteorological and Satellite Data. Geoderma 2021, 393, 115003. [Google Scholar] [CrossRef]
- Maselli, F.; Papale, D.; Chiesi, M.; Matteucci, G.; Angeli, L.; Raschi, A.; Seufert, G. Operational Monitoring of Daily Evapotranspiration by the Combination of MODIS NDVI and Ground Meteorological Data: Application and Evaluation in Central Italy. Remote Sens. Environ. 2014, 152, 279–290. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the Temperature Dependence of Soil Respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Walter, J. Effects of Changes in Soil Moisture and Precipitation Patterns on Plant-Mediated Biotic Interactions in Terrestrial Ecosystems. Plant Ecol. 2018, 219, 1449–1462. [Google Scholar] [CrossRef]
- León-Sánchez, L.; Nicolás, E.; Nortes, P.A.; Maestre, F.T.; Querejeta, J.I. Photosynthesis and Growth Reduction with Warming Are Driven by Nonstomatal Limitations in a Mediterranean Semi-Arid Shrub. Ecol. Evol. 2016, 6, 2725–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León-Sánchez, L.; Nicolás, E.; Goberna, M.; Prieto, I.; Maestre, F.T.; Querejeta, J.I. Poor Plant Performance under Simulated Climate Change Is Linked to Mycorrhizal Responses in a Semi-Arid Shrubland. J. Ecol. 2018, 106, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Gliksman, D.; Rey, A.; Seligmann, R.; Dumbur, R.; Sperling, O.; Navon, Y.; Haenel, S.; De Angelis, P.; Arnone, J.A.; Grünzweig, J.M. Biotic Degradation at Night, Abiotic Degradation at Day: Positive Feedbacks on Litter Decomposition in Drylands. Glob. Chang. Biol. 2017, 23, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Poulter, B.; Frank, D.; Ciais, P.; Myneni, R.B.; Andela, N.; Bi, J.; Broquet, G.; Canadell, J.G.; Chevallier, F.; Liu, Y.Y.; et al. Contribution of Semi-Arid Ecosystems to Interannual Variability of the Global Carbon Cycle. Nature 2014, 509, 600–603. [Google Scholar] [CrossRef] [Green Version]
- Caraux Garson, D.; Lacaze, B. Monitoring Leaf Area Index of Mediterranean Oak Woodlands: Comparison of Remotely-Sensed Estimates with Simulations from an Ecological Process-Based Model. Int. J. Remote Sens. 2003, 24, 3441–3456. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2021.










{kind=link}
{kind=link}
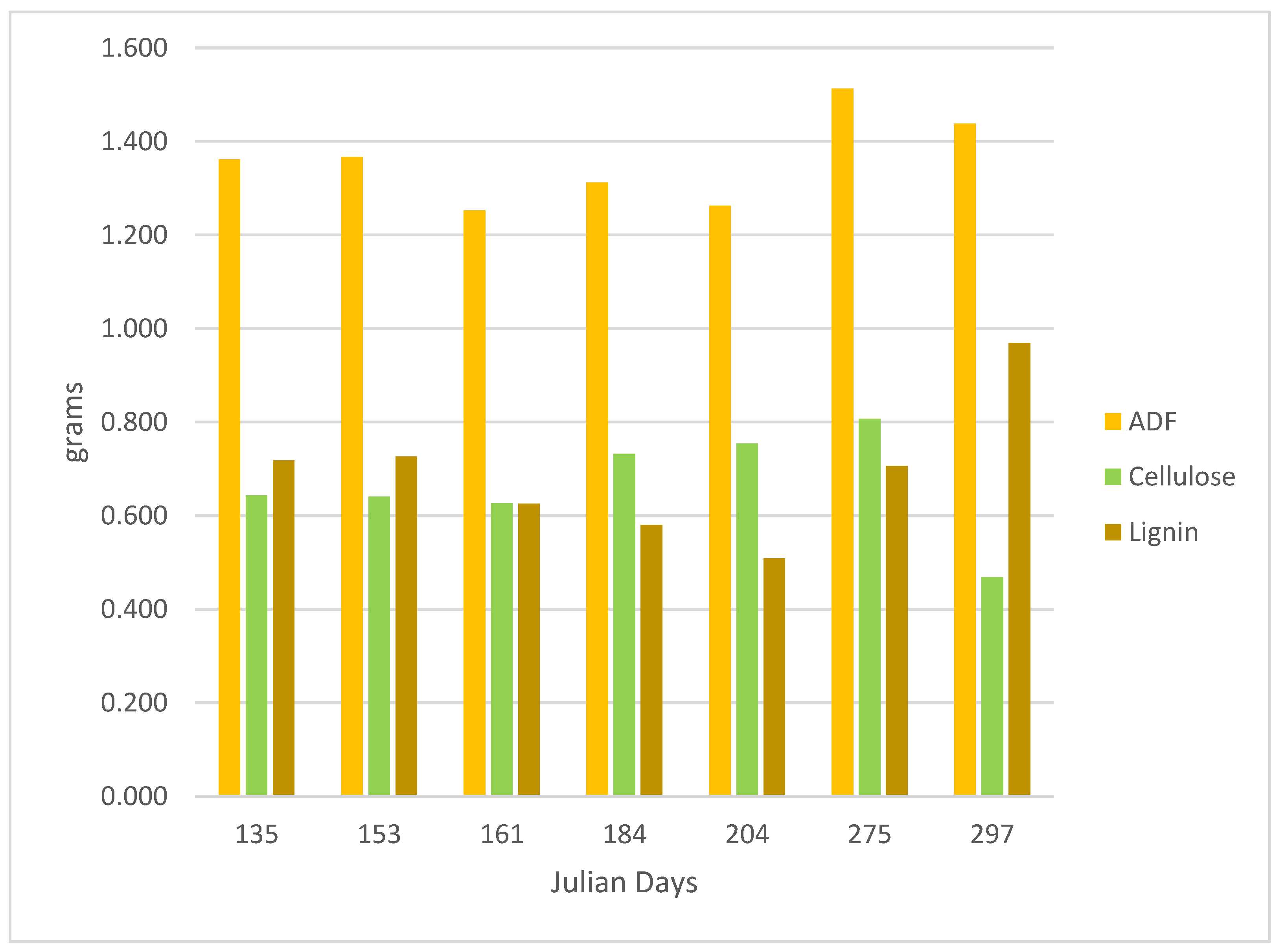{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Date | Sampling Days | Julian Days | Remaining Biomass (Avg; g) ± St. Dev. | Remaining Percentage (%) |
|---|---|---|---|---|
| 1 June 2020 | 15 | 153 | 2.49 ± 0.01 | 99.8 |
| 9 June 2020 | 23 | 161 | 2.40 ± 0.04 | 96.2 |
| 2 July 2020 | 46 | 184 | 2.21 ± 0.04 | 88.4 |
| 22 July 2020 | 66 | 204 | 2.22 ± 0.05 | 89.0 |
| 1 October 2020 | 137 | 275 | 2.08 ± 0.11 | 83.2 |
| 23 October 2020 | 159 | 297 | 1.86 ± 0.11 | 74.6 |
| M1-M2 | R2 | MSE |
|---|---|---|
| M1 Arctan | 0.961 | 0.364 |
| M1 Linear | 0.962 | 0.072 |
| M1 LT | 0.961 | 0.069 |
| M2 Arctan | 0.439 | 0.180 |
| M2 Linear | 0.442 | 0.217 |
| M2 LT | 0.440 | 0.218 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranucci, M.; Perez, M.; Lombardi, D.; Vitale, M. Is the Current Modelling of Litter Decomposition Rates Reliable under Limiting Environmental Conditions Induced by Ongoing Climate Change? Soil Syst. 2022, 6, 81. https://doi.org/10.3390/soilsystems6040081
Ranucci M, Perez M, Lombardi D, Vitale M. Is the Current Modelling of Litter Decomposition Rates Reliable under Limiting Environmental Conditions Induced by Ongoing Climate Change? Soil Systems. 2022; 6(4):81. https://doi.org/10.3390/soilsystems6040081
Chicago/Turabian StyleRanucci, Maddalena, Martina Perez, Danilo Lombardi, and Marcello Vitale. 2022. "Is the Current Modelling of Litter Decomposition Rates Reliable under Limiting Environmental Conditions Induced by Ongoing Climate Change?" Soil Systems 6, no. 4: 81. https://doi.org/10.3390/soilsystems6040081
APA StyleRanucci, M., Perez, M., Lombardi, D., & Vitale, M. (2022). Is the Current Modelling of Litter Decomposition Rates Reliable under Limiting Environmental Conditions Induced by Ongoing Climate Change? Soil Systems, 6(4), 81. https://doi.org/10.3390/soilsystems6040081